睡莲种质资源遗传多样性分析及DNA指纹图谱构建
2020-03-23苏群杨亚涵田敏卜朝阳毛立彦张进忠潘介春卢家仕
苏群 杨亚涵 田敏 卜朝阳 毛立彦 张进忠 潘介春 卢家仕


摘 要:利用ISSR分子标记技术对从国内外引种的46份睡莲种质资源进行遗传多样性分析,并对其中的原生种及原生变种构建DNA指纹图谱,旨在阐明其亲缘关系基础上,为睡莲分类鉴定、杂交育种、功能基因的挖掘和利用及图位克隆等提供理论依据。结果表明:从100条ISSR引物中筛选出10条多态性高、重复性好的引物,在46份睡莲种质资源中共扩增出281条带,平均每条引物扩增条带数为28.1条,多态性条带281条,多态性比例为100%;平均Shannon信息指数(I)为0.4197,平均Neis基因多样性指数(H)为0.2657,平均有效等位基因数(N)1.4139,表现出丰富的遗传多样性;遗传相似系数值在0.51~0.98之间,基于遗传相似系数建立了聚类树状图,揭示了46份睡莲种质资源的亲缘关系,并在相似系数水平为0.68时,可将所有供试材料聚为6类。对总计24个睡莲原生种及原生变种构建了DNA指纹图谱,依据图谱差异可鉴定不同的睡莲种质。
关键词:睡莲种质资源;ISSR分子标记;遗传多样性;DNA指纹图谱
中图分类号:Q949.746.1 文献标识码:A
Abstract: The genetic diversity of 46 waterlily germplasm resources introduced within China and abroad was analyzed by the ISSR molecular marker technology, and the DNA fingerprints of the original species and varieties were constructed in order to clarify the genetic relationship and provide a theoretical basis for the classification and identification of waterlily, hybrid breeding and functional gene mining, utilization and map-based cloning. The results showed that 10 primers with high polymorphism and good repeatability were screened out from 100 ISSR primers. A total of 281 bands were amplified from 46 waterlily germplasm resources, with an average of 28.1 bands per primer, 281 polymorphic bands and 100% polymorphic ratio. The average Shannon information index (I) was 0.4197, the average Nei's gene diversity index (H) was 0.2657, and the average effective allele number (N) was 1.4139, which showed abundant genetic diversity. The genetic similarity coefficient was between 0.51 and 0.98. Based on the genetic similarity coefficient, a clustering tree was constructed to reveal the genetic relationship