峨眉山世界遗产地表土孢粉组合及其生态和古环境启示
2020-03-13李永飞夏中林沈华东陈发军凌超豪
李永飞, 夏中林, 沈华东, 许 斌, 陈发军, 凌超豪
1 吉首大学旅游与管理工程学院 生态旅游湖南省重点实验室, 张家界 427000 2 内江师范学院 地理与资源科学学院, 内江 641112 3 峨眉山林业管理所, 峨眉山 614201 4 河海大学地球与工程学院 水文水资源与水利工程科学国家重点实验室, 南京 210098 5 内江师范学院 生命科学学院, 内江 641112 6 中国科学院南京地理与湖泊研究所 湖泊与环境国家重点实验室, 南京 210008
地层中化石孢粉的分析是探讨第四纪古植被,重建古气候和古地理的重要方法之一,受到古生态学家和古环境学家广泛关注[1- 4]。现代花粉与植被之间关系的正确解译是探讨地层花粉组合和重建地质时期植被的依据,但因受孢粉产量、传播、来源范围、沉积、保存以及自然地理环境等因素影响,致使其与植被的关系变得十分复杂[5- 8]。表土花粉组合与母体植被关系的研究长期以来是第四纪孢粉学中的重要内容,迄今为止仍然受到学者关注[9- 12]。近年以来我国孢粉学者对东北[13- 16]、华北[17- 19]、西北[20- 24]、青藏高原[25- 29]及华东[30]、华中[31-32]和华南[33-35]等地区表土花粉研究仍方兴未艾,成果卓著,西南地区现代花粉分析[36-38]也为该区古植被和古生态恢复提供了重要依据[39-41]。
始新世末至新近纪伴随着青藏高原及喜马拉雅山-横断山的阶段隆升和古地中海的退却[42-43],包括峨眉山在内的康滇古陆植物区系与东部扬子古陆和南部的冈瓦纳古陆植物区系产生交流,峨眉山以其独特的地理、地貌和小气候等因素成为许多北方古近纪植物避难所和新植物类群演化摇篮,致使热带、亚热带和温带植物成分在此交汇、融合[44-46]。现代峨眉山植物区系成分更加复杂和多样化,有高等植物280科1271属3703多种[47],既包含中国-日本植物区系成分,又有中国-喜马拉雅植物区系成分[46,48-49],特有植物种类约106种[47],全山森林覆盖率约98.8%[50]。在如此狭窄的地理区域范围内既广泛分布热带亚热带常绿阔叶代表类群樟科群落,也大量生长第三纪孑遗落叶阔叶珙桐群落,还有四川特有种峨眉冷杉群落,孕育着这样丰富的植物种质资源、完整的植被类型和森林垂直带谱,是全球亚热带山地保存最完好的原始植被景观之一,这在中国乃至世界都是罕见的,具有十分重要的科研价值、观赏价值和经济价值[48, 51]。本研究可以为植物种类丰富的热带亚热带山地和相似地区古植被和古气候重建,环境变迁和植被生态恢复提供基础资料和参考依据。
1 研究区概况
峨眉山位于四川省乐山市峨眉山市,介于29°16′30″—29°43′42″N,103°10′30″—103°37′10″E之间,地处四川盆地西南邛崃山南段余脉向青藏高原过渡地带,系第三纪中新世喜马拉雅造山运动形成剥蚀背斜褶皱断块山,最高峰万佛顶3099 m,与山麓500 m相对高差近2600 m;属中亚热带季风气候,山麓年均气温17.2℃,1月平均气温6.9℃,7月平均气温26.1℃,年降水量1555.3 mm,年日照时数948.5 h;山顶年平均气温3.0℃,1月平均气温-6.0℃,7月平均气温11.8℃,年降水量1922.8 mm,年日照时数1398.1 h;全山相对湿度80%以上;由山麓至山顶主要有紫色土、黄壤、山地黄壤、山地黄棕壤、山地暗棕壤、山地灰化土和山地草甸土等;受地质、地貌、气候、土壤和人类活动等影响,植被垂直地带性分异显著,从山麓至山顶依次分布低山常绿阔叶林间人工次生林-中山常绿阔叶林-常绿落叶阔叶混交林-针阔混交林-针叶林-灌丛草甸[45,47,52],如图1。

图1 研究区和采样点示意图Fig.1 Schematic diagram of study area and sampling sites图中采样点E指峨眉山
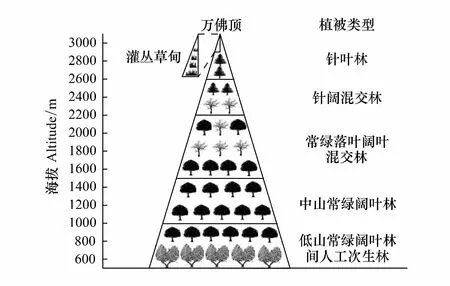
图2 峨眉山植被垂直带谱(修改自[51]) Fig.2 Vertical vegetation zones in the Emei Mountains (modified from references [51])
(1) 低山常绿阔叶林间人工次生林
本区地带性植被类型分布于海拔500—1000 m,为亚热带低山常绿阔叶林,乔木层有细叶楠(Phoebehui)、润楠(Machiluslongipedicellata)、枫杨(Pterocaryastenoptera)、柏木(CupressusfunebrisEndl.)、香樟(Cinnamomumcamphora)、头状四照花(Cornuscapitata)、木姜子(Litseapungens)、灯台树(Bothrocaryumcontroversum)等。林下灌木盖度0.3—0.6,有姜花(Hedychiumcoronarium)、水麻(Boehmeriapenduliflora)、构树(Broussonetiapapyrifera)、女贞(Ligustrumlucidum)、竹(Bambusoideae)、柃木(Euryajaponica)、铁线莲(Clematisflorida)、崖爬藤(Tetrastigmaformosanum)和棕(Trachycarpusfortunei)等。地被植物覆盖度0.8,有冷水花(Pileanotata)、扁竹兰(Irisconfusa)、报春花(Primulamalacoides)和蕨类等。地带性植被遭人工砍伐破坏后,少数地区零星分布,次生植被主要为黄葛树(Ficusvirensvar.sublanceolata(Miq.) Corner)、板栗(Castaneamollissima)、杉木(Cunninghamialanceolata)、柳杉(Cryptomeriafortunei)和水冬瓜(Alnuscremastogyne)等。
(2) 中山常绿阔叶林
