南方红壤侵蚀区不同植被恢复年限下芒萁叶功能性状对土壤因子的响应
2020-03-27王敬哲陈志强陈志彪潘宗涛
王敬哲,陈志强,3,*,陈志彪,3,潘宗涛
1 福建师范大学地理科学学院,福州 350007 2 湿润亚热带山地生态国家重点实验室培育基地,福州 350007 3 福建师范大学地理研究所,福州 350007
植物功能性状被定义为能够强烈影响生态系统功能以及能够反映植被对环境变化响应的核心植物属性,是植物沟通环境的桥梁[1- 5]。叶是植物体与环境相接触、且对环境变化极其敏感的器官[1]。在众多植物功能性状中,叶功能性状与植物C的同化及对资源的利用能力紧密联系,能够反映植物适应环境变化所形成的生存对策[2- 4]。因此,对植物叶功能性状的研究越来越受到研究者的重视[6]。
关于土壤侵蚀退化地植被叶功能性状的变化,黄土高原土壤侵蚀区已做过一定研究。例如,段媛媛等[7]研究了不同林龄下刺槐(RobiniapseudoacaciaL.)叶功能性状差异。结果表明,刺槐具有较强的叶片形态可塑性,其能改变自身的形态结构,形成最佳功能组合以适应环境的变化。赵祥等[8- 10]研究了不同退化草地达乌里胡枝子(Lespedezadavurica)叶片形态特征的变异,发现同一性状在不同生态型内变异幅度差异明显,并作出了生境因子会影响达乌里胡枝子叶片性状的推断。
芒萁(Dicranopterisdichotoma)作为我国亚热带地区广泛分布的古老蕨类植物,具有喜酸、喜阳、耐旱、耐贫瘠的特点[11],根系发达,繁殖力极强[12],是南方红壤侵蚀区生态退化过程中最后退出,以及生态恢复过程中最早进入的草本植物之一[13-14]。关于南方红壤侵蚀区芒萁的生态效应前人已做过大量研究。黄俊等[15]研究了芒萁覆盖对土壤团聚体稳定性的影响,发现芒萁的地下根系促进了土壤有机质的积累,从而显著提高土壤团聚体稳定性。聂阳意等[16]研究了芒萁覆盖下的红壤氮组分,发现有芒萁覆盖的红壤较裸地氮含量显著降低。目前对于在植被恢复过程中芒萁特性的变化还未有研究,尤其是不同恢复阶段芒萁叶功能性状的差异及影响因子我们还尚不清楚。了解植被恢复过程中芒萁叶功能性状的变化、认识芒萁叶功能性状的环境相应机制仍是亟需探讨的问题。
本文以不同植被恢复年限(0年、6年、12年、18年、36年、80年)下的芒萁群落为研究对象,选取南方红壤侵蚀区芒萁的11个有代表性的叶功能性状指标,研究叶功能性状在不同恢复阶段所呈现的环境适应性的变化规律,探讨引起不同叶功能性状变化的关键土壤因子,进而深入认识芒萁对南方红壤侵蚀区贫瘠土壤环境的适应策略及植被恢复过程中土壤质量的演变规律,为侵蚀退化地的植被恢复与管理提供科学依据。
1 研究区域与研究方法
1.1 研究区概况
