不同训练负荷对大鼠骨骼肌超微结构及线粒体功能的影响
2020-03-20任鹤菲于昕仪陈立军张馨予于利人
任鹤菲,于昕仪,陈立军,张馨予,雷 静,陈 健,于利人
不同训练负荷下,骨骼肌的结构和功能会发生明显的变化,肌原纤维和三联体结构出现不同程度的改变,影响收缩和能量供应系统[1]。骨骼肌供能主要依靠线粒体呼吸链,线粒体呼吸链酶复合体Ⅰ、Ⅱ、Ⅲ、Ⅳ的表达水平与线粒体呼吸及能量代谢功能密切相关[2]。骨骼肌供能的同时产生超氧化产物,线粒体超氧化物歧化酶(superoxide dismutase, SOD)可及时清除超氧化产物并修复细胞损害[3, 4]。如何科学训练才能保护骨骼肌结构,最大限度发挥线粒体功能需要进一步探讨。本实验检测4周训练后大鼠骨骼肌超微结构及线粒体功能的变化,探讨训练对骨骼肌结构和能量代谢的影响及保护机制,为科学训练提供实验依据。
1 材料与方法
1.1 实验动物 健康清洁级雄性SD大鼠24只,鼠龄2个月,体质量(214±11) g,由北京大学医学部实验动物科学部提供,许可证号SCXK2002-0001,饲养环境温度维持在21~25 ℃,湿度范围40%~60%,照明按自然昼夜供给,进食和饮水自由。本实验中对动物的处理方法符合《关于善待实验动物的指导性意见》。
1.2 训练模型的建立 实验大鼠先进行适应性饲养1周,然后随机分为正常对照组、有氧训练组和无氧训练组,每组8只,组间动物体质量比较,差异无统计学意义。进行3 d适应性跑台训练(坡度为0°,速度8.2 m/min,10 min/d)后,实验参照Bedford等[5]的方法建立动物模型。参考Shepherd等[6, 7]的方法确定训练强度和分组:训练强度在64%~76%最大摄氧量(maximal oxygen uptake, VO2max)确定为有氧训练组,训练强度>80% VO2max确定为无氧训练组。有氧训练组采用递增负荷训练,以15 m/min为起始速度,速度逐渐增快,第5分钟增至18 m/min,第10分钟增至20 m/min并保持恒定不变,同时增加坡度至5°,计时60 min。无氧训练组以50 m/min为起始速度,训练6 min后休息5 min,此过程重复3次。两组在训练过程中始终保证训练强度恒定。训练4周,每周保证在同一时间段开展训练,时间每周不少于6 d。对照组大鼠于笼内正常生活,不做任何额外处理。
1.3 实验方法
1.3.1 组织匀浆的制备 大鼠训练后立刻处死,迅速取其腓肠肌,用介质Ⅰ(0.12 M KCl,20 mM Hepes,5 mM MgCl2,1 mM EDTA,5 mg defatted BSA/ml pH 7.4)冲洗,剔除脂肪及结缔组织并剪碎,加10 ml介质Ⅰ。电动匀浆器匀浆,1200 r/min,上下3次。600 r/min离心10 min。弃沉淀,上清液于17 000 r/min离心10 min。沉淀用介质Ⅰ悬浮,7000 r/min离心10 min。沉淀悬浮在20 ml介质Ⅱ(0.3M Sucrose,2.0mM Hepes,0.1 mM EDTA,2 mg defatted BSA/ml pH 7.4)中,3500 r/min离心10 min。沉淀悬浮于0.3 ml介质Ⅱ中。以上操作均在0~4 ℃下进行。线粒体蛋白含量用考马斯亮蓝法测定,采用可见-紫外分光光度计280 nm下校正的BSA作标准。
1.3.2 线粒体呼吸链酶复合体活性的测定 参照上海杰美基因医药科技有限公司试剂盒说明书测定。分光光度法分别在340 nm(酶复合体Ⅰ)、600 nm(酶复合体Ⅱ)和550 nm(酶复合体Ⅲ和Ⅳ)处测定,以牛血清白蛋白为标准,应用Folin-酚法测定各组线粒体蛋白浓度。计算公式:复合体酶比活性=[(△A样品-△A背景)×样品稀释倍数]/[0.1(样品质量,mg)×吸光系数×1(反应时间,min)]/线粒体蛋白浓度(mg/ml)。