本带植被类型为亚热带中山常绿阔叶林,分布于海拔1000—1500 m,乔木层主要以壳斗科石栎属(Lithocarpus)、青冈属(Fagus)、栲属(Castanopsis),樟科桢楠属(Phoebe)、润楠属(Machilus)、黄肉楠属(Actinolaphne)、樟属(Cinnamomum)、山胡椒属(Lindera)和木姜子属(Litsea),山茶科木荷属(Schima)等为优势种,主要树种有罗汉松(Podocarpusmacrophyllus)、黄心夜合(Micheliamartinii)、楠木(Phoebezhennan)、柏木、领春木(Eupteleapleiosperma)、红茴香(Illiciumhenryi)、峨眉含笑(Micheliawilsonii)、杉木、曼青冈(Cyclobalanopsisoxyodon)、桃叶珊瑚(Aucubachinensis)、交让木(Daphniphyllummacropodum)、栲树(Castanopsisfargesii)、香樟等。林下灌木丛生,附生植物、藤本植物在林中交织,森林里郁闭度较大,灌木层以山茶科(Theaceae)、五加科(Araliaceae)、杜鹃花科(Ericaceae)、箭竹(FargesiaspathaceaFranch)、方竹(Chimonobambusaquadrangularis)为主;草本植物近50种,盖度20%—50%。
(3) 常绿落叶阔叶混交林
本带分布于1500—2200 m,乔木层种类比较丰富,常绿阔叶林以润楠、峨眉栲(Castanopsisplatyacantha)、包石栎(Lithocarpuscleistocarpua)、交让木等为优势种,并生长着硬壳柯(Lithocarpushancei)、青冈(Cyclobalanopsisglauca)、丝栎(Lithocarpussp.)、黑壳楠(Linderamegaphylla)、柃木(Euryanitida)、西南山茶(Camelliapitardii)、四川山矾(Symplocossetchuensis)、杨叶木姜子(Litseapopulifolia)、西南华木荷(Schimawallichii)、刺榛(Corylusferox)等;落叶树种以白辛树(Pterostyraxpsilophylla)、木瓜红(Rehderodendronmacrocarpum)、鸡爪槭(Acerpalmatum)、稠李(Padusobtusata)、珙桐(Davidiainvolucrata)和华西枫杨(Pterocaryainsignis)占优势,其他可见灯台树(Cornscontroversa)、黄杞(Engelhardtiaspicata)、泡花树(Meliosmacuneifolia)、四照花(Dendrobenthamiacapitata)、水青树(Tetracentronsinense)、领春木(Eupteleapleiosperma)、扇叶槭(A.flabellatum)、糙皮桦(Betulautilis)、野漆(Toxicodendronsuccedaneum)、角翅卫矛(Euonymuscornutus)、山茱萸(Macrocarpiumchinense)、猫儿屎(Decaisneafargesii)、五尖槭(A.maximowiczii)、西南野樱(Prunuspilosiuscula)、四川蜡瓣花(Corylopsiswillmottiae)、安息香(Styraxsp.)等组成,郁闭度约为0.7。灌木层主要有冷箭竹(Bashaniafangiana)、溲疏(Deutziapilosa)、芒刺杜鹃(Rhododendronstrigillosum)、红豆杉(Taxuschinensis)、灯笼花(Fuchsiahybrid)等,覆盖度为0.2—0.4。草本层常见冷水花(Pileanotate)、管花鹿药(Smilacinahenryi)、五匹青(Pternopetalumvulgare)、锈毛金腰(Chrysospleniumdavidianum)、齿裂千里光(Seneciowinklerianus)、红花山酢浆(Oxaliscorymbosa)、吉祥草(Reineckiacarnea)、菝葜(Smilaxchina)等,覆盖度0.1—0.3。层间藤本常见美味猕猴桃(Actinidiachinensisvar.deliciosa)、金刚藤(Smilaxscobinicaulis)、常春藤(Hederanapalensisvar.sinensis)等。
(4) 针阔混交林
本带分布于2200—2600 m,乔木层针叶树以冷杉(Abiesfabri)为主,阔叶树主要有扇叶槭(AcerflabellatumRehd.)、五尖槭(A.maximowiczii)、细齿稠李(Padusobtusata)和糙皮桦(Betulautilis)等,并可见铁杉(Tsugachinensis)、华西枫杨(Pterocaryainsignis)、金顶柳(Salixhsinhsuaniana)、全苞石栎(Lithocarpuscleistocarpus)、海绵杜鹃(Rhododendronpingianum)等。灌木层以峨眉蔷薇(R.omeiensisRolfe)、美丽杜鹃(R.calophytum)、方竹(Chimonobambusaquadrangularis)为主,覆盖度0.3—0.4。草本层以山醡浆草(Oxalisgriffithii)、蟹甲草(Sacaliasp.)、虎耳草(Saxifragasp.)为主。

图3 峨眉山各植被类型景观Fig.3 Landscape of various plant communities in the Emei Mountains
(5)针叶林
本带分布于2600—3099 m,乔木层分布冷杉和铁杉,郁闭度0.6—0.9。灌木层以冷箭竹(Bashaniafangiana)和大箭竹(Sinarundinariachungii)为主,还包括高山绣线菊(Spiraeaalpina)、美容杜鹃(Rhododendroncalophytum)、大白杜鹃(Rh.decorum)、高山柳(Salixcupularis)、冰川茶藨子(Ribesglaciala)、长串茶藨(R.longiracemosum)和峨眉蔷薇(Rosaomeiensis),覆盖度0.6—0.8;草本层主要有山醡浆草(Oxalisacetosella)、石生楼梯草(Elatostemarupestre)、双舌蟹甲草(Cacaliadavidii)、条叶囊瓣芹(Pternopetalumtanakae)、疏穗苔草(Carexremotiuscula)、松下蓼(Polygonumpinetorum)和早熟禾(Poaannua)等,覆盖度0.2左右。
(6)灌丛草甸