研究区位于福建省长汀县河田镇朱溪小流域(116°00′45″—116°39′20″E,25°18′40″—26°02′05″N),属于亚热带季风湿润气候,年均气温约18.5℃,年均降雨量约1710mm,年均蒸发量约1400mm。该区域土壤类型为粗晶花岗岩风化发育形成的红壤、侵蚀性红壤[17]。由于土壤中砂粒含量较高,抗蚀能力差,加之长久以来人类活动的干扰,使得以常绿阔叶林为主的地带性植被遭到严重破坏,地表裸露,形成严重的水土流失。自上个世纪以来,当地水土保持部门采用多种措施恢复裸露的地表,其中较为常用的为乔灌草混交的生态林草措施:乔木以马尾松(PinusmassonianaLamb.)、杉木(Cunninghamialanceolata)为主,灌木主要有木荷(SchimasuperbaGardn. etChamp.)、枫香(LiquidambarformosanaHance),草本主要为芒萁、宽叶雀稗(PaspalumwettsteiniiHackel.)等。可是随着恢复年限的增长,大量的引进草本最终被芒萁所取代。芒萁作为当地乡土植被,在植被恢复地形成最具优势的先锋植被[14]。
1.2 样地选择
根据长汀县水土保持事业局提供的生态林草措施下不同植被恢复期样地的相关资料,于2017年9月对样地进行考察布设。依据典型性和代表性原则,选取植被恢复0年、6年、12年、18年、36年、80年生态恢复地中的芒萁群落为研究对象。因芒萁为乡土先锋植被,且大面积人工繁育困难,各样地芒萁均为自然侵入。利用手持式GPS(集思宝UG905,中国)测量样地详细信息(表1)。

表1 不同植被恢复年限样地概况
来油坑对照地 LYK(C) Laiyoukeng(Contrast); 来油坑治理地 LYK(R) Laiyoukeng(recovery); 龙颈治理地 LJ(R) Longjing(recovery); 游坊治理地YF(R) Youfang(recovery); 八十里河治理地 BSLH(R) Bashilihe(recovery); 露湖对照地 LH(C) Luhu(Contrast)
1.3 样品采集
野外样品采集于2017年9月26至28日。自上坡位至下坡位于各样地设置3个1m×1m芒萁样方。在每个样方内分别从东、西、南、北4个方位不同株芒萁上采集叶片,每个方位随机采集5片,采集时选择高度适中、成熟健康的叶片,并用润湿滤纸相夹,编号后装入自封袋,再放入4℃冰盒以备形态指标测定。
用土钻法采集土样。于样方中心打入10cm土钻,每个样方设置3组重复,相同样地不同样方所得土壤混合均匀,土样风干、研磨、过筛、备用。相同位置用铝盒法采集土样,用以测量土壤含水率。
1.4 测定指标与方法
1.4.1叶功能性状指标的测定
每个方向采集的叶片中,2片用于叶绿素含量(mg/g)的测定,使用UV=2450紫外-可见分光光度计,在波长663、645nm下测定吸光度,在Amon公式下计算叶绿素含量[18]。剩余叶片使用游标卡尺(精度0.001 mm)测算叶厚(cm)。使用HP(M162a)扫描仪扫描叶片,运用PhotoShop 7.0计算叶面积(cm2)。运用电子天平(精度0.0001g)称量鲜叶重量后,于黑暗环境下4℃水中浸泡叶片12h,取出后用吸水纸迅速吸取叶表水分,称量饱和叶鲜质量[17];将叶片样品包好放入烘箱105℃杀青30min,75℃烘干至恒量,称叶干质量[19]。通过Excel计算比叶面积(m2/kg)、叶干物质含量(g/kg)、叶相对含水率(%),叶组织密度(mg/mm3),计算公式如下:
比叶面积=叶面积/叶干质量
(1)
叶干物质含量=叶干质量/叶饱和鲜质量
(2)
