新疆焉耆县不同动物源耐药沙门氏菌的MLST分析
2020-03-15王凯林亚军王舒丰李淑娴向志宇夏利宁
王凯, 林亚军, 王舒丰, 李淑娴, 向志宇, 夏利宁
(新疆农业大学动物医学学院, 乌鲁木齐 830052)
沙门氏菌(Salmonella)因其抗原性复杂、血清型多样以及临床表现和病理变化多,且能够同其他病毒与细菌产生并发性疾病,导致畜牧养殖业出现严重的经济损失[1]。由沙门氏菌造成的食物中毒和食源性疾病案例在国内外一直排名前列[2]。抗菌药物是目前治疗沙门氏菌病的有效手段,但随着其在临床的普遍使用,甚至滥用,造成沙门氏菌耐药菌株数量不断增加以及多重耐药菌株陆续出现,沙门氏菌耐药问题日渐严重,为畜牧养殖业的良好发展与人类的身心健康带来潜在威胁[3]。
新疆为少数民族居住地,养殖业发达,以牛、羊养殖为主,猪、禽养殖为辅。焉耆县位于新疆维吾尔自治区中部,共有23个民族寄居在此。随着多民族聚居,导致其对动物性食品种类的需求多样化,故焉耆县有着丰富的畜牧养殖资源。由于人们生活水平的提高,对肉类的食用量也呈上升趋势,随之由食品动物源耐药菌传播给人类的风险也逐渐增大,却鲜见对新疆不同动物源沙门氏菌耐药的相关研究。多位点序列分型(multi-locus sequence typing,MLST)是近年来发展起来的一种新型的分子分型方法,用于亲缘性关系分析。本研究通过对新疆焉耆县不同动物粪源分离的沙门氏菌进行药敏试验及相关耐药基因检测,分析耐药表型与携带的耐药基因之间的关系;并通过MLST方法分析不同动物源沙门氏菌之间的亲缘关系。本研究一方面可为焉耆县被检养殖场提供科学的临床用药方案,另一方面也为该县动物源沙门氏菌耐药库的建设提供数据,为今后开展动物源细菌耐药性风险评估提供理论依据。亲缘关系分析也为进一步研究沙门氏菌耐药传播机制提供基础。
1 材料与方法
1.1 试验材料
1.1.1样品 2016年7月于新疆焉耆县部分规模化养殖场使用灭菌肛拭子在动物直肠进行不同动物源新鲜粪样样品采集,共采集样品1 147份,其中牛源粪样97份、鸡源粪样400份、羊源粪样250份以及猪源粪样400份。
1.1.2标准质控菌株与培养基 Mueller-Hinton(MH)培养基、麦康凯琼脂培养基、氯化镁孔雀绿增菌液(MM)、沙门氏菌显色培养基、SS琼脂培养基均购自奥博星生物技术有限公司(北京);大肠埃希标准质控菌(ATCC25922)购自天和微生物试剂有限公司(杭州)。