本带分布于海拔2900 m以上的亚高山地带,应为冷杉林遭受破坏后发育起来的,主要有箭竹、杜鹃(Rh.simsiiPlanch.)、高山柏(Sabinasquamata)、金顶柳(S.luctuosa)、西南樱桃(Prunusduclouxii)、峨眉蔷薇、陕甘花楸(SorbuskoehneanaSchneid.)及冷杉幼苗等。地被植物有早熟禾和冰川茶藨子等。
2 研究方法
样品采集。表土样品采集和植被样方调查同时进行,样方面积森林为10 m×10 m,灌木为5 m×5 m,采用全球定位系统(GPS)定位样方的经度、纬度和海拔,自山麓至山顶顺坡而上海拔每升高大约100 m依据梅花5点法采集地表苔藓样品约200 g,各样点主要植物类型如表1。
实验分析。样品采用常规的氢氟酸HF法处理[53],取样约5—10 g,加入1片石松孢子 (27637粒)计算孢粉浓度,去除样品中碳酸盐、硅酸盐、腐殖质和有机质等,然后在超声波振荡器中用10 μm筛网富集孢粉,最后加入甘油保存,待制片鉴定。
孢粉鉴定统计参照《中国热带亚热带被子植物花粉形态》[54]、《中国植物花粉形态》[55]和《中国第四纪孢粉图鉴》[56]等植物花粉形态书刊以及现代花粉标准片在德国Zeiss生物显微镜下完成。
每个样品统计孢粉478—1709粒,平均756粒。
数据处理。将孢粉统计结果以占陆生种子植物花粉总和为基数进行百分比计算,并运用Tilia 软件进行孢粉图式制图[57]。Canoco for Windows软件是生态学用于约束与非约束排序工具,能获得生态数据统计模型,可以洞察生物群落结构、植物与植物群落以及它们之间的环境联系[58],文中利用Canoco软件对峨眉山表土孢粉数据进行统计和排序。

表1 峨眉山40个表土样点位置和植被类型
E :东,即East缩写;NE :北东,即Northeast缩写;W :东,即West缩写;NW :北西,即Northwest缩写;SW :南西,即Southwest缩写;SE :南东,即Southeast缩写;S :南,即South缩写;NNE :北北东,即North-northeast缩写;N :北,即North缩写;SSW :南南西,即South-southeast缩写;ENE :东北东,即East-northeast缩写;NNW :北北西,即North-northeast缩写;WNW :西北西,即West-northeast缩写
3 结果与分析
峨眉山40个表土孢粉样品共鉴定出136科(属),其中乔木46科(属),灌木35科(属),草本38科(属)和蕨类孢子16科(属)等。主要孢粉类型有:针叶树花粉有冷杉属、柏科、杉科和松属等;常绿阔叶树花粉有栲属/柯属、常绿栎类、青冈属和桃金娘科等;落叶阔叶树花粉可见栗属、落叶栎类、桦属、桤木属、胡桃属、枫杨属、榆属、青檀属、枫香属、金缕梅科、槭属、桑属和盐肤木属等;灌木植物花粉有鹅耳枥属/榛属、杜鹃花科、接骨木属、蜡瓣花属、绣线菊属、蔷薇科、野桐属和木樨科等;陆生草本植物花粉有禾本科、蒿属、藜科、十字花科、蝶形花科、毛茛科、葎草属、荨麻属、虎耳草属、凤仙花属和莎草科等;水生植物花粉可见香蒲属、鸭跖草科、泽泻属、慈姑属等;蕨类孢子主要有水龙骨科、里白属、卷柏属、凤尾蕨属、紫萁属、桫椤属等(图4,7,8,9)。

图4 峨眉山表土孢粉百分图谱Fig.4 Pollen percentage diagram of the surface samples from the Emei MountainsCupressaceae:柏科; Taxodiaceae:杉科; Abies:冷杉属; Pinus:松属;Castanopsis/Lithocarpus:栲属/柯属; Evergreen quercus:常绿栎类; Cyclobalanopsis:青冈属; Myrtaceae:桃金娘科; Castanea:栗属; Deciduous quercus:落叶栎类; Betula:桦属; Alnus:桤木属; Juglans:胡桃属; Pterocarya:枫杨属; Hamamelidaceae:金缕梅科;Morus:桑属; Cornaceae:山茱萸科; Rhus:盐肤木属; Carpinus/Corylus:鹅耳枥属/榛属;Spiraea:绣线菊属; Rosaceae:蔷薇科; Mollotus:野桐属; Oleaceae:木樨科;Gramineae:禾本科; Artemisia:蒿属; Chenopodiaceae:藜科; Fabaceae:蝶形花科;Ranunculaceae:毛茛科; Urtica:荨麻属; Saxifraga:虎耳草属; Impatiens:凤仙花属;Cyperaceae:莎草科; Conifer:针叶树; Evergreen broad-leaved trees:常绿阔叶树;Deciduous broad-leaved trees:落叶阔叶树; Shrubs:灌木; Terrestrial herbs:陆生草本;Ferns:蕨类; Spores & pollen sum:孢粉总数; Pollen concentration:孢粉浓度
3.1 花粉组合与浓度整体特征
峨眉山所有样品花粉组合中木本植物百分含量(44.5%—96.0%)占绝对优势,平均83.3%,其中针叶树、常绿阔叶树、落叶阔叶树和灌木花粉含量分别为24.1%、5.1%、36.2%和17.9%;草本植物花粉含量16.7%。木本植物中落叶阔叶树花粉含量(12.2%—77.6%)最高,以桤木属(2.7%—33.4%)和桦属(0.4%—33.4%)为主,枫杨属(0.1%—65.1%)和盐肤木属(0—68.9%)有一定含量;针叶树花粉含量(6.6%—62.5%)次之,主要有松属(1.6%—34.3%)和杉科(0.5%—53.6%),柏科(0—9.6%)和冷杉属(0—12.9%)有一定含量;灌木植物花粉含量(1.5%—54.9%)较低,蔷薇科占优势(0—50.2%),鹅耳枥属/榛属(0—32.3%)、绣线菊属(0—14.5%)和木樨科(0—10.4%)有一定含量;常绿阔叶树花粉含量(0.4%—59.9%)最低,以栲属/柯属(0—57.7%)和常绿栎类(0—6.2%)为主。草本植物花粉(4.0%—55.5%)中禾本科(0—21.3%)、蒿属(0—7.4%)和毛茛科(0—27.4%)含量较高,十字花科(0—4.9%)、虎耳草属(0—15.3%)、凤仙花属(0—14.3%)和莎草科(0—14.8%)有一定含量。木本植物花粉(Arboreal Pollen)与草本植物花粉(Nonarboreal Pollen)比值(AP/NAP)约为5。