of the 46 waterlily germplasm resources. When the similarity coefficient was 0.68, all the tested materials could be clustered into six groups. DNA fingerprints were constructed for 24 original species and varieties of waterlily, and different waterlily germplasms could be identified according to the differences of DNA fingerprints.
Keywords: waterlily germplasm resources; ISSR molecular markers; genetic analysis; DNA fingerprintings
睡蓮为睡莲科(Nymphaeaceae)睡莲属(Nymphaea L.)多年生草本宿根花卉。全世界睡莲属植物有50余种,主要分布在热带、亚热带和温带地区[1]。我国有5种[2],也有的报道是4种[3]。睡莲属可分为5个亚属:广热带睡莲亚属(Brachyceras)、广温带睡莲亚属(Nymphaea)、古热带睡莲亚属(Lotos)、新热带睡莲亚属(Hydrocallis)及澳大利亚睡莲亚属(Anecphya)[4]。作为水生花卉中的名贵花卉,睡莲以其丰富的花色、圣洁的寓意及茎叶对水中富营养物和有害物质的强吸附能力,深受人们的喜爱。此外,睡莲花朵可泡茶,叶柄和花柄可食用,全草宜作绿肥,又可入药,具有极高的食用价值和观赏旅游、水景造景价值[5]。
睡莲种质资源是发展睡莲鲜切花、食用生产的物质基础,也是睡莲研究领域尤其是睡莲新品种选育和生物技术研究的基础。我国对睡莲的研究虽处于起步阶段,但发展迅猛,现阶段对睡莲的研究主要集中在引种栽培[6-11],生殖繁育[12-13]、功能活性[14-17]、生理生化分析[18-20]以及新品种选育[21-23]等方面。对睡莲种质资源遗传多样性和分子图谱的研究则少有报道,因此,开展睡莲种质资源的遗传性状分析和DNA指纹图谱构建,有利于充分利用睡莲种质资源和发掘、创制出性状优异的睡莲新种质,对睡莲育种研究和创新具有十分重要的意义。
ISSR(Inter-Simple Sequence Repeat)分子标记技术是一种基于PCR的新兴DNA分子标记技术。与其他分子标记相比,ISSR分子标记具有很大的优越性,如试验操作简单、成本低、快速灵敏、稳定性高、所需DNA量少等,而且不需要预先知道研究对象的基因组序列,大大减少了多态性分析的预备工作[24-25]。近年来,ISSR分子标记在其他花卉,如杜鹃[26]、荷花[27]、樱花[28]、桂花[29]、风信子[30]及百合[31]等种质资源的鉴定与遗传多样性分析中得到了广泛应用。此外,ISSR分子标记技术在构建遗传图谱和指纹图谱尤其是背景种质狭窄的指纹图谱具有很大潜力[32]。本研究基于ISSR分子标记技术对从国内外引种的46份睡莲种质资源进行遗传多样性分析,并对其中的原生种及原生变种构建DNA指纹图谱,旨在阐明其亲缘关系基础上,为睡莲分类鉴定,杂交育种及功能基因的挖掘、利用,图位克隆等提供理论依据。
1 材料与方法
1.1 材料
本研究以46份睡莲种质为实验材料,其中含有原生种21个,原生变种3个,园艺品种22个。供试样本均种植于广西农业科学院花卉研究所睡莲种质资源圃。采摘完整的、无病虫害、未展开的沉水幼叶1~2片,浸没于装有纯净水的标记袋中4 ℃冰箱保存,样本顺序及编号见表1。
1.2 方法
1.2.1 DNA提取及检测 DNA提取采用TIANGEN新型植物基因组DNA提取试剂盒,在1%的琼脂糖凝胶中电泳检测DNA质量,用Gene Spec核酸检测仪检测DNA纯度和浓度。最后将样品浓度稀释为50 ng/μL,–20 ℃保存。
