一个新的水稻无中叶脉突变体的表型分析和基因定位
2020-03-20黄仁良刘建华曹丰生
杨 宙,黄仁良,何 虎,刘建华,曹丰生
(江西省农业科学院 水稻研究所/水稻国家工程实验室(南昌)/国家水稻改良中心 南昌分中心,江西 南昌 330200)
叶脉是植物茎秆中维管束在叶片中的延伸和分支,能够发挥支撑叶片和输送水分、营养的作用。植物叶脉的形态结构较为简单,但是其形成过程涉及到非常复杂的分子机制。叶脉发育包括维管束的结构建成和排布延伸两个步骤,受到多个基因控制和内外因素共同影响[1]。单子叶植物和双子叶植物叶片中维管束的排布延伸方式不同,分别形成网状叶脉和平行叶脉。
水稻是单子叶植物中的模式植物,其中脉和侧脉沿着叶片纵向分布,其间通过细脉相连[2]。中脉位于水稻叶片的中央,最大的结构特征在于含有两个被称为透明细胞的小腔,是由大量细胞的细胞程序性死亡形成[3]。中脉在叶片远轴面和近轴面还分别具有一个中心维管束和小维管束,是支撑水稻叶片挺立的主要结构。中脉正常发育可以保证叶片维持直立的形态,增加叶片受光率,提高光合效能。因此,对水稻中叶脉相关性状的研究不仅有利于探明单子叶植物叶脉的发育机制,还能为构建水稻理想株型提供依据。
突变体是进行水稻分子生物学研究的重要材料,中叶脉缺失会引起水稻叶片披垂,是一个很明显的突变性状。Nagasawa等[4]在早期报道了4个叶片披垂的水稻突变体材料dl-1、dl-2、dl-sup1和dl-sup2,它们都不能形成中叶脉,因突变效应不同,颖花发育也出现了不同程度的变异。后续研究发现DL基因位于3号染色体的短臂上,上述4个突变体是DL基因发生了不同等位突变的结果。DL基因属于YABBY基因家族,编码的蛋白具有一个N端锌指结构域和一个C端YABBY结构域。水稻DL基因发生变异后,叶片中央区域的细胞不能沿着叶原基轴正常增殖,导致无法形成中叶脉[5]。此外,DL基因突变还会引起雌蕊发育异常,并且与SPW1基因之间存在负调控[6]。
国内也报道了多个水稻叶片披垂的无中叶脉突变体,普遍表现为隐性遗传,单个基因控制。利用遗传群体和分子标记分析,突变基因dl大多被定位在3号染色体的短臂上[7-11],也有个别被定位在1号染色体上[12]。另外,还有一个名为MR304的突变体的叶片披垂性状由两对隐性基因控制,其中一对为主效基因[13]。
水稻的中叶脉缺失会导致叶片披垂,影响叶片的光合性能,是一种不利突变性状。但是在水稻育种方面,可以将dl基因导入不育系中,培育叶片披垂的不育系。这种不育系的穗层明显高于叶层,在杂交制种过程中更容易授粉,从而获得较高的产量[14-16]。叶片披垂是隐形突变性状,与野生型外观区别明显,因此还可以作为形态学标记用于水稻纯度鉴定和杂株去除。
目前国内外报道的水稻中叶脉缺失突变的症状除叶片披垂外,大多还伴随花器官发育异常。本研究发现了一个中叶脉缺失的自然突变体,其花器官发育没有受到影响,与之前报道的突变表型不完全相同,命名为dl-7。本研究对该突变表型及遗传特性进行了分析,利用分子标记精细定位了突变基因,旨在为研究水稻叶脉的发育提供材料和依据。
1 材料与方法
1.1 试验材料
水稻材料来源于中种恢629与镇恢084杂交F2代中的一个自然变异单株,突变性状为叶片披垂。筛选至F10代后,获得了突变性状和农艺性状同时稳定的株系,命名为dl-7。将中种恢629作为野生型对照材料。
1.2 突变性状和农艺性状的观察