峨眉山表土孢粉浓度(17513—767450粒/g)很高,平均174003粒/g(图5),桤木属浓度最高,达到24349粒/g;其次杉科(10176粒/g)、松属(16831粒/g)和蔷薇科(17964粒/g)等浓度在10000粒/g以上,桦属(9399粒/g)、枫杨属(6389粒/g)、蒿属(5639粒/g)、毛茛科(7628粒/g)和水龙骨科(9383粒/g)等浓度均超过5000粒/g;此外,柏科(2628粒/g)、冷杉属(1823粒/g)、栲属/柯属(3872粒/g)、常绿栎类(2645粒/g)、落叶栎类(2743粒/g)、桑属(2952粒/g)、盐肤木属(2419粒/g)、绣线菊属(2078粒/g)、禾本科(3671粒/g)、虎耳草属(2589粒/g)、和里白属(2704粒/g)等浓度均较高,高于2000粒/g。桤木属、蔷薇科、松属和杉科花粉浓度远远高于其他孢粉类型,这可能反映其母体植物在峨眉山花粉产量总量较大,或易散播的特征。

图5 峨眉山表土孢粉浓度图谱Fig.5 Pollen concentration diagram of the surface samples from the Emei MountainsCupressaceae:柏科; Taxodiaceae:杉科; Abies:冷杉属; Pinus:松属; Tsuga:铁杉属Castanopsis/Lithocarpus:栲属/柯属; Evergreen quercus:常绿栎类; Cyclobalanopsis:青冈属Myrtaceae:桃金娘科; Castanea:栗属; Deciduous quercus:落叶栎类; Betula:桦属; Alnus:桤木属; Juglans:胡桃属; Pterocarya:枫杨属; Ulmus:榆属; Pteroceltis:青檀属;Liquidambar:枫香属; Hamamelidaceae:金缕梅科; Mangnoliaceae:木兰科; Acer:槭属;Morus:桑属; Cornaceae:山茱萸科; Rhus:盐肤木属; Carpinus/Corylus:鹅耳枥属/榛属;Ericaceae:杜鹃花科;Sambucus:接骨木属; Corylopsis:蜡瓣花属; Spiraea:绣线菊属; Rosaceae:蔷薇科; Mollotus:野桐属; Oleaceae:木樨科;Gramineae:禾本科; Artemisia:蒿属; Chenopodiaceae:藜科; Fabaceae:蝶形花科;Ranunculaceae:毛茛科; Humulus:葎草属; Urtica:荨麻属; Saxifraga:虎耳草属; Impatiens:凤仙花属;Cyperaceae:莎草科;Pollen concentration:孢粉浓度
3.2 DCA
利用Canoco 5.0 软件对峨眉山表土花粉百分含量进行降趋势对应分析(DCA) (图6),从分析的136科(属)中除去含量小于1%和只在个别样品中出现的科(属),并对剩余的52科(属)中含量较小的科(属)降低权重进行DCA。
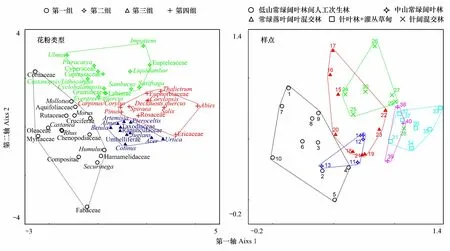
图6 花粉类型与样点的DCA排序图Fig.6 DCA results of surface pollen taxa and pollen sampling sitesCupressaceae:柏科; Taxodiaceae:杉科; Abies:冷杉属;Pinus:松属; Castanopsis/Lithocarpus:栲属/柯属; Evergreen quercus:常绿栎类; Cyclobalanopsis:青冈属;Myrtaceae:桃金娘科; Castanea:栗属; Deciduous quercus:落叶栎类; Betula:桦属; Alnus:桤木属; Juglans:胡桃属; Pterocarya:枫杨属; Ulmus:榆属; Pteroceltis:青檀属;Liquidambar:枫香属;Hamamelidaceae:金缕梅科;Eupteleaceae:领春木科; Acer:槭属;Cotinus:黄栌属; Morus:桑属;Cornaceae:山茱萸科;Rhus:盐肤木属; Carpinus/Corylus:鹅耳枥属/榛属;Salix:柳属;Euphorbiaceae:大戟科; Aquifoliaceae:冬青科; Rutaceae:芸香科; Securinega:叶底珠属; Ericaceae:杜鹃花科;Sambucus:接骨木属; Corylopsis:蜡瓣花属; Spiraea:绣线菊属; Rosaceae:蔷薇科; Mollotus:野桐属; Oleaceae:木樨科;Gramineae:禾本科; Artemisia:蒿属; Compositae:菊科;Chenopodiaceae:藜科; Cruciferae:十字花科;Fabaceae:蝶形花科;Labiatae:唇形科;Umbelliferae:伞形科;Ranunculaceae:毛茛科; Thalictrum:唐松草属; Humulus:葎草属; Urtica:荨麻属; Saxifraga:虎耳草属; Impatiens:凤仙花属;Cyperaceae:莎草科
结果显示,前两轴累积方差贡献率为19.5%(第一轴=12.1%,第二轴=7.4%)。第一轴正方向分布的样品主要为针阔混交林+针叶林+灌丛草甸,负方向为低山常绿阔叶林间人工次生林(图6),花粉类型正方向为灌木+针叶树,负方向为人工种植树+常绿阔叶树类型(图6),第一轴可能和温度变化相联系,这与峨眉山植物生长的影响因子有关,温度是控制植物分布的最主要因子之一,第一轴可能受海拔高度变化有关,反映峨眉山随海拔升高气温降低的规律。第二轴正方向为针阔混交林+常绿落叶阔叶混交林样品,花粉类型主要有枫杨属、榆属、柏科、枫香属、凤仙花属和山茱萸科等,负方向样品为低山常绿阔叶林间人工次生林+中山常绿落叶林,花粉类型为蝶形花科、血桐属、金缕梅科、黄栌属等,第二轴可能与水分条件变化有关,反映峨眉山山麓至山顶大气降水的差异。DCA表明,通过样品花粉百分含量,能较好地区分人工扰动林、阔叶林和针叶林,但常绿阔叶林、常绿落叶阔叶林和针阔混交林难以区分,这可能由于常绿阔叶林、常绿落叶阔叶林和针阔混交林样品中落叶阔叶树分布较广,花粉含量均较高的缘故;针叶林和灌丛草甸也较难区分,这可能源于灌丛草甸分布区原生植被为针叶林,只是后来遭人工砍伐次生演替所致,且灌丛草甸与周围冷杉林相距较近,受冷杉林影响明显,故其与冷杉林的花粉组合相似。