1.3.3 总超氧化物歧化酶、锰超氧化物歧化酶和铜锌超氧化物歧化酶活性的测定 采用比色法测定总超氧化物歧化酶(total superoxide dismutase, T-SOD)、锰超氧化物歧化酶(manganese superoxide dismutase, Mn-SOD)和铜锌超氧化物歧化酶(copper zincsuperoxide dismutase, CuZn-SOD)活性,试剂盒为南京建成生物工程研究所产品,严格按照试剂盒说明操作。
1.3.4 透射电镜观察骨骼肌线粒体的超微结构 大鼠处死后迅速取腓肠肌,取1 mm2的骨骼肌组织用2.5%戊二醛孵育过夜。1%锇酸后固定,梯度乙醇脱水后氧化丙烯浸透,环氧树脂包埋,包埋好的样品制成超薄切片,醋酸铅铀双重染色,进行电镜样品制备。Philips EM400ST透射电镜观察骨骼肌线粒体超微结构,采集图像。

2 结 果
2.1 不同训练条件下骨骼肌形态及线粒体分布 在正常对照组中,肌浆内含大量与细胞长轴平行排列的肌原纤维,相邻肌原纤维间可见线粒体,每条肌原纤维上有明暗相间的带(图1A)。肌原纤维由大量平行排列的粗、细肌丝组成,可见粗、细肌丝横断面;肌膜下的肌浆中可见电子密度较高的线粒体(图1B)。在有氧训练组中,与正常对照组相比,肌原纤维排列更加规则,肌原纤维增粗,肌浆网、横小管发达,构成明显的三联体结构,线粒体数量增多(图1C、D)。在无氧训练组中,与正常对照组相比,肌浆中,肌原纤维之间可见大量发生肿胀的线粒体,电子密度较正常染色体低;肌原纤维结构发生变化,明带与暗带界线不清,明带中Z线不明显,暗带中H带与M线不明显,提示粗细肌丝排列紊乱(图1E、F)。

图1 不同训练负荷下大鼠骨骼肌线粒体超微结构透射电镜特点
A.正常对照组骨骼肌纵切面,Bar=0.5 μm;B.正常对照组骨骼肌横切面,Bar=0.5 μm;C.有氧训练组骨骼肌纵切面,Bar=1 μm;D.有氧训练组骨骼肌纵切面,Bar=0.5 μm;E.无氧训练组为骨骼肌纵切面,Bar=1 μm;F.无氧训练组为骨骼肌纵切面,Bar=0.5 μm
2.2 线粒体呼吸链酶复合体Ⅰ、Ⅱ、Ⅲ、Ⅳ活性变化 训练4周后,有氧训练组大鼠线粒体呼吸链酶复合体Ⅰ、Ⅱ、Ⅲ、Ⅳ活性水平均高于正常对照组和无氧训练组,无氧训练组大鼠线粒体呼吸链酶复合体Ⅰ、Ⅱ、Ⅲ、Ⅳ活性水平较正常对照组明显降低,差异均有统计学意义(P<0.05,表1)。
2.3 T-SOD,Mn-SOD和CuZn-SOD活性测定 训练4周后,有氧训练组大鼠线粒体T-SOD、CuZn-SOD和Mn-SOD水平高于正常对照组和无氧训练组,无氧训练组大鼠线粒体T-SOD、Mn-SOD水平较正常对照组明显降低,差异有统计学意义(P<0.05,表2)。CuZn-SOD活性与正常对照组相比,差异无统计学意义。
表1 大鼠线粒体呼吸链酶复合体活性
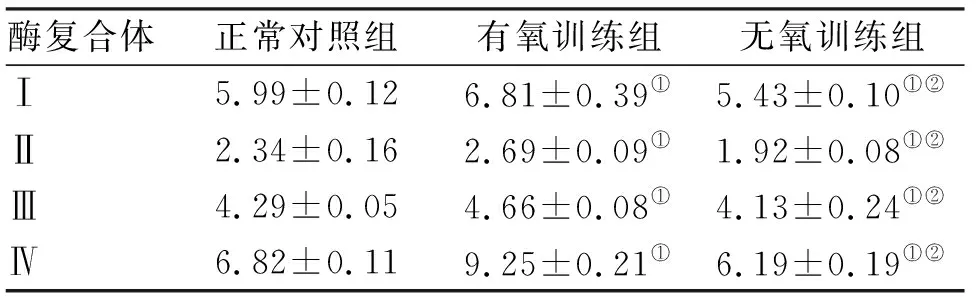
酶复合体正常对照组有氧训练组无氧训练组Ⅰ5.99±0.126.81±0.39①5.43±0.10①②Ⅱ2.34±0.162.69±0.09①1.92±0.08①②Ⅲ4.29±0.054.66±0.08①4.13±0.24①②Ⅳ6.82±0.119.25±0.21①6.19±0.19①②
注:与正常对照组相比,①P<0.05;与有氧训练组相比,②P<0.05
表2 T-SOD, Mn-SOD和CuZn-SOD活性
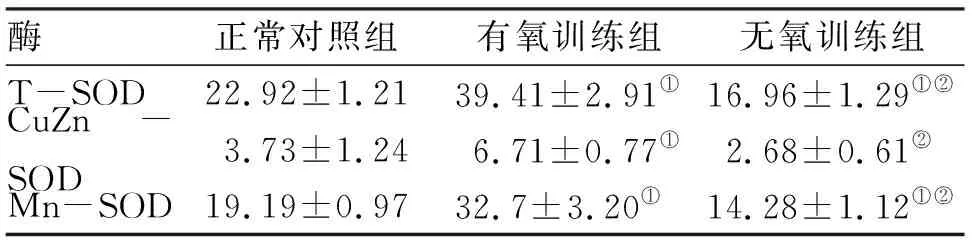