1.1.3药敏试验所用药品 喹诺酮类:环丙沙星(ciprofloxacin,CIP)、诺氟沙星(norfloxacin,NOF)和恩诺沙星(enrofloxacin,ENR);β-内酰胺类:阿莫西林(amoxicillin,AML)、头孢噻呋(ceftiofur,CEF)和氨苄西林(ampicillin,AMP);氨基糖苷类:安普霉素(apramycin,APR)、阿米卡星(amikacin,AMK)、庆大霉素(gentamicin,GEN)和卡那霉素(kanamycin,KANA);四环素类:四环素(tetracycline,TE);酰胺醇类:氟苯尼考(florfenicol,FLR);多肽类:多黏菌素(polymyxin,CL)。上述药品的标准品均购自中国兽药监察所。
1.2 试验方法
1.2.1沙门氏菌的分离鉴定 将采集的肛拭子放入装有1 mL灭菌肉汤的2 mL EP管里,从EP里吸取50 μL样品至1 mL灭菌MM增菌液中,37 ℃培养12 h后用灭菌接种环在SS琼脂培养基上划线培养,37 ℃恒温培养18~24 h后,挑取中间黑色周围透明的单菌落至1 mL MM增菌液中,培养12 h后在沙门氏菌显色培养基上划线,37 ℃恒温培养18~24 h,培养基上有紫红色菌落长出,可初步鉴定为沙门氏菌,之后挑取单菌落保存备用。
对初步鉴定为沙门氏菌的菌株通过PCR检测沙门氏菌持家基因invA,根据是否检出invA基因来鉴定沙门氏菌[4]。引物由上海生工生物工程股份有限公司合成,序列见表1。PCR扩增反应体系25 μL,包括ddH2O 9.5 μL、2×TaqPCR Master Mix 12.5 μL、上下游引物各1 μL、1 μL模板。PCR反应程序见表2。PCR产物进行1.0%琼脂糖凝胶电泳检测。
1.2.2沙门氏菌的药敏试验及其判定 采用琼脂稀释法,对分离鉴定出的沙门氏菌进行上述6类13种抗菌药物最小抑菌浓度(minimal inhibitory concentrations,MICs)的测定,测定结果以3种形式记录:敏感、中介和耐药,结果判定参照CLSI标准[5]。