1.2.2 ISSR-PCR 扩增反应体系及程序 采用20 μL的反应体系,包含了2×Es Taq Master Mix(含染料)10 μL,供试睡莲模板DNA及ISSR引物各1 μL,8 μL的ddH2O补齐。PCR扩增程序参考Poczai等[33] 的方法并进行适当优化,具体为:94 ℃预变性1 min;94 ℃变性45 s,不同温度下退火45 s,72 ℃延伸2 min,共35个循环; 72 ℃延伸5 min,4 ℃保存?ISSR-PCR反应产物用1.8%的琼脂糖凝胶电泳,缓冲液为1TAE。
1.2.3 ISSR引物合成及筛选 根据加拿大哥伦比亚大学设计并公布的ISSR引物序列,由上海生物工程有限公司合成本实验需用的100条ISSR引物。预先用5个不同睡莲亚属供试样本进行初筛,最后选取10条重复性好、条带清晰、多态性高的引物用于本实验。
1.3 数据处理
根据ISSR图谱条带的有无,利用人工方法统计读带,将图谱中稳定出现且肉眼可见的电泳条带赋值为“1”,同一扩增水平位置无电泳条带赋值为“0”,将读带数据输入Excel表格,建立原始矩阵数据库,统计扩增产物的条带总数和多态性条带数。用POPGEN 1.32软件计算多态位点百分率、Neis基因多样性指数(H)、有效等位基因数(Ne)和Shannon多样性信息指数(I)。用软件NTSYS 2.10e计算遗传相似系数GS(genetic similarity),根据GS值按非加权配对算术平均法(UPMGA)进行聚类分析,建立聚类图。
2 结果与分析
2.1 引物扩增多态性及遗传多样性分析
2.1.1 引物扩增多态性分析 采用试剂盒提取的46份睡莲基因组DNA电泳结果见图1。10条ISSR引物在46份睡莲种质资源中共扩增出281条谱带,其中281条均具有多态性,占100%(表2)。单条引物扩增出的清晰条带数在23~35条之间,扩增位点数最多引物为UBC835,扩增出35个位点,最少为UBC845,有23个位点,平均多态性条带28.1条。可见ISSR检测睡莲属种质资源遗传多样性的效率很高,也表明睡莲属种质资源在分子水平上具有极为丰富的遗传多样性。引物UBC?841对46份睡莲种质基因组DNA的扩增结果如图2所示。
2.1.2 遗传多样性分析 等位基因数(Na)、有效等位基因數(Ne)、Shannon多样性信息指数(I)和Nei's基因多样性指数(H)均是衡量遗传多样性水平的常用指标,是目前应用较为广泛的遗传多样性指数。46份睡莲种质资源的平均Na、Ne、H和I值分别为2.000、1.4139、0.2657和0.4197(表3)。
2.2 睡莲种质资源的ISSR聚类分析
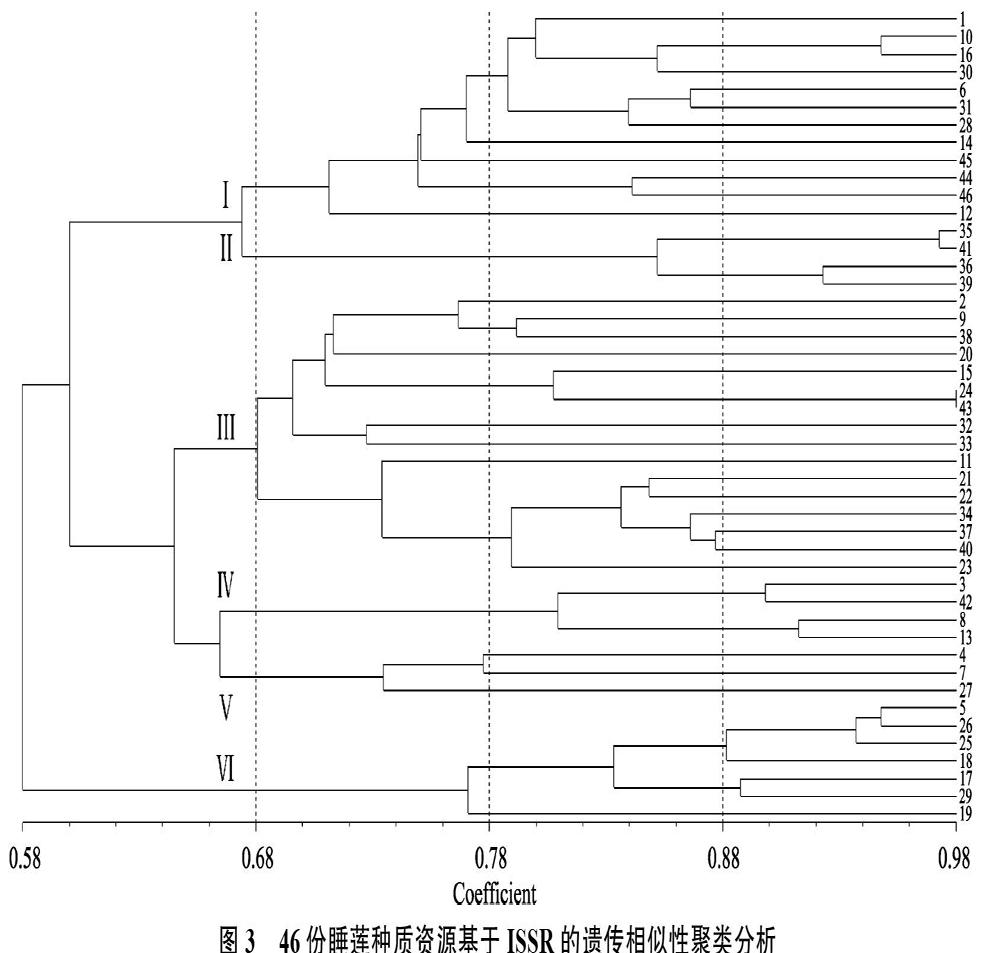
2.2.1 遗传相似系数 利用10个引物产生的标记信息,建立了供试样本的相似系数矩阵。遗传相似系数值越大,说明材料间的亲缘关系越近,反之,亲缘关系越远。46份睡莲种质资源的遗传相似系数在0.51~0.98之间,平均遗传相似系数为0.78。其中24号的白蓝星与43号蓝星种质之间的遗传相似系数最大为0.98,说明他们之间的亲缘关系最近,遗传差异相对较小。7号的秘鲁睡莲与35号的紫色幻想之间的遗传相似系数最小为0.51,说明他们之间的亲缘关系最远,遗传差异相对较大。
2.2.2 聚类分析 为了确定各供试材料间的遗传关系,通过软件NTSYS 2.10e对46份睡莲种质资源计算遗传相似系数GS(genetic similarity),根据GS值按非加权配对算术平均法(UPMGA)进行聚类分析,建立遗传关系聚类图(图3)。由图3可知,供试材料以遗传相似系数0.68为阈值时,可将46份睡莲种质资源聚为6类,第I类由属于广温带亚属的12份睡莲种质资源组成,其中的12号小白子午莲单独聚成一支;第II类由属于人工跨亚属杂交育成的跨亚属的4份睡莲种质资源组成,具体为35号紫色幻想与41号诗丽吉皇后聚在一起,36号暹罗紫与39号暹罗粉聚在一起。