4 讨论
4.1 花粉组合与植被类型的关系
研究区植被类型主要为亚热带常绿阔叶林、常绿落叶阔叶混交林、针阔混交林、针叶林和灌丛草甸,其中亚热带常绿阔叶林由于受海拔高度和人类活动影响,又可划分为低山常绿阔叶林间人工次生林和中山常绿阔叶林。
1)低山常绿阔叶林间生人工次生林 花粉组合中落叶阔叶树占优势(22.3%—77.6%),以桤木属(2.7%—33.4%)、桦属(2.6%—20.3%)和盐肤木属(0—68.9%)为主,栗属(0—4.7%)、落叶栎类(0.2%—7.8%)、胡桃属(0.2%—6.0%)、枫杨属(0.2%—2.8%)、榆属(0—3.3%)、桑属(0—12.6%)、山茱萸科(0—14.4%)等含量较高。针叶树花粉含量(7.8%—62.5%)其次,以松属(3.0%—34.4%)和杉科(0.5%—53.6%)为主,且含有少量的柏科(0.1%—6.5%)。灌木花粉含量(1.5%—20.1%)较低,以蔷薇科(0—9.2%)为主。常绿阔叶树花粉含量(0.6%—12.3%)最低,主要见栲属/柯属(0—9.7%)和桃金娘科(0—5.5%)。草本花粉含量(4.0%—21.0%)较低,以禾本科(0.7%—8.1%)和蒿属(0.9%—6.4%)为主。水生草本植物花粉含量(0—2.3%)很低,约0.4%;蕨类孢子比重(4.3%—336.3%)较大,约54.3%,以水龙骨科(1.0%—57.8%)、凤尾蕨属(0—124.8%)和桫椤属(0—94.6%)为主。木本植物与草本植物花粉AP/NAP比值约为10.1;孢粉浓度191572粒/g。花粉组合中常绿阔叶树含量为4%,主要见栲属/柯属、常绿栎类和青冈属等,柏科、杉科、桃金娘科、栗属、桤木属、胡桃属、枫杨属、盐肤木属和禾本科等人工栽培植物花粉占约47.6%。群落中常绿阔叶优势种主要有樟科细叶楠、润楠、香樟等,样品中未发现其花粉,这可能与樟科花粉壁较薄很难保存下来有关[59];人工种植的林木树种主要有柏木、水杉、柳杉、桉树、板栗、水冬瓜、野核桃、枫杨、五倍子和慈竹等,其花粉含量约占一半,能较好地对应。故低山常绿阔叶林间生人工次生林花粉组合能较好地反映群落的人类活动痕迹,但未能指示母体植被的地带性特征。
2)中山常绿阔叶林 花粉组合中落叶阔叶树占优势(21.6%—39.8%),以桤木属(7.3%—16.4%)和桦属(4.2%—6.7%)为主。常绿阔叶树花粉含量(1.9%—4.7%)最低,主要见常绿栎类(0.2%—2.7%)和冬青科(0.4%—1.6%)。草本花粉含量(9.7%—55.5%)较高,以蒿属(2.7%—6.0%)、毛莨科(1.6%—27.4%)和虎耳草属(0—14.9%)为主。针叶树花粉含量(10.1%—35.7%)较低,以松属(5.8%—32.0%)和杉科(1.6%—6.7%)为主。灌木花粉含量(5.4%—38.3%)较低,以蔷薇科(1.6%—8.8%)和鹅耳枥属/榛属(0.6%—32.3%)为主。蕨类孢子含量(3.8%—21.1%)较低,以水龙骨科(0.8%—3.9%)和里白属(0.4%—9.5%)为主。木本植物与草本植物花粉AP/NAP比值约为4.3;孢粉浓度265198粒/g。花粉组合中常绿阔叶树含量仅为3.3%,主要见常绿栎类、冬青科、桃金娘科、栲属/柯属和青冈属等。群落分布的优势树种有壳斗科石栎属、栲属、青冈属,樟科桢楠属、润楠属、黄肉楠属、樟属、山胡椒属和木姜子属,山茶科木荷属等,花粉组合中仅可见优势种类型,不占优势,也不能反映群落总体特征。故中山常绿阔叶林花粉组合未能反映母体植被的植物组成特征。
3)常绿落叶阔叶混交林 花粉组合中落叶阔叶树占优势(12.2%—74.1%),以桤木属(5.4%—20.4%)、桦属(1.2%—9.0%)和枫杨属(0.1%—55.8%)为主。常绿阔叶树花粉含量(0.8%—6.9%)最低,主要见栲属/柯属(0—3.6%)、常绿栎类(0.4%—3.1%)和青冈属(0—1.6%)。针叶树、灌木和草本植物花粉含量相当,约19%,分别为20.1%、19.3%和18.9%。针叶树花粉含量(6.6%—31.7%)超过1/5,以松属(1.6%—18.5%)和杉科(2.1%—13.3%)为主。灌木花粉含量(3.3%—41.3%)略高于19%,以蔷薇科(0—40.6%)为主。草本花粉含量(12.0%—28.4%)略低于19%,以禾本科(0.3%—21.3%)、凤仙花属(0—14.3%)和蒿属(0—7.4%)为主。水生草本植物花粉含量约为0.1%;蕨类孢子含量(0.9%—25.8%)较高(8.4%),以水龙骨科(0.1%—17.1%)为主。木本植物与草本植物花粉AP/NAP比值约为4.7;孢粉浓度174652粒/g。花粉组合中阔叶树含量(41.7%)占优势,主要花粉类型有桤木属、桦属、枫杨属、松属、杉科、蔷薇科和禾本科等,柏科、栲属/柯属、落叶栎类、青冈属、落叶栎类花粉含量也较高;植物群落中优势种常绿阔叶树种有润楠、峨眉栲、包石栎和交让木等,落叶阔叶树种有华西枫杨、白辛树、木瓜红、鸡爪槭、稠李和珙桐等。花粉组合特征可以指示植物群落的外貌特征,但只能指示群落的部分优势种和建群种,难以反映母体植被中植物组成规律。
4)针阔混交林 花粉组合中落叶阔叶树占优势(13.9%—72.9%),以桤木属(3.3%—18.5%)和枫杨属(0.2%—65.1%)为主,桦属(0.4%—9.0%)、落叶栎类(0—2.9%)、胡桃属(0.5%—2.2%)和桑属(0—1.3%)等含量较高。针叶树花粉含量(12.9%—41.4%)其次,以松属(6.3%—24.4%)和杉科(1.0%—14.7%)为主,柏科(0.6%—9.6%)和冷杉属(0—2.6%)有一定含量。灌木花粉含量(3.8%—46.4%)再次,以蔷薇科(1.9%—35.2%)为主。常绿阔叶树花粉含量(0.4%—59.9%)最低,主要见栲属/柯属(0.2%—57.7%)和常绿栎类(0.2%—3.6%)。草本花粉含量(6.5%—36.9%)较低,以禾本科(0—6.6%)和蒿属(1.9%—4.4%)为主。水生草本植物花粉含量(0.4%)很低;蕨类孢子比例(5.8%)较低,以水龙骨科为主。木本植物与草本植物花粉AP/NAP比值约为8.4;孢粉浓度130794粒/g。花粉组合中乔木占绝对优势,达2/3,针叶树花粉以柏科、杉科、冷杉属和松属为主,阔叶树花粉主要有栲属/柯属、桦属、桤木属、枫杨属等,见槭属,灌木花粉主要有蔷薇科、绣线菊属,还可见杜鹃花科和柳属,禾本科花粉含量也较高,这和母体植被中主要分布冷杉、槭树、峨眉栲、全苞石栎、细齿稠李、峨眉蔷薇、悬钩子、糙皮桦、华西枫杨、金顶柳和方竹等对应很好。因此,针阔混交林花粉组合不但能很好地反映群落特征,而且花粉百分含量高的类型可以与母体植被中优势种很好地对应。