1.2.3耐药基因合成及序列分型 参考文献[6-16],委托生物工程股份有限公司(上海)分别合成β-内酰胺酶基因(blaTEM、blaCTX-M、blaSHV、blaLAP-1、blaKPC、blaOXA和blaCMY-2)、扩增PMQR因子(qnrA、qnrB、qnrC、qnrD、qnrS、oqxA、oqxB和aac(6′)-Ib)、四环素类基因(tetA和tetB)、氨基糖苷类基因[armA、rmtB、aadA2和ant(3″)-Ia]、酰胺醇类基因(floR)和多肽类基因(mcr-1)的引物(表1)。对检测出耐药的沙门氏菌分别进行上述基因的PCR检测,反应运行参数详见表2。PCR产物经1.0%琼脂糖凝胶电泳后使用凝胶成像系统进行拍照分析,将目的片段进行胶回收并送至生物工程股份有限公司(上海)测序,比对后确定基因型。进一步用BstF5Ⅰ酶进行酶切已检出aac(6′)-Ib基因的PCR产物,如果存在-cr突变情况,那么aac(6′)-Ib基因将失去BstF5 I酶的限制性位点并出现不能被切割现象。再次通过观察琼脂糖凝胶电泳图谱的条带可以得知aac(6′)-Ib基因中-cr有无突变现象。

表1 本研究所用PCR引物序列
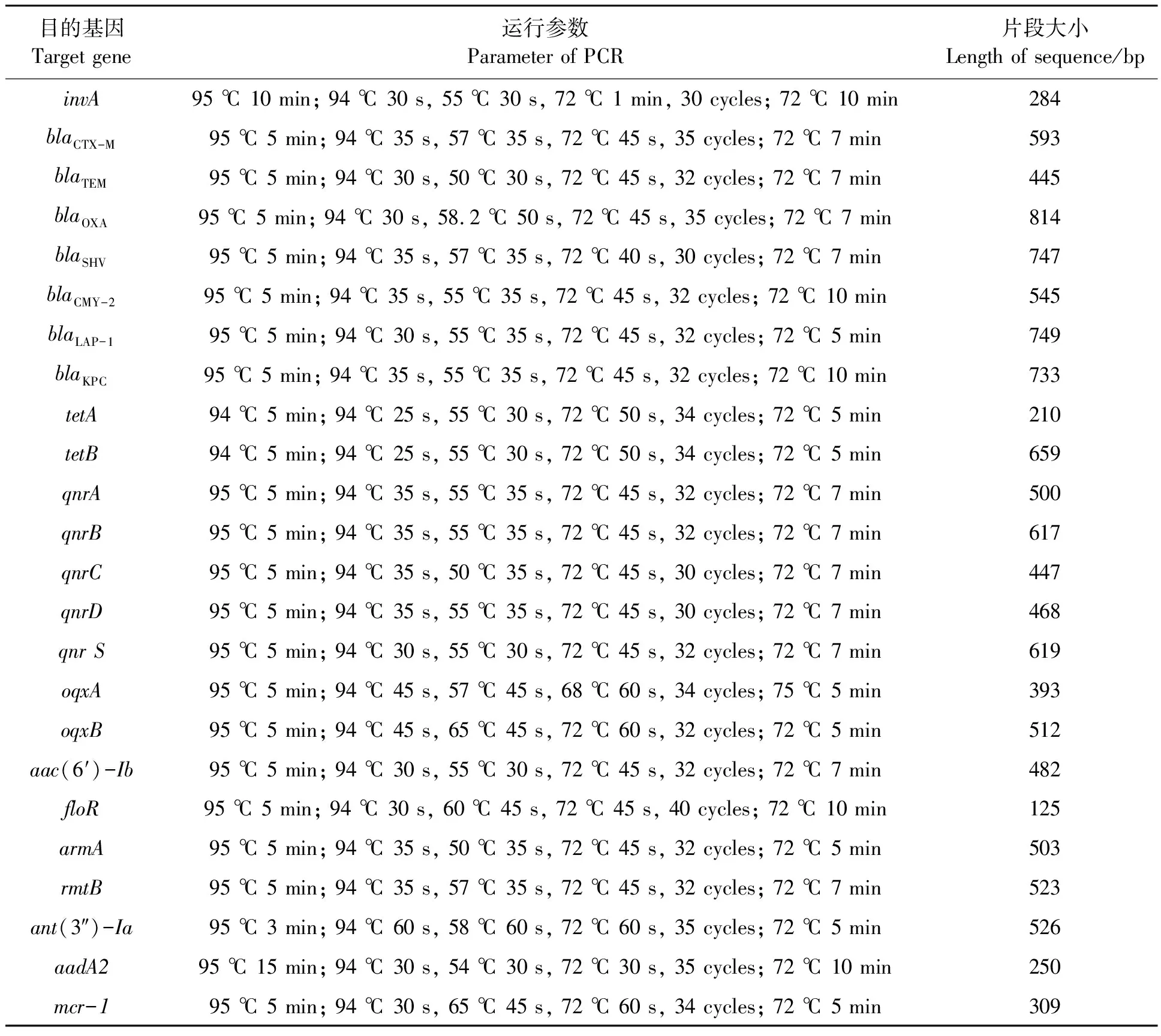
表2 PCR反应运行参数
1.2.4MLST分析 根据不同动物源样品的分离率,分别从焉耆县鸡源、牛源、羊源和猪源沙门氏菌中挑取多药耐药且携带多种耐药基因的8株菌,其中鸡源沙门氏菌3株,牛源沙门氏菌2株,羊源沙门氏菌1株和猪源沙门氏菌2株。
参照MLST网站(http://www.mlst.net)选择沙门氏菌的7个管家基因。根据Kidgell等[17]合成thrA、purE、sucA、hisD、aroC、hemD和dnaN共7对扩增引物,引物序列详见表1,扩增引物均委托生物工程股份有限公司(上海)合成。用MEGA6软件将测序结果进行剪切,将序列中用于分型部分序列提交至上述的MLST网站数据库中进行等位基因对比,得到相应的等位基因编号,之后把等位基因图谱输入Hps MLST数据库,按照指定排列顺序形成ST(sequence type)型[18-20]。等位基因的不同直接决定菌株的ST型不同,比较菌株的ST型即可得知菌株间的亲缘性关系,最后利用DNA star的MegAlign功能对不同菌株序列构建核苷酸序列比较图,利用Mega构建进化树。
1.2.5数据分析 耐药率结果与多药耐药结果采用SPSS 17.0软件进行卡方检验做差异显著分析。
2 结果与分析
2.1 不同动物源沙门氏菌分离鉴定