叶相对含水率=(叶鲜质量-叶干质量)/(叶饱和鲜质量-叶干质量)×100%
(3)
叶组织密度=叶干质量/(叶面积×叶厚)
(4)
将烘干称重后的芒萁叶研磨后进行养分的测定:叶全碳(g/kg)含量和叶全氮含量(g/kg)测定采用碳氮元素分析仪(Elemantar vario MAX,德国);叶全磷含量(g/kg)采用HSO-HClO4消煮定容过滤,提取上层清液后用连续流动分析仪(Skalar san++,荷兰)测定;叶全钾含量(g/kg)测定采用火焰分光光度仪(FP6140,中国)。
1.4.2土壤因子的测定
土壤物理性质测定指标为土壤含水率(%),采用方法为铝盒烘干法。土壤化学性质的测定包括pH值、土壤全碳含量(g/kg)、土壤全氮含量(g/kg)、土壤全磷含量(g/kg)、土壤全钾含量(g/kg)。其中pH值以水土比例2.5∶1,使用便携式酸度计(Starter 300)测定,元素测量方法与叶样品相同。
1.5 数据统计与处理
用Excel 2010和SPSS 20.0软件对所得数据进行统计分析。采用单因素(One-ANOVA)和Duncan法进行方差分析和多重比较(显著性水平α=0.01),用Pearson法分析芒萁叶功能性状各指标间的相关性,用主成分分析法(Principal Component Analysis, PCA)分析影响芒萁叶功能性状的主要指标,并采用冗余分析法(Redundancy Analysis, RDA)分析影响芒萁叶功能性状的主要土壤因子。利用Origin 2017和Canoco 4.8和Excel 2010软件制作图表。图表数据表达形式为平均值±标准差(M±SD)。
2 结果与分析
2.1 不同植被恢复年限下芒萁叶功能性状差异
如图2所示,随着植被恢复年限的增加,芒萁叶各功能性状指标表现出明显的差异(P<0.05)。其中叶厚、叶干物质含量和叶组织密度总体变化趋势为逐渐下降,极小值均出现于植被恢复80年地。比叶面积、叶绿素含量的变化总体表现为逐步上升,极大值均出现于植被恢复80年地。叶全碳含量、叶全氮含量和叶全磷含量总体变化趋势相似,表现为先上升再小幅回落。叶全碳含量与叶全氮含量极大值出现在植被恢复36年地,而叶全磷含量的极大值则出现于植被恢复18年地。叶面积的变化总体呈现倒U状趋势,极大值出现于植被恢复18年地,并由此逐渐减少,直至植被恢复80年地达到最小。叶全钾含量变化与叶面积总体相反,呈U状趋势,极小值出现在植被恢复18年地,而后逐渐上升,至植被恢复80年地达到最大。叶相对含水率变化波动剧烈,在植被恢复12年地之前的数据表现为逐渐下降,在植被恢复18年地逆向波动,并到达该组数据的极大值,而后又急剧下降,在植被恢复36年地达到该组数据的最小值,并在植被恢复80年地再次达到较高的水平。值得注意的是,在植被恢复6年地,芒萁叶功能性状中叶厚、叶面积、比叶面积、叶干物质含量、叶组织密度、叶绿素含量、叶全氮含量、叶全磷含量均表现出相较于趋势逆向波动的特征。这些现象与长汀所采用的植被恢复技术有关,在恢复植被的前3年,当地依据退化地的土壤肥力水平施加复合肥,以提高土壤养分,促进恢复植被的存活与生长,从而影响到该样地芒萁叶功能性状指标的趋势变化。