第II类的4个人工杂交的跨亚属品种,在遗传相似系数0.66时与第I类的广温带亚属聚在一起,说明相对于其他亚属,第II类人工杂交的跨亚属品种与广温带亚属亲缘关系较近。这在表观形态特征上也能体现,如上述跨亚属杂交的4个品种叶片呈椭圆或卵圆型,叶片边缘全缘,花朵浮于或略高于水面,其均是典型的广温带亚属睡莲具有的特征,但其花部特征尤其是雄蕊与广温带亚属睡莲差异较大;第III类由属于广热带亚属的16份睡莲种质资源组成,其中小花睡莲、21号暹罗皇后、34号华丽、37号约瑟芬及40号紫乔伊这5份睡莲种质资源聚在一起;第IV类由同属于澳洲亚属的4份睡莲种质
图3 46份睡莲种质资源基于ISSR的遗传相似性聚类分析
Fig. 3 Cluster analysis of 46 waterlily germplasm resources based on ISSR genetic identities
资源组成,具体为3号蓝巨睡莲和42号白巨睡莲聚在一起,8号变色澳洲睡莲与13号澳洲IM睡莲聚在一起;第V类由3种同属于新热带亚属的睡莲种质组成,其中27号具备花胎生能力的增殖睡莲单独聚为一支;第VI类由同属于古热带亚属的7份睡莲种质组成,其中19号喀麦隆睡莲单独聚为一支。
2.3 部分睡莲种质资源的DNA指纹图谱构建
用筛选出的10个ISSR引物对24个睡莲原生
种及变种构建DNA指纹图谱,通过对比发现引物UBC840和UBC841均可单独鉴别出24份睡莲原生种质资源。本研究利用UBC840构建了24份睡莲原生种质资源DNA指纹图谱(图4),其中黑色表示该位点有条带,即该位点0-1矩阵中为“1”,无色表示该位点无条带,即该位点在0-1矩阵中为“0”。构建的指纹图谱可用于24份睡莲原生种质资源的分类与鉴定。
图4 基于UBC840分子标记构建的睡莲原生种及原生变种的DNA指纹图谱
Fig. 4 DNA fingerprint of primordial species and varieties of waterlily based on UBC840 molecular markers
3 讨论
3.1 睡莲种质资源遗传多样性及亲缘关系分析
遗传多样性是生物所携带的遗传信息总和,是每种生物所固有的特性,一个种群遗传多样性越高或越丰富,在环境发生变化时生存下来的可能性就越大,越容易扩展其分布范围和开拓新环境[34-35]。ISSR分子标记具有扩增多态性高、稳定性好、操作简便等优点[36],现已被应用于热带睡莲的遗传关系分析、杂交种鉴定[33]、聚群内遗传变异及基因流分析中[37]。本研究通过反复试验,得到优化后的ISSR-PCR体系,筛选出10个引物
对46份睡莲种质资源进行了扩增,多态性位点百分率高达100%,Shannon信息指数(I)为0.4197,有效等位基因数(Ne)为1.4139,Neis基因多样性指数(H)为0.2657,表明睡莲种质资源具有极为丰富的遗传多样性,蕴涵着广泛的遗传变异,这也与张海平[38]通过研究睡莲的表观性状多样性及同工酶分析得出的结果一致,同时也说明ISSR分子标记可以较好地用于睡莲属的多样性分析。睡莲属植物开花习性表现为雌蕊先熟(protogyny)[1],这种性成熟时间自然隔离特点导致绝大部分睡莲属植物为异花授粉,致使睡莲属植物种间基因交流频繁,杂合度和多态性很高,可能是影响其种间在形态和分子遗传特性上的巨大差异的重要原因。实验结果显示,供试睡莲属种质资源的遗传相似系数为0.51~0.98,丰富的遗传多样性为睡莲杂交亲本选择和育种提供了重要的科学依据。从聚类图上看,在阈值0.78时,很多睡莲属种质资源单独聚成一支,说明其与同亚属的其他睡莲种质资源差异较大,亲缘关系较远,而且很多均是育性非常好的原生种质资源,如12号小白子午莲,32号米奴塔等,其育种潜力还未被完全开发,可作为亚属内或跨亚属杂交时的优良亲本选择。