5)针叶林 花粉组合中针叶树含量(18.2%—40.5%)最高,以松属(7.3%—21.2%)、杉科(2.1%—13.0%)和冷杉属(0—12.9%)为主,见柏科(0—2.0%)和铁杉属(0.2—0.9%)。落叶阔叶树花粉含量(12.7%—37.5%)其次,以桤木属(7.2%—23.2%)和桦属(1.9%—10.0%)为主。灌木花粉含量(8.3%—54.9%)再次,以蔷薇科(1.5%—50.2%)为主。常绿阔叶树花粉含量(1.3%—7.1%)最低,主要见常绿栎类(0.6%—6.2%)。草本花粉含量(7.2%—28.5%)较低,以蒿属(1.3%—5.8%)为主,禾本科(0.2%—3.2%)、荨麻属(0—11.9%)和毛莨科(0—2.0%)有一定含量。蕨类孢子含量(4.6%)较低,以水龙骨科为主。木本植物与草本植物花粉AP/NAP比值为9.1;孢粉浓度126881粒/g。花粉组合中针叶树含量最高,约为30%,以松属为主,冷杉属、铁杉属、杉科和柏科含量较高,这能部分地反映母体植被中建群种冷杉和铁杉的分布。灌木花粉含量25%,主要有蔷薇科和绣线菊属,杜鹃花科和柳属含量也较高,禾本科也有一定含量,这与母体植被中峨眉蔷薇、高山绣线菊、杜鹃、高山柳、冷箭竹和大箭竹等灌木分布对应良好。花粉组合中针叶树+灌木+禾本科含量超过56%,这和群落中由于冷杉过熟,树木老朽,林间空隙较大,灌木生长茂盛的群落特征对应较好,但未能反映出优势种冷杉的主体特征。故针叶林花粉组合只能基本反映母体植被的群落特征。
6)灌丛草甸 花粉组合中灌木花粉含量(16.6%—41.8%)较高,平均25.3%,以蔷薇科(9.5%—38.4%)为主,见鹅耳枥属/榛属(0.8%—0.8%)、绣线菊属(1.1%—4.7%)和柳属(0.3%—0.6%)。草本花粉含量(18.1%—28.8%)较低,以禾本科(3.1%—7.7%)、蒿属(3.5%—7.1%)、毛莨科(0.5%—14.1%)和莎草科(0—14.8%)为主,见虎耳草属(0—0.6%)。落叶阔叶树花粉含量(21.0%—39.4%)最高,以桤木属(13.3%—26.1%)和桦属(1.9%—10.0%)为主。常绿阔叶树花粉含量(3.4%—4.9%)最低,主要见常绿栎类(2.2%—4.1%)和青冈属(0—1.1%)。针叶树含量(9.6%—27.4%)较低,以松属(6.7%—20.3%)为主,见冷杉属(0—3.3%)。蕨类孢子比例(5.6%)较低,以水龙骨科为主。木本植物与草本植物花粉AP/NAP比值为3.2;孢粉浓度202673粒/g。花粉组合中灌木和草本植物含量超过一半,灌木花粉以蔷薇科和绣线菊属为主,柏科、冷杉属、杜鹃花科、柳属和接骨木属等有一定含量,这和植物群落中主要分布峨眉蔷薇、微毛野樱桃、陕甘花楸、高山柏、金顶柳、杜鹃、冷杉幼苗、蒴藋和银叶委陵菜能很好地对应。草本花粉以禾本科、唐松草属和虎耳草科为主,这和母体植被中冷箭竹、茶藨子、亚欧唐松草等草本植物生长茂盛相对应。花粉组合中含量较高的类型与母体植被中建群种、优势种对应良好,这表明灌丛草甸的花粉组合能够较好地反映母体植被的群落组成特征。
4.2 主要花粉类型的散布特征
冷杉属花粉 E22—E38号样品样点周围有冷杉分布,冷杉属花粉含量2.9%左右;其他样点附近未见冷杉,其花粉含量0.02%左右,低于0.2%。其中冷杉林中冷杉属花粉含量为5.3%,E35号样品其含量最高,达12.9%;针阔混交林中其含量为0.8%,E29号样品其含量更高,为2.6%。冷杉属花粉具低代表性,这和前人在吉林长白山和陕西秦岭研究结果相一致,R值介于0.1—0.5之间,其花粉含量低于20%[60-61];四川螺髻山和湖北神农架地区研究也显示,冷杉属花粉代表性很好,随冷杉优势度下降其花粉含量迅速降低,无冷杉分布时仅见个别冷杉属花粉[62-63]。所以,样品中5%以上含量冷杉属花粉的出现,就可指示冷杉林的存在;其0.8%以上含量可以表明样点周围冷杉的分布;其0.2%以下含量不能确定样点附近有冷杉生长。
杜鹃花科花粉 E16、E20和E22—E40号样品样点周围有杜鹃分布,杜鹃花科花粉含量约0.2%左右;其余样品样点附近未见杜鹃分布,其花粉含量仅为0.03%。灌丛草甸杜鹃花科花粉含量约0.07%,最高可达0.2%;乔木林下其花粉含量约0.2%,最高可达1.4%。杜鹃花科花粉代表性很低,R值为0.79—0.83[64-65],其花粉产量较低,传播能力较弱[62],受外来花粉百分比递减补偿作用影响较大[61],其0.2%以上的花粉含量就能反映样点附近杜鹃的生长。
蔷薇科花粉 E9、E14—E15、E18—E20、E22—E23、E27—E35、E38—E40号等样品样点中周围有蔷薇、悬钩子、花楸和峨眉蔷薇等生长,蔷薇科花粉含量约17.7%;其余样品样点附近未见其母体植被分布,其花粉含量约3.5%。灌丛草甸中峨眉蔷薇和微毛野樱桃呈优势种群分布,其覆盖度平均可达40%左右,蔷薇科花粉含量约20%,最高可达38%,R值约为0.5;乔木林下蔷薇作为林间植物分布,其花粉含量约17.4%。甘肃兴隆山研究显示蔷薇科花粉具低代表性[66],在蔷薇灌丛群落中代表性较低[67],在山体样品花粉组合中其花粉的发现能够反映母体植被的生长[68],这可能由于蔷薇科是虫媒植物,花粉产量低所致[69]。
珙桐属花粉 E16—E19和E22号样品样点周围有珙桐树生长,但仅在E18号样品中检测到1粒珙桐属花粉,且E16样品采自珙桐树开花后不久的大树荫下苔藓,也未发现其花粉。这可能源于珙桐属花粉外壁比较薄,仅2—3 μm[70],实验处理易破碎,或者由于保存环境相对湿度大易氧化,这可能导致其在花粉组合中发现较少。故珙桐属花粉具很低代表性,珙桐属花粉的出现即指示样点周围有珙桐分布。
槭属花粉 E15—E25,E27—E33号样品样点周围分布长尾槭、青榨槭、杈叶槭、扇叶槭、疏花槭、五尖槭、五裂槭、中华槭等多种槭树分布,槭属花粉含量约为0.3%,最高为1.6%;其余样品样点附近未见槭树生长,其花粉含量约为0.09%。槭属花粉代表性很低,R值0.23—0.38[64-65],其1%以上的花粉含量就能够反映样点周围母体植物的存在。