在水稻抽穗期观察突变体和野生型的植株形态,比较各叶位上叶片的形态和中脉发育。然后随机选择20个突变体单株,每个单株取50个颖花观察其形态结构。取剑叶的中间段,先用FAA固定液(70%乙醇90 mL,冰醋酸5 mL,37%甲醛5 mL)固定,然后用乙醇脱水。经石蜡包埋后用切片机制作5 μm厚的横切面切片,进行番红-固绿染色,最后用显微镜观察。待水稻成熟后,收获突变体和野生型各10个单株,考察结实率,用成组数据的t测验比较两者之间的差异。
1.3 遗传特性分析
以dl-7为母本,分别与本课题组的3个恢复系材料R071、R077和R112杂交,在F1代观察突变表型。将自交F2代群体各种植约300株,单本移栽,在水稻抽穗期观察突变表型,并统计叶片直立和叶片披垂的单株数量,检验遗传分离比。
1.4 基因定位
以dl-7与R077杂交的F2代作为遗传群体,对突变基因dl-7进行定位。取水稻叶片用CTAB法抽提小量基因组DNA;SSR标记引物序列根据网上(http://www.gramene.org/)已发布的序列进行设计,由江西鼎国创飞生物科技有限公司合成。用512个SSR标记对两个亲本进行分析,筛选具有多态性的标记。PCR反应体系和程序参照相关文献进行,取2 μL扩增产物用10%的非变性聚丙烯酰胺凝胶进行电泳,经银染显色后观察。
用筛选到的多态性SSR标记对30个F2代单株进行检测,将突变体带型记为1,野生型带型记为2,杂合体带型记为3,利用MAPMAKER(EXP3.0)作图软件对植株表型和SSR带型进行连锁分析[17]。然后在SSR标记定位的目标区域中开发序列缺失或插入位点(InDel),由Primer Premier 5.0设计引物。用多态性的InDel标记分析94个F2代突变单株,进一步精细定位dl-7。
2 结果与分析
2.1 突变性状及其影响
水稻株系dl-7在抽穗期的剑叶完全没有形成中脉,倒二叶、倒三叶和倒四叶的中脉以叶柄为起点,仅分别延伸到叶片1/4、1/2和1/2长度的位置。因此,dl-7各叶位上叶片表现出不同程度的披垂,导致整个植株叶片披散(图1A)。剑叶由于缺少中脉的支撑,呈现翻转180°~270°的状态(图1B)。20个突变体单株的所有颖花发育正常,形态结构完整,没有受到突变的影响(图1C)。考察了10个突变体单株的农艺性状,平均结实率为80.5%±1.7%,野生型对照为81.2%±1.9%,两者之间没有显著差异。

A.植株形态;B.剑叶形态;C.颖花结构。图1 dl-7的突变表型和颖花结构
从水稻叶片横切面可以看到,野生型对照的中脉显微结构中具有透明细胞(CC)、中央大维管束(CLV)和近轴端小维管束(ASV),以及叶肉细胞和薄壁细胞(图2A)。透明细胞是叶片中央区域的细胞大量增殖后,再经过细胞程序性死亡形成的空腔,成为中脉的主要结构特征。突变体叶片在中脉位置上形成的叶脉(突变体中脉)没有透明细胞,其形态和结构与侧脉类似(图2B)。将图2B中虚线框部分放大观察发现,突变体中脉除了没有透明细胞外,还缺少了近轴端小维管束,而中央大维管束正常存在(图2C)。突变体中脉的结构与侧脉类似,细微的差别体现在向近轴端凸起的高度不同,原因在于中脉中央大维管束上方的大型薄壁细胞(LPC)堆积了5~6层,而侧脉仅有2~3层(图2D)。
2.2 突变表型的遗传特性
dl-7与3个父本杂交获得的F1代植株都能形成正常的中叶脉,与野生型一致,表明突变性状受隐形基因控制。3个F2代群体都能分离出正常单株和突变单株,此外还出现了一种共显性单株。共显性单株的部分分蘖为突变型,其余分蘖为野生型;大部分共显性单株的突变型分蘖多于野生型分蘖。共显性单株的表型不同于F1代,因此在统计遗传分离比时将其归为突变单株。3个F2代群体中的正常单株和突变单株数比例符合3∶1,表明突变性状由1对隐形基因控制(表1)。