睡莲属下分类现今依旧没有非常明确的标准。根据生态学特征,通常将睡莲分为耐寒睡莲(hardy waterlily)和热带睡莲(tropical waterlily)2大类[1],根据心皮的离生或聚合以及花柱的有无可将睡莲属植物分为2大组5个亚组[1, 9],根据地理来源可將睡莲属分为5或6个亚属[4, 39],即将睡莲属分为:广温带亚属、广热带亚属、古热带亚属、新热带亚属及澳洲亚属,后4个亚属通常简单称为热带睡莲。本研究以遗传相似系数GS值按非加权配对算术平均法(UPMGA)进行聚类分析,建立了46份睡莲种质资源的遗传关系聚类图,从聚类图上看,全部耐寒睡莲聚在一起构成单系类群,这与张海平[38]基于形态性状的聚类分析和刘艳玲等[40]基于ITS的睡莲属系统分类的研究一致,但热带睡莲分类地位与他们有较大出入。王海平[38]和刘艳玲等[40]认为,热带睡莲聚在一起构成单系类群,在其内部分成2支,一支是印度蓝睡莲、蓝星睡莲等昼开夜闭型睡莲,另一支则由印度红睡莲、埃及白睡莲为代表的古热带亚属组成的夜开昼闭型睡莲。本研究显示古热带亚属单独聚为一支,且位于聚类图基部,支持黄国振等[1]提出的认为晚上开花类型是比较原始的,相当于塔氏图示的第2阶段,白天开花类型相当于第4阶段,耐寒睡莲类型相当于第5、第6阶段的观点。此外从聚类图上看部分地理来源相同的睡莲种质资源聚在一起,如第IV类供试材料全部来源于澳洲,归属于澳洲睡莲亚属,具有较近的亲缘关系,支持根据地理来源的将睡莲属分为5个亚属的分类方式。
从聚类图上看,在阈值约为0.69时,可将第III类广热带亚属分为2支。其一支由9份非胎生能力的睡莲种质组成,另一支是由7份睡莲属种质聚在一起,其中5份具有胎生能力的睡莲种质聚在一起,说明具有胎生能力的睡莲种质遗传关系较近,表明它们很可能具有共同的起源。2份非胎生能力的睡莲种质分别是11号黄金国和23号苏皖娜,黄金国俗称黄色九品香水莲,是台湾园林部门在引种美国睡莲基础上培育而成[41-42],苏皖娜是由泰国著名睡莲育种家N. Nopchai Chansilpa博士选育出的著名洒金睡莲名品,这2个品种亲本来源复杂,之所以与具胎生能力睡莲种质聚为一支,很可能是其亲本中混有胎生能力的亲本祖先,致使其亲缘关系更靠近于具胎生能力的睡莲种质。
3.2 睡莲属原生种质资源分子指纹图谱
目前,我国在花卉品种资源生产和经营方面尚不规范,品种引种混乱或品种造假的现象时有发生,因此,开展花卉品种资源鉴定显得尤为重要[43]。传统的花卉品种资源鉴定方法主要依靠表型特征,虽然快速便捷,但是表型性状受环境影响较大,鉴别错误率较高,加之花卉品种资源逐年扩大,品种的相似度也越来越高,导致通过传统的表型鉴定方法越来越难[43]。DNA指纹图谱是指DNA样品用特定分子标记技术处理显示出具有特定DNA片段的总称[44]。DNA遗传物质由于受环境影响较小、多态性高,基于DNA扩增的PCR技术已经成为花卉品种鉴定最有效的方法[42, 45-46]。绝大部分睡莲原生种及其变种在未开花前非常相似,很难辨别,如蓝星和白蓝星,其叶型、叶色都极为相似,需开花后才可区分。少部分在开花条件下也很难区分开,如埃及白睡莲及其变种柔毛齿叶白睡莲,花色均为纯白色,锯齿状叶片,单从表观形态上很难区分开来,本研究通过筛选出的10个ISSR引物对24个睡莲原生种及原生变种构建DNA指纹图谱。分子身份证构建的目的是准确、快速和便捷地鉴别出不同种质的遗传信息,因此,在进行分子身份构建时,不仅要求标记位点的多态性要丰富,而且检测方法要操作简便、重复性强[47]。通过对比发现引物UBC840和UBC841均可单独鉴别出24份睡莲原生种质资源。本研究基于引物UBC840构建了24份供试睡莲原生种质的DNA指纹图谱,该图谱可作为相应睡莲原生种质资源鉴定和分类的依据,同时也为新种质和新品种的分类鉴定及知识产权保护提供分子技术水平上的保障。
参考文献
黄国振, 邓惠勤, 李祖修, 等. 睡莲[M]. 北京: 中国林业出版社, 2009: 1-10.