共分离沙门氏菌230株,分离率为20.1%(230/1 147)。其中牛粪源沙门氏菌分离率为64.9%(63/97),鸡粪源沙门氏菌分离率为27.5%(110/400),猪粪源沙门氏菌分离率为10.0%(40/400),羊粪源沙门氏菌分离率为6.8%(17/250)。
2.2 不同动物源沙门氏菌的耐药结果
由图1可知新疆焉耆县不同动物源沙门氏菌的整体耐药严重程度由高到低依次为牛源>猪源>羊源>鸡源。不同动物源沙门氏菌对头孢噻呋、阿米卡星和多黏菌素均高度敏感,无耐药株检出,但对其他被检抗菌药物均有不同程度耐药。牛源沙门氏菌对氟苯尼考(55.6%)、氨苄西林(50.8%)和卡那霉素(50.8%)耐药率均超过50%,对诺氟沙星(49.2%)、四环素(46.0%)、环丙沙星(44.4%)和阿莫西林/克拉维酸(44.4%)的耐药率超过40%,对安普霉素耐药率仅为1.6%;猪源沙门氏菌对四环素(55.0%)和氨苄西林(50.0%)两种抗菌药物耐药率超过50%,对阿莫西林/克拉维酸(40.0%)和氟苯尼考(40.0%)耐药率超过40%,对其他被检抗菌药物耐药率均未超过25%;鸡源沙门氏菌对氟苯尼考(19.1%)、氨苄西林(17.3%)、诺氟沙星(16.4%)、阿莫西林/克拉维酸(15.5%)、四环素(13.6%)、卡那霉素(12.7%)、环丙沙星(11.8%)和恩诺沙星(7.3%)的耐药率在7.3%~19.1%之间,对其他被检抗菌药物未检出耐药菌株。

注:不同字母表示不同动物源沙门氏菌对同种抗菌药物耐药性差异在P<0.05水平具有统计学意义。
牛源沙门氏菌对环丙沙星、诺氟沙星、恩诺沙星的耐药率显著高于其他三种动物源(P<0.05);对阿莫西林/克拉维酸与四环素耐药率显著高于鸡源与羊源沙门氏菌(P<0.05);猪源沙门氏菌对阿莫西林/克拉维酸耐药率显著高于鸡源和羊源耐药率(11.8%)(P<0.05),而对安普霉素耐药率显著高于牛源与羊源沙门氏菌(P<0.05);而鸡源与羊源沙门氏菌对试验中的抗菌药物的耐药率差异不显著(P>0.05)。
2.3 不同动物源沙门氏菌多药耐药结果
由图2可知,不同动物源沙门氏菌多药耐药结果以0耐为主,即对被检药物均不耐药。其中,牛源沙门氏菌多药耐药以0耐(33.3%)和7耐(对被检药物中的任意7种药物耐药)(33.3%)为主,除5耐和8耐外,均有分布;猪源沙门氏菌以0耐(32.5%)和6耐(17.5%)为主,在0耐~7耐均有分布;羊源沙门氏菌多药耐药以0耐(58.8%)和4耐(23.5%)为主,1耐、3耐和6耐也有分布;鸡源沙门氏菌以0耐(72.7%)为主,在0耐~8耐均有分布。表明焉耆县不同动物源沙门氏菌多药耐药情况较为严重,应在临床用药时替换已经产生耐药的抗菌药物,选择敏感药物进行治疗,以便达到较好的治疗效果。