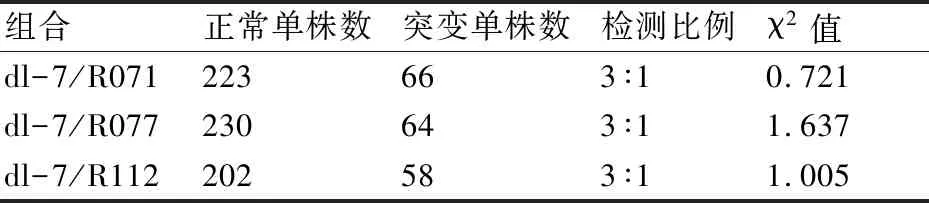
表1 突变表型在F2代群体中的分离
2.3 突变基因的精细定位
均匀分布于12条染色体的512个SSR标记中,在两个亲本之间表现出多态性的标记有110个,多态性频率为21.5%。用这110个多态性SSR标记对30个F2代单株进行检测,发现RM3392和RM1256两个标记与突变性状之间存在明显的连锁关系。RM3392和RM1256都位于水稻3号染色体的短臂上;根据交换单株的信息确定突变基因dl-7与这两个标记之间的遗传距离分别为6.7 cM和5.0 cM。
为了精细定位突变基因,在RM3392和RM1256之间的区段内开发了17个具有多态性的InDel标记。通过对94个F2代突变单株的检测,最终将dl-7定位在I3-14和I3-16之间280 kb的区域内,该区域涵盖了PAC克隆AC134241、AC135158和AC105732的区段(图3)。

A:野生型中脉(50×);B:突变体中脉(50×);C:突变体中脉(200×);D:突变体侧脉(200×)。 ASV:近轴端小维管束;CC:透明细胞;CLV:中央大维管束;LPC:大型薄壁细胞。图2 水稻叶片的横切面显微结构
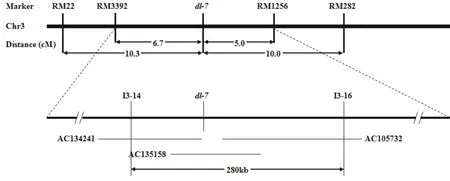
图3 突变基因dl-7的精细定位结果
3 讨论与结论
目前国内外报道了多个中叶脉缺失引起叶片披垂的水稻突变体,它们大多表现为中叶脉完全缺失,叶片严重披垂,并对颖花发育造成不同程度的影响。这些突变大多是由单个隐性基因控制的,突变基因被定位在3号或1号染色体上[7-12]。本研究中的水稻突变体与已报道的突变体不完全相同,具有自身的特点,在突变表型和遗传特性上都有体现,因此将其命名为dl-7。
dl-7仅有剑叶完全没有形成中脉,出现严重披垂和翻转的特征。倒二叶至倒四叶的中脉都有不同长度的延伸,仅表现出一定程度的披垂,这与罗琼等报道的整株叶片完全披垂的突变体dl(t)有所不同[7]。在叶片显微结构上,突变体dl-7的中央叶脉和侧脉结构类似,但是由于中央大维管束上方的大型薄壁细胞堆积层数有差异,使得两者向近轴端凸起的高度不同。而在Wang等报道的dl-6突变体中,中央叶脉和侧脉的结构是完全相似的[10]。这些表型特征说明dl-7的中叶脉发育过程并没有被完全阻断,只是在时间或空间上受到了抑制。
dl-7的颖花结构正常,结实率相比于野生型没有显著差异,表明突变没有对花器官的发育产生影响。dl-7最重要的特点在于F2代群体中分离出了一种突变型分蘖和野生型分蘖共存的共显性单株,这一现象在之前的突变体中未见报道。结合上述两个特征,我们认为dl-7是该类型中突变效应最弱的突变体。
突变基因dl-7是隐性基因,被定位在3号染色体短臂上的280 kb区段内,与多个文献报道的结果[7-11]一致。这个区段内包含一个YABBY基因,它的变异已经被证明能引起水稻中叶脉的缺失[5,10-11]。据此,我们推测这个YABBY基因是dl-7的候选基因,dl-7是这个YABBY基因的一个新的等位突变体。突变可能没有导致YABBY基因功能完全丧失,只是改变了表达量或表达调控模式,因此仅产生了较弱的突变效应。有关dl-7发生突变的具体位点和类型,以及对基因功能的影响,还有待后续进一步研究。