图1 不同植被恢复年限下芒萁叶功能性状Fig.1 Characteristics of leaf function of Dicranopteris dichotoma under different vegetation restoration yearsa、b、c、d不同字母表示差异显著(P<0.05)
2.2 芒萁叶功能性状间相关性及主成分分析
2.2.1芒萁叶功能性状间相关性分析
由芒萁叶功能性状间相关性分析表明(表2),叶厚与叶干物质含量、叶组织密度在(P<0.01)水平表现出明显正相关,叶厚与比叶面积、叶绿素含量、叶全氮含量、叶全磷含量在(P<0.01)水平表现为明显负相关。叶面积与叶全磷含量的正相关关系表现为在(P<0.01)水平极显著,与叶组织密度的负相关关系表现为在(P<0.01)水平极显著。比叶面积与叶绿素含量、叶全氮含量、叶全碳含量、叶全磷含量在(P<0.01)水平表现出明显正相关,与叶干物质含量、叶组织密度在(P<0.01)水平表现为明显负相关。叶干物质含量与叶组织密度的正相关关系在(P<0.01)水平上极显著,与叶绿素含量、叶全氮含量和叶全磷含量的负相关关系在(P<0.01)水平上极显著。叶组织密度与叶全氮含量、叶全碳含量、叶全磷含量在(P<0.01)水平上呈极显著负相关。叶绿素含量与叶全氮含量在(P<0.01)水平上呈极显著正相关。叶全氮含量与叶全碳含量、叶全磷含量在(P<0.01)水平上呈极显著正相关。
2.2.2芒萁叶功能性状主成分分析
据表3所示,各芒萁叶功能性状指标的公因子方差均有较大分值,其中具有最小公因子方差的功能性状指标为叶全磷含量,为0.825,表明在因子信息归类过程中因子信息损失较小。以特征值>1为原则,提取3个主成分,其中特征值分别为6.373、2.069和1.433。这3个主成分贡献率分别为57.937%、18.806%和13.025%,累积贡献率为89.769%,>85%,说明这3个主成分因素可以代表芒萁叶功能性状变化的主要因素。依据初始因子的旋转成份矩阵得到各功能性状指标综合得分,其得分大小表明各个功能性状指标对主成分的贡献情况。通过对综合得分位次进行排名可知,叶全氮含量>比叶面积>叶全磷含量>叶绿素含量>叶面积>叶有机体碳>叶全钾含量>叶相对含水率>叶组织密度>叶厚>叶干物质含量,指标因子排名越靠前表明其在芒萁叶功能性状各指标中占据更重要的比重。

表2 芒萁叶功能性状相关性系数
**P< 0.01 ; *P<0.05;叶厚 LT Leaf thickness; 叶面积 LA Leaf area; 比叶面积 SLA Specific leaf area;叶干物质含量 LDMC Leaf dry matter content; 叶组织密度 LTD Leaf tissue density; 叶相对含水率 LRWC Leaf relative water content;叶绿素含量 LCC Leaf chlorophy content; 叶全氮含量LTN Leaf total N content;叶全碳含量 LTC Leaf total C content;叶全磷含量 LTP Leaf total P content; 叶全钾含量 LTK Leaf total K content

表3 初始因子旋转成份矩阵与主成分贡献率