酶正常对照组有氧训练组无氧训练组T-SOD22.92±1.2139.41±2.91①16.96±1.29①②CuZn-SOD3.73±1.246.71±0.77①2.68±0.61②Mn-SOD19.19±0.9732.7±3.20①14.28±1.12①②
注:与正常对照组相比,①P<0.05;与有氧训练组相比,②P<0.05
3 讨 论
不同训练负荷可以诱发骨骼肌发生相应的适应性改变,无氧训练超过机体承受能力会导致骨骼肌产生暂时性的生理机能减退现象,即会发生训练疲劳。本实验中,电镜下观察骨骼肌超微结构呈现不同程度的变化,有氧训练组大鼠骨骼肌肌原纤维排列更加规则,三联体结构明显。无氧训练组大鼠骨骼肌肌浆中,电子密度较正常染色体低,粗细肌丝排列紊乱,超微电镜结果显示,有氧训练可加强骨骼肌收缩能力,提高应激状态下骨骼肌能量供应,无氧训练则导致骨骼肌损伤。
不同训练负荷可诱导运动性疲劳,线粒体结构和功能的失调是运动性疲劳发生的重要潜在因素。高强度氧化应激可使线粒体通透转运孔道(mitochondrial permeability transition pore,mPTP)更易于开放,线粒体膜电位的缺失及mPTP的开放进一步诱导线粒体肿胀,进而影响骨骼肌的结构。线粒体反向电子传递理论指出,在应激缺氧状态下,大量电子在复合体Ⅱ处聚集无法向下传递,而当无氧训练终止时,大量氧气致使游离电子涌向复合体Ⅰ,导致活性氧产生,诱发机体损伤[8]。本实验结果显示,4周有氧训练后大鼠骨骼肌线粒体数量增多,线粒体呼吸链酶复合体活性显著增强,这可能与持续性的有氧训练使得骨骼肌增加对能源物质的需求有关。无氧训练组线粒体呼吸链酶复合体活性低于正常对照组和有氧训练组,肌原纤维之间可见大量发生肿胀的线粒体,这可能是因为无氧训练损伤了线粒体呼吸链,使得电子传递发生障碍。
不同训练负荷下机体可产生如氧自由基等训练副产物,自由基的细胞毒作用可引起膜脂质过氧化,导致细胞损伤[9]。自由基如不能得到及时有效的清除将导致机体氧化与抗氧化功能平衡的失调, 破坏组织细胞,影响生物体行为训练能力及机体正常的氧化代谢功能[10, 11]。正常机体内存在以SOD为代表的自由基清除体系,其活力可以反映机体清除自由基的能力[12]。SOD以锰或铜和锌等作为辅因子,两者均是自由基的清除酶,是维持细胞正常代谢和生长必不可少的抗氧化物质[13]。过氧化氢是一类自由基,无氧训练可引起小鼠腓肠肌线粒体过氧化氢含量明显增加[14]。Mn-SOD基因的启动子通过与过氧化氢相结合,使得Mn-SOD基因转录水平显著提高[15],有利于提高自由基的清除能力。本实验结果显示,有氧训练组T-SOD、Mn-SOD和CuZn-SOD水平均高于正常对照组和无氧训练组。说明有氧训练提高能量代谢能力显著高于无氧训练,能通过进一步提高SOD的表达水平,增加超氧阴离子自由基的清除,使得抗氧化能力得到了增强。高强度的无氧训练由于线粒体呼吸链电子传递发生障碍,还原型辅酶Ⅰ得不到氧化而堆积,导致氧自由基生成增加,破坏了机体氧化与抗氧化功能的平衡,进而影响线粒体的呼吸及氧化代谢功能。此外,实验发现,无氧训练并不能显著降低CuZn-SOD水平,原因可能是因为无氧运动主要影响线粒体,而CuZn-SOD主要存在于胞浆中,受影响变化不明显。
综上所述,不同训练负荷对骨骼肌超微结构和线粒体功能影响不同,其机制复杂,对其进行深入的研究具有重要意义。有氧训练可以改善骨骼肌的能量代谢及抗氧化功能,增加细胞内线粒体数量及相关代谢酶活性,从而改善骨骼肌形态,提高机体抗疲劳能力,这与线粒体能量代谢及SOD等抗氧化功能密切相关。但对不同训练负荷引起骨骼肌结构和功能改变的机制,仍有待进一步研究。