注:不同字母表示不同动物源沙门氏菌的多药耐药差异在P<0.05水平具有统计学意义。
牛源沙门氏菌7耐菌株数显著高于其他3种动物源(P<0.05);且牛源、羊源、猪源沙门氏菌6耐菌株数均显著高于鸡源(P<0.05);羊源沙门氏菌4耐菌株数显著高于牛源和鸡源(0.0%)(P<0.05);而羊源与猪源沙门氏菌多药耐药结果差异不显著(P>0.05)。
2.4 不同动物源沙门氏菌耐药基因检测结果
由耐药基因检测结果(表3)可知,焉耆县不同动物源沙门氏菌均携带β-内酰胺酶基因、PMQR因子、氨基糖苷类基因、四环素类基因和酰胺醇类基因,其中β-内酰胺酶基因以blaTEM(牛源87.3%、羊源41.2%、鸡源19.1%和猪源12.5%)和blaOXA(牛源42.9%、羊源23.5%、鸡源19.1%和猪源15.0%)为主;PMQR因子以oqxA(牛源44.4%、羊源29.4%、鸡源19.1%和猪源15.0%)、oqxB(牛源42.9%、羊源29.4%、鸡源19.1%和猪源15.0%)和aac(6′)-Ib-cr(牛源44.4%、羊源23.5%、鸡源20.0%和猪源20.0%)为主;氨基糖苷类基因以ant(3″)-Ia(牛源44.4%、羊源29.4%、鸡源19.1%和猪源22.5%)和aadA2(牛源44.4%、羊源29.4%、鸡源19.1%和猪源10.0%)为主;四环素类基因以tetB(牛源46.0%、羊源41.2%、鸡源19.1%和猪源20.0%)为主;酰胺醇类基因以floR(牛源44.4%、羊源23.5%、鸡源16.4%和猪源32.5%)为主。此外,猪源沙门氏菌还检出blaCMY-2(15.0%)、qnrB(5.0%)、qnrS(12.5%)、tetA(15.0%)。而不同动物源分离的沙门氏菌均未检出基因blaSHV、blaKPC、blaLAP-1、blaCTX-M、armA、rmtB、qnrA、qnrC、qnrD和mcr-1。

表3 焉耆县不同动物源沙门氏菌耐药基因检测结果