盐肤木属花粉 E04、E06—E11、E20、E22和E25号样品样点周围有五倍子树分布,盐肤木属花粉含量约8.6%,其中E10号样品采自五倍子树林底下苔藓,盐肤木属花粉含量高达68.9%,样品附近零星生长五倍子树,其花粉含量约为2%;其余样品样点周围未见五倍子树生长,其花粉含量约为0.3%,个别含量高约1.5%。盐肤木属花粉具低代表性,R值介于0.6—0.7之间[64-65],其花粉2%以上含量就可指示样点周围五倍子树的存在,60%以上含量表明样点附近五倍子林的分布。
枫杨属花粉 所有样品中均可见枫杨属花粉,其含量介于0.1%—65.1%之间,平均4.2%。样品周围无枫杨生长时其花粉含量约为1.3%,个别样品含量能达到3.7%;E17和E26号样品采自枫杨林树底下,其花粉含量分别为55.8%和65.1%。枫杨属花粉具超代表性,5%以下含量时不能确定样点附近有枫杨分布,50%以上含量时能够指示样品来自于枫杨林。也有研究认为枫杨属具低代表性,R值为0.64—0.69之间[64-65]。
栲属/柯属花粉 E16—E20和E22—E24号样品样点周围有峨眉栲或硬壳柯分布,其花粉含量约为8.3%,其中E24号样品采自峨眉栲林,其花粉含量高约57.7%;其余样品附近未见其母体植物分布,但花粉组合中大多数可见栲属/柯属花粉的出现,含量约为0.9%。因此,栲属/柯属花粉具超代表性,1%以下花粉含量不能确定样点周围母体植物的存在,50%以上花粉含量能够指示样点附近有栲树林的分布。
桤木属花粉 所有样品中均有桤木属花粉(2.7%—33.4%)出现,平均约14.6%。E4—E6号样品样点周围有人工种植的水冬瓜生长,其花粉含量约为26.7%;其余样品样点附近未见其母体植被分布,其花粉含量约为13.6%。桤木属花粉具超代表性,R值为1.16[71];重庆缙云山样点周围没有母体植物分布,其花粉均有出现(2.8%—10.5%)[72];昆明西山桤木属花粉含量为9.0%—21.4%,样点附近400 m2母体植被中无该植物分布,认为桤木属花粉不但产量大[73],而且具有较强的传播能力[73-74],本研究和前人结果相一致。
杉科花粉 所有样品均有杉科花粉(0.5%—53.6%)出现,平均7.5%。E01—E08、E11、E14和E17—E22号等样品样点周围有柳杉或(和)杉木生长,其花粉含量约11.6%,其中E02—E04号样品采自柳杉林中,其花粉含量约为41.1%;其余样品样点附近未见其母体植物分布,其花粉含量为4.9%。因此,杉科花粉具超代表性,40%以上花粉含量表明样点周围母体植物纯林分布,10%以上含量指示样点附近母体植被的分布,5%以下含量不能确定样点周围母体植物的生长。
4.3 孢粉组合的群落生态和古环境意义

图7 表土花粉类型(Ⅰ)Fig.7 Types of surface pollen (Ⅰ)1. 里白属Hicriopteris(样号E09),2. 卷柏属Selaginella(样号E08),3、4、13. 金星蕨科Thelypteridaceae(样号E08、E09、E10),5、7—10. 水龙骨科Polypodiaceae(样号E01、E02、E03);6、12. 中国蕨科Sinopteridaceae(样号E15),11. 桫椤科Cyatheaceae(样号E09),14、15. 凤尾蕨属Pteris(样号E02、E09). 图示标本保存于吉首大学湖南省生态旅游重点实验室
峨眉山孢粉组合与其母体植被特征对比显示:中山常绿阔叶林花粉组合未能反映植物的群落特征;低山常绿阔叶林间生人工次生林和常绿落叶阔叶混交林花粉组合只能反映母体植被的部分植物组成;针叶林花粉组合基本可以指示母体植被的群落特征;灌丛草甸花粉组合能够较好地反映母体植被的群落组成特征;针阔混交林花粉组合不仅可以很好地指示群落特征,花粉高含量类型还可以与植物群落的优势种很好的对应。导致产生上述结果的原因可能是:(1) 由于受峨眉山地形地势限制,多数地段坡陡崖直,通行困难,所以,大部分样品只能沿着现有山路两侧10—20 m距离内采集,难以采用100 m×100 m乃至更大样方,很难均匀地覆盖整个山体,导致部分集群分布的种类难以区分开来,如华西枫杨群落和峨眉栲群落中枫杨属和栲属花粉含量较之邻近样品高出很多。(2) 由于峨眉山地处华西雨屏之南端[75],山麓至山顶年平均气温较高,低云、多雾、雨量充沛,样点附近灌木和草本分布非常茂密,部分地段其盖度达50%—70%,个别样点附近草本植物尤其繁盛,其花粉含量较高,稀释了木本植物花粉含量。(3) 由于峨眉山开发历史悠久,不论是山麓地区,还是山顶地带,都深深地打上了人类活动的烙印;由山麓至山顶沿途种植了大量栽培林木,主要有柳杉、水冬瓜、马尾松、华西枫杨、糙皮桦等,这使峨眉山植被分布带有很强的斑状性质。(4) 峨眉山植物种类极为丰富,现有高等植物约280科3703种[47],其中种子植物就达2400种[46],孢粉鉴定虽然平均每样约756粒以上,部分样品统计达1709粒,但仍然难以囊括群落所有类型,这也是孢粉组合与群落中植物组成对应关系复杂的原因。(5) 受上升气流的影响,山麓地区花粉被风力沿坡逐次向上携带至山顶,使得山顶花粉组合受到山麓地带人工栽培种类干扰,样品中见大量松属和桤木属花粉。

图8 表土花粉类型 (Ⅱ)Fig.8 Types of surface pollen (Ⅱ)1—3. 桤木属Alnus(样号E05、E06、E07),4. 桦属Betula(样号E30),5. 胡桃属Juglans(样号E20),6. 榆属Ulmus(样号E30),7、8. 青檀属Pteroceltis(样号E12、E15),9. 山黄麻属Trema(样号E15),10—15. 蔷薇科Rosaceae(样号E06、E15、E19、E23、E28),16、17. 绣线菊属Spiraea(样号E30、E38),18. 栲属/柯属Castanopsis/Lithocarpus(样号E24),19、23. 松属Pinus(样号E16、E36),20. 杉科Taxodiaceae(样号E05),21. 柏科Cupressaceae(样号E01),22. 铁杉属Tsuga(样号E36),24. 冷杉属Abies(样号E36)
峨眉山常绿阔叶林樟科群落是热带亚热带地区代表类群之一,现已查明约有樟科11属60种[47],占四川现有樟科分布14属的78.6%,96种的62.5%[45],占中国樟科分布20属的55%,423种的14.2%,占全世界樟科分布45属的1/4[59]。因此,常绿阔叶林樟科类群的研究具有典型意义。由于樟科花粉外壁薄,易破碎[59],