2.3 壤因子随植被恢复年限变化差异
通过对比不同植被恢复年限下土壤因子的差异(P<0.05)可知(表3),土壤全碳含量总体变化趋势为随着植被恢复年限的增加而呈不断上升趋势,但在植被恢复12年地(14.118 g/kg)出现相对峰值,较植被恢复18年地(12.62 g/kg)更大。土壤全氮含量也表现出相同规律,植被恢复12年地(0.834 g/kg)的土壤全氮含量大于植被恢复18年地(0.724 g/kg),土壤全磷含量的变化也表现为随植被恢复年限的变化而不断增加的趋势,峰值出现于植被恢复36年地(0.336 g/kg),而后略有下降。土壤全钾含量在不同植被恢复年限下变化有所波动,总体呈上升趋势,极大值出现于植被恢复36年地(3.878 g/kg),而后略有下降。土壤pH值与土壤含水率随植被恢复年限的变化表现不明显,土壤pH值最大值出现在植被恢复0年地(4.483),最小值出现于植被恢复36年地(4.16)。土壤含水率最大值出现在植被恢复80年地(9.764%),最小值出现于植被恢复18年地(6.731%)。总体而言,土壤养分含量随植被恢复年限的增加表现为不断增加的趋势,土壤含水率则因地点不同表现出较大差异。

表4 不同植被恢复年限土壤因子变化差异
同列不同字母表示差异显著(P<0.05) ;土壤全碳含量STC Soil total C content;土壤全氮含量STN Soil total N content; 土壤全钾含量STK Soil total K content; 土壤全磷含量STP Soil total P content; 土壤含水率SMC Soil moisture content
2.4 芒萁叶功能性状与土壤因子的关系
本文通过线性约束性RDA排序,分析芒萁叶功能性状对土壤因子的响应关系。结果显示(表4),RDA排序轴1和排序轴2的特征值分别为0.744和0.042,累积解释百分率为78.7%,总体解释百分率达到79.2%,可反映叶功能性状与土壤因子的关系。排序轴1主要反映了土壤全碳含量、土壤全氮含量、土壤全磷含量和土壤含水率对芒萁叶片功能性状的影响。排序轴2主要反映了土壤全钾含量和土壤pH对叶片功能性状的影响。从RDA排序图可以看出,各叶功能性状指标对土壤因子的响应不同,且程度也有差异。比叶面积、叶全氮含量、叶绿素含量、叶全磷含量与土壤全碳含量、土壤全氮含量和土壤全磷含量存在较强的正相关关系,对土壤pH呈负相关关系,但相关程度相对较弱。由此可知,土壤全碳含量、土壤全氮含量和土壤全磷含量是影响芒萁叶主要功能性状的关键土壤因子(图2)。

表5 芒萁叶功能性状与土壤因子的RDA排序
LFT:叶功能性状 Leaf functional traits; SF:土壤因子 Soil factors
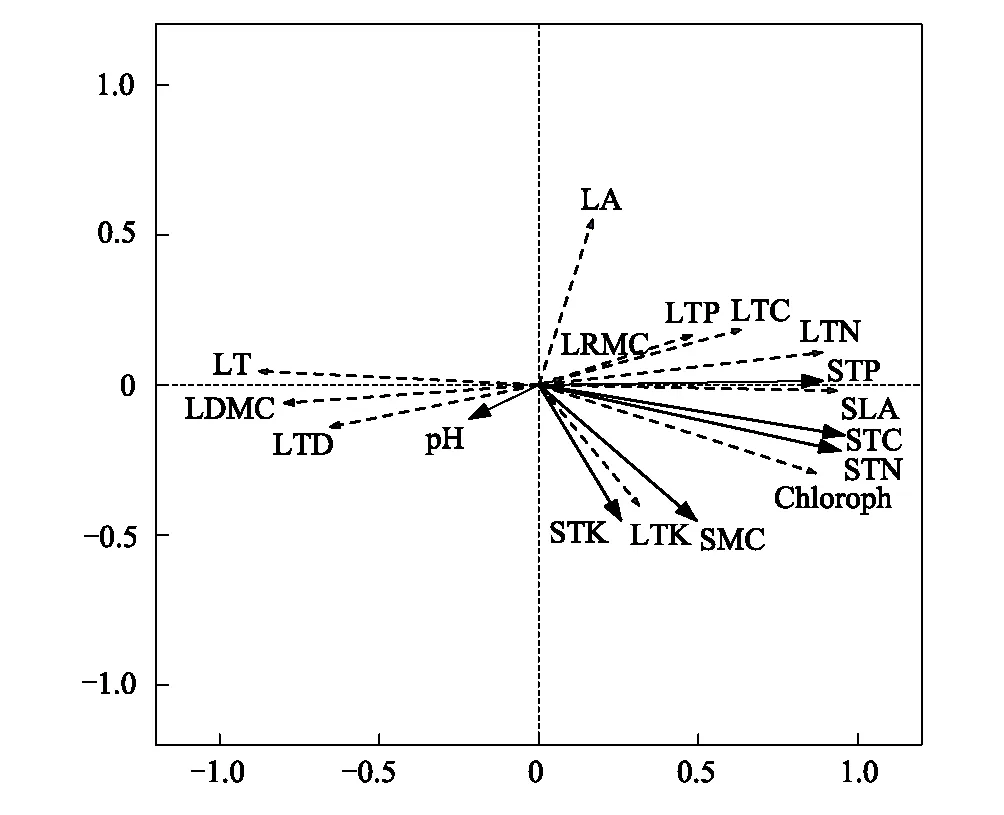
图2 芒萁叶功能性状与土壤因子的RDA排序 Fig.2 RDA sorting of functional traits and soil factors in leaves of Dicranopteris dichotoma
3 结论与讨论