牛源沙门氏菌的耐药基因携带率明显高于其他3种动物源沙门氏菌,牛源菌对blaTEM(87.3%)检出率超过85.0%,对blaOXA(42.9%)、aadA2(44.4%)、ant(3″)-Ia(44.4%)、oqxA(44.4%)、oqxB(42.9%)、aac(6′)-Ib(44.4%)、tetB(46.0%)和floR(44.4%)的检出率也超过40.0%。分析原因可能在牛沙门氏菌病临床用药过程中β-内酰胺类药物使用过度,导致牛源沙门氏菌β-内酰胺酶基因检出率过高,其次对喹诺酮类药物、四环素类药物以及酰胺醇类药物的使用不当造成PMQR因子、四环素类基因以及酰胺醇类耐药基因检出率较高。
2.5 不同动物源沙门氏菌多种耐药基因共存情况
对不同动物源分离的沙门氏菌进行多种耐药基因共存情况统计,结果(表4)可知,被检的110株鸡源沙门氏菌共有5种耐药基因共存类型,以blaTEM+blaOXA+oqxA+oqxB+aac(6′)-Ib-cr+aadA2+ant(3″)-Ia+tetB+floR为主,占15.5%(17/110);63株牛源沙门氏菌共有6种耐药基因共存类型,以blaTEM+blaOXA+oqxA+oqxB+aac(6′)-Ib-cr+aadA2+ant(3″)-Ia+tetB+floR为主,占33.3%(21/63);17株羊源沙门氏菌共有3种耐药基因共存类型,以blaTEM+blaOXA+oqxA+oqxB+aac(6′)-Ib-cr+aadA2+ant(3″)-Ia+tetB+floR为主,占23.5%(4/17);40株猪源沙门氏菌共有10种耐药基因共存类型,以blaOXA+oqxA+oqxB+aac(6′)-Ib-cr+aadA2+ant(3″)-Ia+tetB+floR(12.5%,5/40)为主。

表4 不同动物源沙门氏菌多种耐药基因共存情况
可见,除猪源沙门氏菌外,不同动物源沙门氏菌均有部分菌株同时携带5类耐药基因,并且鸡源(15.5%,17/110)、牛源(33.3%,21/63)和羊源(23.5%,4/17)沙门氏菌耐药基因共存类型均以blaTEM+blaOXA+oqxA+oqxB+aac(6′)-Ib-cr+aadA2+ant(3″)-Ia+tetB+floR为主,而猪源沙门氏菌以blaOXA+oqxA+oqxB+aac(6′)-Ib-cr+aadA2+ant(3″)-Ia+tetB+floR(12.5%,5/40)基因共存类型为主,分析原因可能是焉耆县规模化养殖场存在携带多药耐药基因的优势耐药菌株,其携带的耐药基因可以通过水平或垂直传播,进而使养殖场内出现大量携带相同耐药基因的耐药菌株。