图9 表土花粉类型 (Ⅲ)Fig.9 Types of surface pollen (Ⅲ)1、2. 青冈属Cyclobalanopsis(样号E15、E22),3、4. 叶下珠属Phyllanthus(样号E17、E20),5、6. 叶底珠属Securinega(样号E13、E18),7. 栎属Quercus(样号E16),8. 山麻杆属Alchornea(样号E15),9. 楝属Melia (样号E10),10. 山矾属Symplocos(样号E16),11、18. 豆科Leguminosae(样号E11、E15),12、19. 七叶树属Aesculus(样号E15),13. 桑属Morus(样号E18),14. 棕榈科Palmae(样号E15),15. 野桐属Mallotus(样号E15),16. 蒿属Artemisia(样号E20),17. 唐松草属Thalictrum(样号E28),20、21. 卫矛科Celastraceae(样号E15、E19),22、41. 伞形科Umbelliferae(样号E18、E22),23. 柳属Salix(样号E40),24、25. 十字花科Cruciferae(样号E20、E25),26、30. 水丝梨属Sycopsis(样号E08、E15),27. 八角属Illicium(样号E24), 28、29. 山茱萸属Cornus(样号E01、E15),31. 蓼科Polygonaceae(样号E08),32. 蓼属Polygonum(样号E10),33. 芸香科Rutaceae(样号E07),34. 盐肤木属Rhus(样号E10),35—38. 杜英科Elaeocarpaceae(样号E10),39. 藜科Chenopodiaceae(样号E19),40. 唇形科Labiatae(样号E15),42. 杜鹃花科Ericaceae(样号E37),43. 凤仙花属Impatiens(样号E22)
此次所有样品中均未检测到,这也是为什么利用孢粉学构建古气候缺少热带亚热带代表类群樟科的原因。但有学者在晚白垩纪至始新世新泽西州、北卡罗来纳州以及北美等地层中发现Perseanthus属、Neusenia属和Androglandula属等原位花粉化石[76-78],早期樟科花粉基本无萌发孔,花粉大小差异较大,Androglandula属花粉直径约16 μm左右,Perseanthus属和Neusenia属则在28—34 μm之间,此外,Perseanthus属花粉壁上具小刺。未来改良样品前处理方法和研究花粉保存环境,提高地层中孢粉化石鉴定统计精度,以及结合植物大化石[79]、分泌细胞[80]和气孔器[81]等其他遗留特征,可能有助于地质时期樟科类群的恢复。
珙桐为中国特有珙桐科单型属植物,是第三纪古热带植物区系的孑遗种,晚白垩纪和第三纪时期曾广布世界许多地区,中国亚热带地区也曾有普遍分布,但现今仅在中国西南及长江中上游地区有所残存[82]。峨眉山是珙桐在四川分布最集中和最典型的地区之一,珙桐作为中亚热带中山山地常绿落叶阔叶混交林的主要优势种和建群种[83],在植物系统发育和地史变迁研究中有很高的学术地位[84],江西省地层分析曾发现过珙桐的孢粉材料[85]。本研究5个样品采自珙桐群落,只有1个样品中发现1粒珙桐属花粉。因此,改良样品实验分析方法以及花粉保存环境的分析,改善样品中珙桐花粉的鉴定精度,提高其花粉统计效率,可为植物发展演化、地质变迁和环境演变研究提供花粉学证据。
峨眉山针叶冷杉纯林由于林分结构不同,群落特征差异较大,天门寺至金顶冷杉植株较小,多数为幼年期和青年期植株,其繁殖能力强,花粉产量大,树冠常常可见紫色球果,样品中冷杉属花粉含量可达12.3%,孢粉浓度为26534粒/g;万佛顶、太子坪至接引殿分布冷杉过熟林,植株高大,林龄较大,老者可超过500年以上,处于衰老期,生命力较弱,近期由于病虫害入侵[86],大量冷杉衰亡,枯死,导致冷杉花粉产量锐减,其百分含量只有2.9%,花粉浓度约1978粒/g。因此,某种植物在相同植被盖度条件下,较大差异的花粉产量在一定程度上往往可以指示其不同的林分结构特征。所以,峨眉山针叶冷杉纯林中冷杉属花粉含量较低,除了受其花粉低代表性影响,还与峨眉山主要为冷杉过熟林,花粉产量相对较低有关,致使花粉组合难以反映冷杉的建群种特征。
因此,在植物种类非常丰富的热带亚热带地区,尤其是其代表性类群花粉代表性低的山地,在利用孢粉学资料进行植被生态保护与恢复和地质历史时期气候与环境反演与重建时,应该谨慎分析地层孢粉数据,尽量与其他生物环境指标 (植物大化石、分泌细胞和气孔器等) 进行综合分析,力求获得相互支持,相互印证的准确结论。
5 结论
通过峨眉山40个样点表土孢粉组合及其与植物群落之间关系的分析,结果表明:
(1) 花粉组合中木本植物含量(83.3%)占绝对优势,木本植物花粉与草本植物花粉比值(AP/NAP)约为5,松属、杉科、桤木属、蔷薇科、桦属、枫杨属、蒿属、毛茛科和水龙骨科为主要孢粉类型;孢粉浓度174003粒/g。
(2) 中山常绿阔叶林花粉组合未能反映植物的群落特征;低山常绿阔叶林间人工次生林和常绿落叶阔叶混交林花粉组合只能反映母体植被的部分植物组成;针叶林花粉组合基本可以指示母体植被的群落特征;灌丛草甸花粉组合能够较好地反映母体植被的群落组成特征;针阔混交林花粉组合不仅可以很好地指示群落特征,花粉高含量类型还可以与植物群落的优势种很好的对应。
(3) 主要花粉类型冷杉属、杜鹃花科、蔷薇科、珙桐属、槭属和盐肤木属具低代表性;枫杨属、栲属/柯属、桤木属和杉科花粉具超代表性。
(4) DCA表明,通过样品花粉百分含量,能较好地区分人类扰动植被、阔叶林和针叶林,但常绿阔叶林、常绿落叶阔叶混交林和针阔混交林之间,针叶林和灌丛草甸之间难以区分。
(5) 热带亚热带常绿阔叶林代表类群樟科和常绿落叶阔叶林珙桐群落受其主体植物花粉外壁薄,易破碎的影响,利用孢粉学恢复原生植被时缺失优势种;因此探索孢粉实验处理改良、孢粉保存环境研究,与其他生物学指标(植物大化石和气孔器)相结合重建古植物群落具有重要意义。
(6) 不同林型的冷杉纯林,幼苗期因繁殖力强,花粉产量高;过熟林由于衰老、病虫害影响,繁殖率下降,花粉产量低,这表明植物花粉含量一定程度上可以反映其林分结构。
本研究可为热带亚热带山地及相似地区利用孢粉学进行地质时期气候与环境重建提供理论支持和基础资料,并对植被生态恢复提供实例和参考。