3.1 不同植被恢复年限芒萁叶功能性状变化规律
不同植被恢复年限代表了土壤侵蚀区由次生裸地至演替顶级之间生态环境变化的每个阶段,生境条件随演替产生较大改变,进而对芒萁叶片的功能性状产生影响[20-21]。叶厚、叶组织密度和叶干物质含量分别体现出植物体的自我保护机制和养分分配与保持能力[20, 22-23]。本研究发现,叶厚、叶组织密度、叶干物质含量在0年地均处于高水平,在80年地处于低水平,在植被恢复初期,地表裸露,阳光直射地表,致使近地面温度高,蒸发量大,芒萁叶片通过增加厚度锁住水分,提高叶片组织密度以减缓植物体生长速率,进一步防止叶片失水过多和抵御高温伤害,叶干物质含量较高也说明在这一阶段芒萁具有较高的养分保持能力,锁住植物体内的营养成分减少流失维持生命过程;随着植被恢复年限的增加,叶厚、叶组织密度和叶干物质含量均趋于减小,这也说明在生态恢复过程中,水热、养分条件不断改善。芒萁通过降低叶组织密度和叶干物质含量提升生长速率,最大限度获取生存空间。比叶面积反映了植物体投入单位质量的干物质所获得的捕光面积[24-25]。伴随着植物叶片干物质含量的不断增加,芒萁比叶面积相对减小,在植被恢复初期生境资源有限、芒萁通过降低比叶面积,提高叶干物质含量减缓代谢及生长速率,因此比叶面积在未治理对照地出现极小值的特征,随着植被恢复年限增长,生境条件转好,芒萁又通过提高比叶面积,降低叶干物质含量,提升植物体生长代谢速率,在物种竞争激烈的环境条件下获取更多生存资源。
植物对环境的适应也体现于碳收获的最大化[26-27]。在植被恢复年限的增长过程中,芒萁叶片通过吸收大气中的CO2形成光合产物并作为生物量存储于叶片中,伴随植被恢复年限增加,环境因素趋好,叶片固碳能力也随之增强。叶绿素含量常被作为植物氮含量的诊断指标,叶绿素含量直接决定着植物光合能力及氮素含量的高低[28]。在植被恢复初期,芒萁叶片氮、磷含量极度缺乏,进而导致叶片叶绿素含量低,生长状况差;随着植被恢复年限的增长,叶片氮、磷含量不断增加,叶绿素含量提高,芒萁生长状况也明显改善。
在不同植被恢复年限下芒萁叶片各功能性状组合表现出不同特征,在植被恢复0年地自然环境恶劣的背景下,芒萁通过调整叶片功能性状来应对恶劣的生存环境,表现出胁迫型的组合状况。在植被恢复18年地各形态功能特征达到一个峰值,该阶段生境条件有了很大改善,且其他物种还未大范围侵入,芒萁种具有较强的生存优势,其功能性状表现出拓展型的组合状况。随着植被恢复进入后期阶段,生态演替趋于顶级,物种多样性增强,林下分层明显,各层级物种均处于相互抑制的状态,芒萁叶片各形态功能特征趋于下降,表现出竞争型的组合状况。
3.2 芒萁叶功能性状相关性与主成分分析
植物叶功能性状间关系密切,通过一系列性状组合的调整和平衡以适应特定的生境[5]。在本研究中,芒萁比叶面积与叶厚呈极显著的负相关,因为这两项功能性状指标都与植物的干旱胁迫有关。较大的叶厚可阻碍叶片与大气进行热量交换,减缓水汽和CO2进出叶片的速率,而比叶面积则主要影响着叶片构建过程中资源的分配,因而比叶面积较小的植物往往具有更大的叶厚[5,20]。这一结果与施宇等[27]对黄土丘陵地区植被的研究结果一致,反映出芒萁为适应水分胁迫在功能性状上与其他植物具有相似表现。与此同时,比叶面积和叶干物质含量作为植物适应环境的关键叶性状,综合反应了植物利用资源的能力[29]。本研究中,芒萁比叶面积与叶干物质含量呈极显著负相关,这与周济源等[30]对淮北相山优势植物种的研究结论一致,并说明芒萁可维持自身生长与体内养分循环间的平衡。本研究表明,比叶面积减小,叶体积也随之减小,在水分不足时叶干重减小,导致叶组织密度增大,体现芒萁通过比叶面积和叶组织密度共同作用以维持体内水分的平衡[5]。