2.6 不同动物源沙门氏菌MLST分析结果
从分离鉴定的230株不同动物源沙门氏菌挑选8株用于MLST试验,所挑选耐药株的来源、耐药表型及耐药基因携带耐药情况见表5。
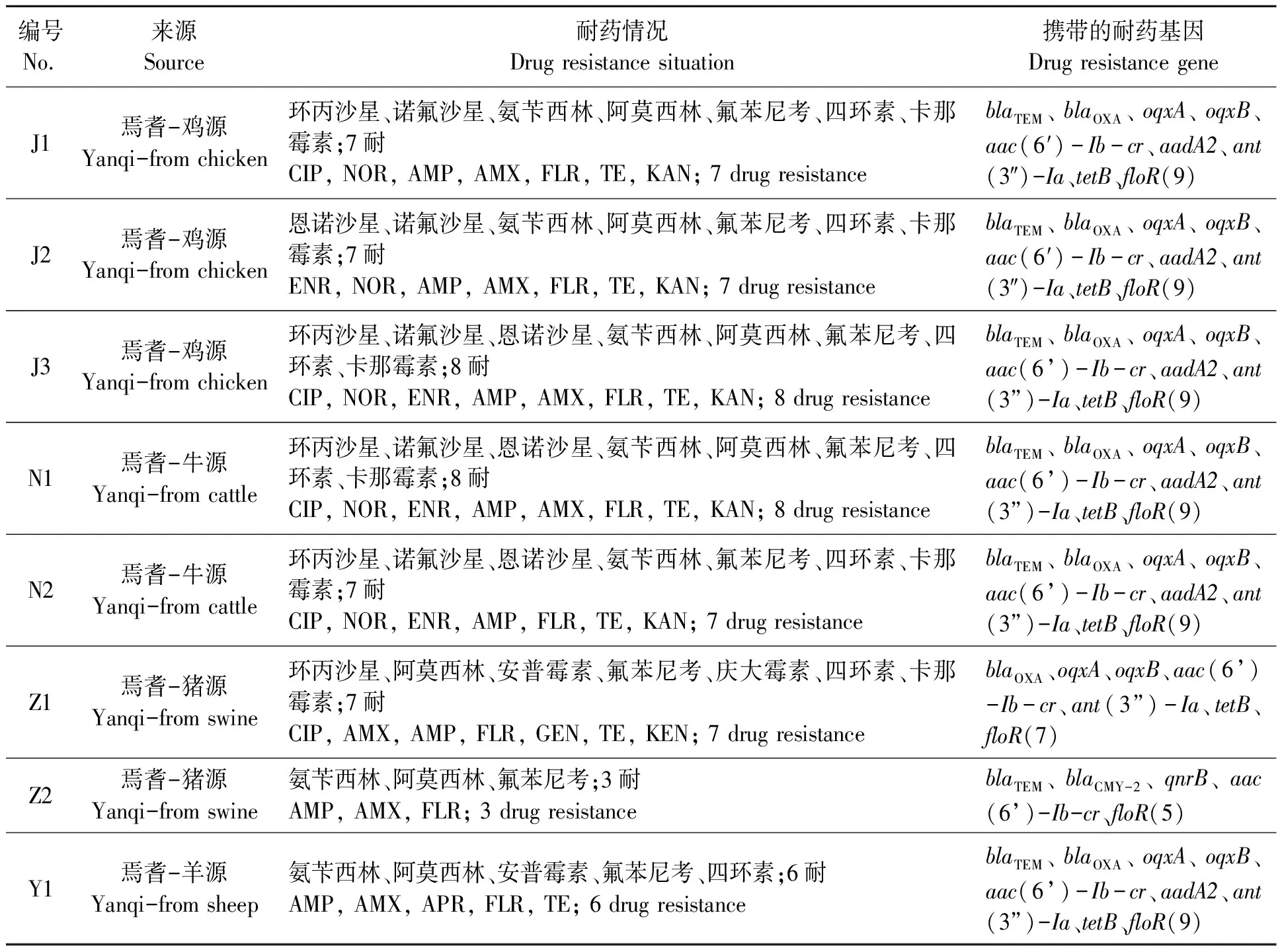
表5 MLST试验菌株背景
8株沙门氏菌MLST分型结果(表6)可知,不同动物源分离的沙门氏菌属于同一种ST型,即ST34,表明这8株沙门氏菌可能来源于同一克隆群,一方面说明焉耆地区沙门氏菌MLST分型以ST34为主,另一方面也说明焉耆地区沙门氏菌菌株具有在不同动物间和/或相同动物间进行传播的可能性,即在同一地区分离的不同动物源耐药沙门氏菌可能存在种间和/或种内克隆传播的耐药传播机制。
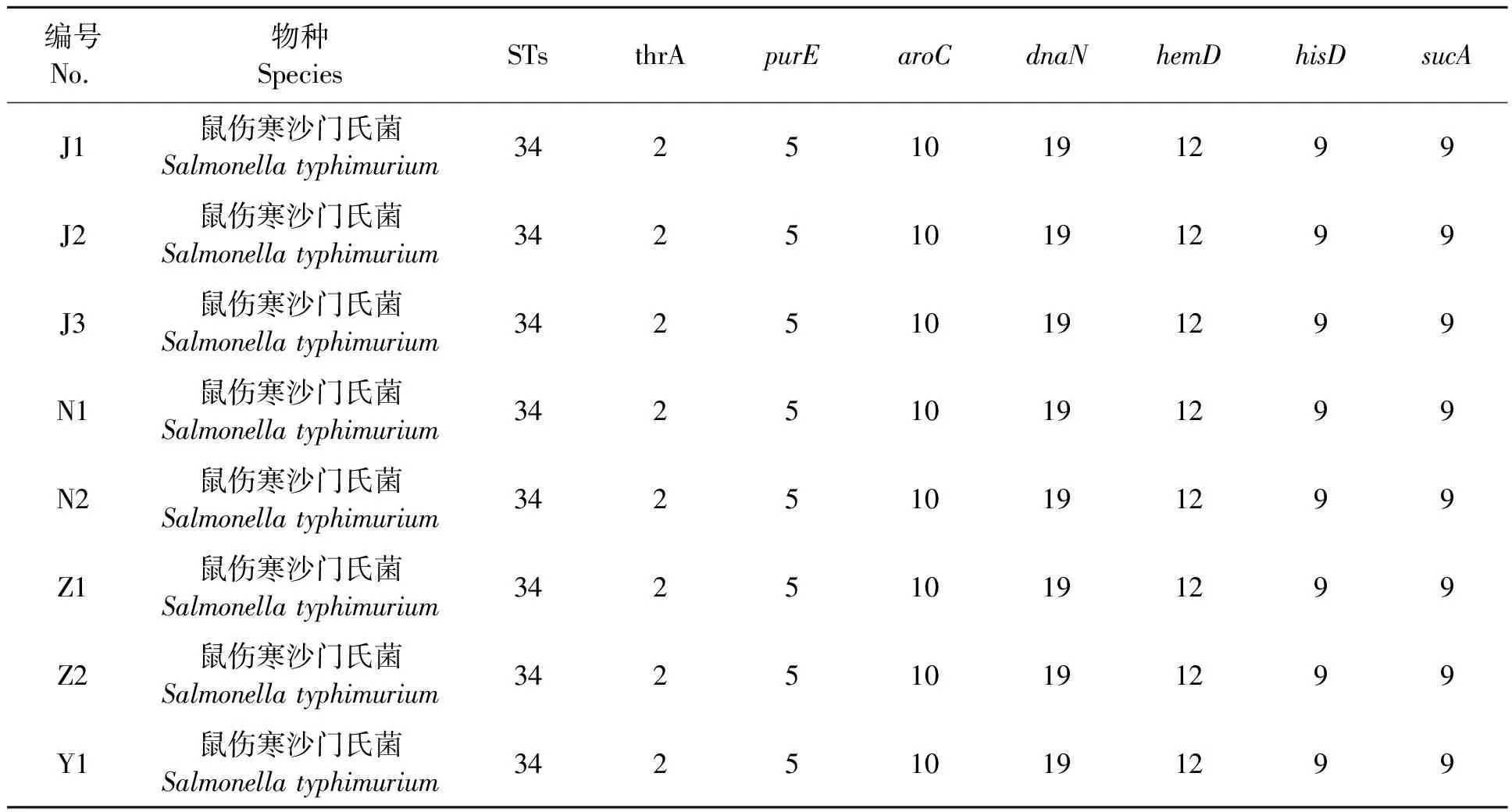
表6 焉耆县不同动物源沙门氏菌等位基因情况
2.7 8株沙门氏菌进化树结果
对8株不同动物源分离的沙门氏菌进行进化树比较,以此来分析不同动物源沙门氏菌的亲缘关系,不同动物源分离的沙门氏菌同源性高达99.7%及以上。进化树如图3所示,可以看出,J2与J3亲缘性关系较近;N1与Z1亲缘关系较近,而Y1与其他7株沙门氏菌亲缘关系相对较远;8株不同动物源分离的沙门氏菌亲缘关系较近。
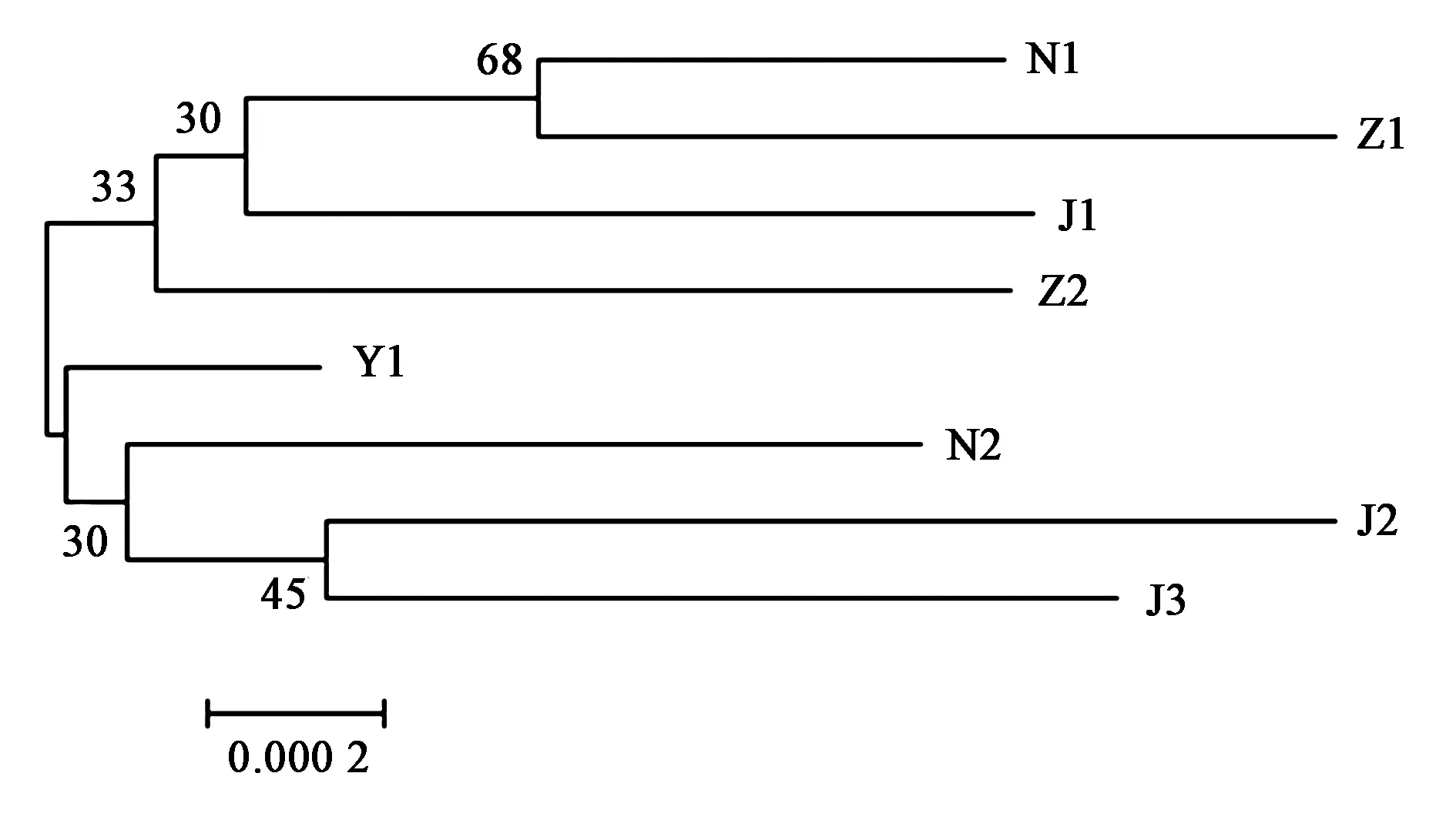
图3 8株沙门氏菌核苷酸序列进化树分析
3 讨论
牛源沙门氏菌对氟苯尼考的耐药率最高(55.6%),略高于赵俊利等[21]对氟苯尼考耐药率(42.1%)的报道。分析原因可能是牛为焉耆县常见大型家畜,而夏季是沙门氏菌的高发期。因此,本研究出现低采样率而沙门氏菌分离率高的现象,而在应对牛沙门氏菌病时,养殖人员可能运用过多种抗菌药物进行治疗,从而导致牛源沙门氏菌耐药情况最严重。牛源沙门氏菌对氨苄西林和卡那霉素的耐药率均为50.8%,远低于赵俊利等[21]的报道,可能是因为用药习惯的不同所致。猪源沙门氏菌对四环素的耐药率为55.0%,低于支威等[22]的报道(75.0%);对氨苄西林的耐药率(50.0%)低于曹正花等[23]的报道(80.7%),但高于王伟等[24]和杜雄伟等[25]的报道(20.0%和6.7%);可能是因为不同地区与不同饲养环境造成。