中国科學院中国植物志编辑委员会. 中国植物志[M]. 北京: 科学出版社, 1979(27): 8-12.
肖克炎. 中国野生睡莲的分布、现状和保护[J]. 人文园林, 2017(8): 34-35.
余翠薇, 陈煜初, 余东北, 等. 不同类型睡莲的特点及栽培技术[J]. 浙江农业科学, 2016, 57(10): 1694-1695.
廖卫伟, 杨志娟, 朱天龙, 等. 睡莲的植物学性状及其分类[J]. 现代农业科技, 2016(24): 148, 154.
陈发棣, 崔娜欣, 丁跃生. 南京地区新引耐寒睡莲主要观赏性状初步评价[J]. 上海农业学报, 2002, 18(3): 51-55.
李淑娟, 樊 璐, 原雅玲, 等. 关中地区引进耐寒睡莲品种观赏性状评价及优良品种筛选[J]. 陕西林业科技, 2010(3): 15-18.
吉建斌, 李清清, 黄娟英, 等. 京津冀耐寒睡莲品种引繁及栽培技术研究(1)——评分标准及生物学特性研究[J]. 林业实用技术, 2007(3): 3-5.
李尚志, 李国泰, 王 曼. 荷花·睡莲·王莲[M]. 北京: 中国林业出版社, 2002: 86-116.
赵福康. 不同施肥模式对香水莲生长发育的影响[J]. 安徽农业科学, 2015, 43(2): 115 -116.
屠 莉. 盆栽睡莲养护技术[N]. 中国花卉报, 2006-08-17.
戚华沙, 潘 梅, 符瑞侃, 等. 蓝药睡莲组织培养外植体消毒方法的研究[J]. 中国园艺文摘, 2016, 32(2): 38-39, 116.
孙春青, 戴忠良, 潘跃平. 睡莲不同杂交组合幼胚愈伤组织诱导研究[J]. 江西农业学报, 2014, 26(1): 49-52.
杨英豪. 重金属镉胁迫对睡莲生理生态效应的研究[D]. 南京: 南京农业大学, 2013: 1-40.
章志琴, 方弟安, 徐卫红, 等. 荷花和睡莲对景观水净化的效果研究[J]. 江苏农业科学, 2009(5): 320-322.
张卫明, 姜洪芳, 徐 辉, 等. 香水莲花降血脂作用的实验研究[J]. 食品工业科技, 2009(2): 111-112, 116.
车 璐, 吴晓琴, 郑茜茜, 等. 香水莲花抑制大鼠前列腺增生的试验研究[J]. 中国食品学报, 2015, 15(2): 28-33.
朱满兰, 王亮生, 张会金, 等. 耐寒睡莲花瓣中花青素苷组成及其与花色的关系[J]. 植物学报, 2012, 47(5): 437-453.
石 凝, 刘晓静, 杜凤凤, 等. 热带睡莲鲜花中挥发油成分的GC-MS分析[J]. 植物资源与环境学报, 2017, 26(4): 104-106.
徐 辉, 张卫明, 姜洪芳, 等. 香水莲花挥发油的气相色谱-质谱分析[J]. 食品研究与开发, 2008, 29(9): 101-103.
王秀英. 李子俊: 花香沁人 坚持最美[J]. 中国花卉园艺, 2017(3): 22-23.
李淑娟, 尉 倩, 张 昭, 等. 耐寒睡莲新品种‘天赐的选育[J]. 北方园艺, 2018(3): 208-210, 2.