氮与叶绿素的合成密切相关[28,31],因而叶全氮含量与叶绿素含量呈极显著的正相关关系,这与宋彦涛等[28]的研究一致。在植物体内碳被称为植物有机体的“骨架”[32],是构成生物大分子结构的基本元素,且为氮、磷的协同元素,他们之间一般呈正相关关系[20]。在本研究中,芒萁叶氮-叶碳、叶氮-叶磷均具有较强的正相关关系,且叶片碳、氮、磷含量与叶厚、比叶面积、叶干物质含量、叶组织密度同样具有显著相关性,这充分说明养分对于植物形态构建的重要作用。
通过对芒萁叶功能性状主成分分析结果的探讨,在对芒萁叶功能性状的研究评价过程中,应着重考虑综合得分较高的指标因子。在本研究中,叶全氮含量、比叶面积和叶全磷含量作为综合排名前三的指标因子可以被视为影响芒萁叶功能性状最重要的指标。比叶面积突出反映植物获取环境资源能力的高低[30],氮和磷影响植物生理过程及碳的分配[20],其共同表征出芒萁在不同植被恢复年限下叶片为适应环境而表现出的不同表型性状的能力,成为影响不同植被恢复年限下芒萁群落结构与性质最重要的叶功能性状。
3.3 芒萁叶功能性状与土壤因子的关系
随着植被恢复年限的增加,土壤因子随之改变,进而引发植物体对环境的适应策略及资源的利用方式也发生改变[26]。根据RDA分析结果表明不同植被恢复年限下芒萁叶主要功能性状指标均受土壤全碳含量、土壤全氮含量、土壤全磷含量的影响。其中,土壤全碳含量影响最大,其他因素影响相对较小。植被恢复年限的增加影响着土壤中碳、氮、磷含量的变化。其中,叶厚、比叶面积、叶干物质含量、叶组织密度及叶全碳含量与土壤全碳含量均有密切关系。旺盛的植物生命生产过程会增加植物体的固碳作用,生产更多的有机物质和产生更多的生物量,植物生产的有机物最终以植物凋落物的形式输入土壤被微生物分解,因而使得土壤全碳的含量产生差异,进而影响植物叶功能性状的变异[26-27]。比叶面积、叶全氮含量、叶绿素含量与土壤全氮含量直接相关。根据刘冬伟等[33]研究表明,生态系统氮循环主要受生物因素调控。植物通过吸收土壤氮一部分转化为氨基酸,合成蛋白质,另一部分与碳、氢、氧离子结合形成叶绿素[34]。凋落的植物体经微生物分解又以铵盐的形式重新输入土壤当中,从而成为土壤氮元素的重要来源[29]。旺盛的植被生长会增加氮需求,从而供应植物各器官功能,芒萁比叶面积的变化正体现出不同土壤条件下芒萁叶片资源配置和养分保持能力的不同。土壤全磷含量与叶绿素含量、叶全氮含量和叶全磷含量有密切关系。磷是植物生命活动的重要元素[35]。在植物细胞中,液泡与细胞质中稳定的磷浓度对于保持叶片的正常光合有重要作用[36-37]。土壤为植物体磷素的主要提供者,不同植被恢复年限下土壤磷含量的高低直接影响芒萁对磷的吸收,进而影响芒萁生命过程与叶功能性状的变化。