猪源沙门氏菌对本研究抗菌药物耐药率高于羊源和鸡源沙门氏菌,可能是因为规模化养殖下,为降低猪只的发病率,会在饲养过程中使用带有抗菌药物的饲料对猪只饲喂;此外,治疗细菌病时用药多样,导致猪源沙门氏菌耐药率较高且耐药谱型多样化。采集的250份羊粪源样品中仅分离出17株沙门氏菌,对氟苯尼考和四环素耐药率均为35.3%,远低于江萍等[26]对氟苯尼考耐药率(92.3%)和四环素耐药率(90.4%)的报道,可能与所采集的羊本身健康状况及养殖场的用药差异有关。由于鲜见对羊源沙门氏菌耐药性相关报道,研究结果可为羊源沙门氏菌的研究提供数据参考。而鸡源沙门氏菌耐药情况并不严重,对氟苯尼考的耐药率最高,仅为19.1%,低于彭斌等[27]的报道(25.0%);对氨苄西林的耐药率(17.3%)远低于其他研究[27-30]报道。鸡源沙门氏菌对被检药物的耐药率低的原因,还需要结合用药调研及与饲养场养殖模式进一步分析。
本研究中,4种动物源均检测出β-内酰胺酶耐药基因、PMQR因子、氨基糖苷类耐药基因、四环素类耐药基因和酰胺醇类耐药基因,β-内酰胺酶耐药基因以blaTEM和blaOXA基因为主;PMQR因子以oqxA、oqxB和aac(6′)-Ib-cr基因为主;氨基糖苷类耐药基因以ant(3″)-Ia和aadA2基因为主;四环素类耐药基因以tetB基因为主;酰胺醇类耐药基因以floR基因为主。此结果与申永秀等[31]报道的结果(以携带基因blaTEM、tetA为主)不同,可能是由于地域不同,临床用药差异导致。PMQR因子在4种动物源中的检出率虽然不高(5.0%~44.4%),但是PMQR基因的流行可以促进编码DNA促旋酶的gyrA和gyrB基因突变,及编码拓扑异构酶Ⅳ的parC和parE基因突变,从而引起高水平的耐药性[32-33]。本研究中不同动物源沙门氏菌的耐药表型及对应的耐药基因检出率不高,远低于罗永乾[34]的报道,可能是由于焉耆地区养殖密度远低于其他地区,临床用药较少,所以整体耐药情况不严重。造成不同动物源耐药情况不同,耐药基因检出率有差异可能是由于养殖场技术人员在进行沙门氏菌病的治疗时,不同养殖场用药背景和用药习惯不同造成。
本研究发现焉耆地区沙门氏菌MLST分型以ST34为主,与陈健皓[35]和刘慧玲等[36]的研究结果不同,与赵翠等[37]报道的潍坊与烟台猪源沙门氏菌的MLST分型结果一致。表明虽然不同地区用药情况不同,但却有可能获得相同的MLST分型,具体耐药传播机制及MLST分型形成原因有待进一步研究。