柏 斌. 云南睡莲新品种‘粉月亮获国际登录[J]. 中国花卉园艺, 2017(16): 24.
孙 洪, 程 静, 詹克慧, 等. ISSR标记技术及其在作物遗传育种中的应用[J]. 分子植物育种, 2005, 3(1): 123-127.
刘志华, 于 卓, 董 鹏, 等. 罗布麻遗传多样性的ISSR分析[J]. 中国草地学报, 2009, 31(5): 96-101.
徐静静. 杜鹃花遗传多样性及品种分类研究[D]. 杨凌: 西北农林科技大学, 2017.
何天友, 沈少炎, 瞿印权, 等. 基于ISSR标记的20种荷花品种资源遗传多样性分析[J]. 安徽农业科学, 2017, 45(6): 128-132.
蒋冬月, 沈 鑫, 吴 帆, 等. 基于ISSR标记分析浙江省樱花种质资源的遗传多样性[J]. 浙江林业科技, 2018, 38(4): 1-7.
罗仙英, 桂敬飞, 严 治, 等. 贵州桂花种质资源遗传多样性的ISSR分析[J]. 种子, 2017, 36(6): 62-66.
胡凤荣, 胡月苗, 王 斐, 等. 利用 ISSR分子标记分析29个风信子品种的遗传多样性[J]. 分子植物育种, 2015, 13(2): 379-385.
侯 珺, 罗建让, 肖菲菲, 等. 21种野生百合亲缘关系的ISSR分析[J]. 广西植物, 2016, 36(9):1032-1038.
刘威生, 冯晨静, 杨建民, 等. 杏ISSR反应体系的优化和指纹图谱的构建[J]. 果树学报, 2005, 22(6): 626-629.
Poczai P, Hyvonen J, Matyas K K, et al. Genetic variability of thermal nymphaea (Nymphaeaceae) populations based on ISSR markers: Implications on relationships, hybridization, and conservation[J]. Plant Molecular Biology Reporter, 2011, 29(4): 906-918.
付晨熙, 方 明, 朱友林, 等. 直立百部遺传多样性的ISSR分析[J]. 西北植物学报, 2012, 32(8): 1553-1559.
中国科学院生物多样性委员会. 生物多样性研究的原理与方法[M]. 北京: 中国科学技术出版社, 1994: 141-165.
陈志丹, 林志坤, 孙威江, 等. 125份武夷山茶树种质资源的遗传多样性分析及DNA指纹图谱的建立[J]. 分子植物育种, 2016, 14(12): 3601-3608.
Chen Y Y, Fan X H, LI Z, et al. Low level of genetic variation and restricted gene flow in water lily Nymphaea tetragona populations from the Amur River[J]. Aquatic Botany, 2017, 140: 55-61.
张海平. 部分睡莲属植物的形态多样性及同工酶分析[D]. 南京: 南京农业大学, 2008.
Primlarp W, Komgrit C. Thai nationality waterlily in PANG U BON[M]. Thailand: Plusprass Co. Ltd, 2018.
刘艳玲, 徐立铭, 倪学明, 等. 睡莲科的系统发育:核糖体DNA ITS区序列证据[J]. 植物分类学报, 2005, 43(1): 22-30.
黄家祥. 四季开花—香水莲[J]. 北方园艺, 2005(3): 44.
李红斌. 台湾九品香水莲特征特性及栽培技术[J]. 上海蔬菜, 2015(4): 31-32.
葛亚英, 张 飞, 沈晓岚, 等. 丽穗凤梨ISSR遗传多样性分析与指纹图谱构建[J].中国农业科学, 2012, 45(4): 726- 733.
Vos P, Hogers R, Bleeker M, et al. AFLP: a new technique for DNA fingerprinting[J]. Nucleic Acids Research, 1995, 23(21): 4407-4414.
徐宗大, 赵兰勇, 张 玲, 等. 玫瑰SRAP遗传多样性分析与品种指纹图谱构建[J]. 中国农业科学, 2011, 44(8): 1662-1669.
缪恒彬, 陈发棣, 赵宏波, 等. 应用ISSR对25个小菊品种进行遗传多样性分析及指纹图谱构建[J]. 中国农业科学, 2008, 41(11): 3735-3740.
李国田, 张美勇, 相 昆, 等. 基于ISSR标记的16个核桃品种遗传多样性分析及分子身份构建[J]. 核农学报, 2015, 29(10): 1884-1892.
