MiR-19a-3p regulates the Forkhead box F2-mediated Wnt/β-catenin signaling pathway and affects the biological functions of colorectal cancer cells
2020-03-13FuBingYuJuanShengJiaManYuJingHuaLiuXiangXinQinBoMou
Fu-Bing Yu, Juan Sheng, Jia-Man Yu, Jing-Hua Liu, Xiang-Xin Qin, Bo Mou
Abstract BACKGROUND Colorectal cancer (CRC) is one of the most common malignancies worldwide.AIM To explore the expression of microRNA miR-19a-3p and Forkhead box F2(FOXF2) in patients with CRC and the relevant mechanisms.METHODS Sixty-two CRC patients admitted to the hospital were enrolled into the study group, and sixty healthy people from the same period were assigned to the control group. Elbow venous blood was sampled from the patients and healthy individuals, and blood serum was saved for later analysis. MiR-19a-3p mimics,miR-19a-3p inhibitor, miR-negative control, small interfering-FOXF2, and short hairpin-FOXF2 were transfected into HT29 and HCT116 cells. Then quantitative polymerase chain reaction was performed to quantify the expression of miR-19a-3p and FOXF2 in HT29 and HCT116 cells, and western blot (WB) analysis was conducted to evaluate the levels of FOXF2, glycogen synthase kinase 3 beta (GSK-3β), phosphorylated GSK-3β (p-GSK-3β), β-catenin, p-β-catenin, α-catenin, Ncadherin, E-cadherin, and vimentin. The MTT, Transwell, and wound healing assays were applied to analyze cell proliferation, invasion, and migration,respectively, and the dual luciferase reporter assay was used to determine the correlation of miR-19a-3p with FOXF2.RESULTS The patients showed high serum levels of miR-19a-3p and low levels of FOXF2,and the area under the curves of miR-19a-3p and FOXF2 were larger than 0.8.MiR-19a-3p and FOXF2 were related to sex, tumor size, age, tumor-nodemetastasis staging, lymph node metastasis, and differentiation of CRC patients.Silencing of miR-19a-3p and overexpression of FOXF2 suppressed the epithelialmesenchymal transition, invasion, migration, and proliferation of cells. WB analysis revealed that silencing of miR-19a-3p and FOXF2 overexpression significantly suppressed the expression of p-GSK-3β, β-catenin, N-cadherin, and vimentin; and increased the levels of GSK-3β, p-β-catenin, α-catenin, and Ecadherin. The dual luciferase reporter assay confirmed that there was a targeted correlation of miR-19a-3p with FOXF2. In addition, a rescue experiment revealed that there were no differences in cell proliferation, invasion, and migration in HT29 and HCT116 cells co-transfected with miR-19a-3p-mimics+sh-FOXF2 and miR-19a-3p-inhibitor+si-FOXF2 compared to the miR-negative control group.CONCLUSION Inhibiting miR-19a-3p expression can upregulate the FOXF2-mediated Wnt/βcatenin signaling pathway, thereby affecting the epithelial-mesenchymal transition, proliferation, invasion, and migration of cells. Thus, miR-19a-3p is likely to be a therapeutic target in CRC.
Key words: MiR-19a-3p; Forkhead box F2; Wnt/β-catenin signaling pathway; Biological function; Colorectal cancer; Western blot
INTRODUCTION
Colorectal cancer (CRC) is a malignant tumor of the digestive system, also known as the gastrointestinal tract, with an incidence second only to gastric cancer. With the development of the social economy, an increasing number of young people suffer from CRC due to unhealthy living and eating habits[1,2]. Early detection, diagnosis, and radical therapy are pivotal to the treatment of CRC, but most patients already have advanced-stage disease at the time of diagnosis due to the insidious clinical characteristics in the early stage; thus, CRC patients suffer a poor prognosis and extremely high mortality[3,4]. At present, CRC is mainly treated with surgical resection,radiotherapy, or chemoradiotherapy in clinical practice[5,6]. In recent years, based on in-depth studies on the mechanism of CRC, it has been found that the regulation of tumor-suppressor or tumor-promoting genes can affect the occurrence and development of tumors, thereby offering novel insights into CRC therapy and prognosis[7]. Studies on tumors have concluded that the invasion and metastasis of cancer cells are strongly linked to activation of the epithelial-mesenchymal transition(EMT) pathway as follows. With the EMT, tumor epithelial cells have enhanced invasion and migration ability, transform into mesenchymal-like cells, and then fall off from the primary foci to form new metastatic foci. The EMT of tumors involves a series of complex molecular events and regulatory pathways[8]. MicroRNA (miRNAs),as endogenous non-coding single-stranded RNAs with cancer-promoting or cancerinhibiting functions, may participate in the EMT of tumors through a variety of signaling pathways[9]. Studies have shown that miR-19a-3p is an miRNA with abnormal expression in a variety of tumor cells[10]. For example, a study revealed that highly expressed miR-19a-3p could regulate cell adhesion molecule 2 and affect the proliferation and migration mechanism of renal carcinoma cells through the AKT signaling pathway[11]. In addition, a study found that miR-19a-3p reversed the tumorassociated macrophage phenotype by regulating Fos-related antigen 1/IL-6/signal transducer and activator of transcription 3 signaling pathways, thus inhibiting breast cancer metastasis[12]. Although these studies have demonstrated that miR-19a-3p is extensively involved in cancer such as renal cell carcinoma and breast cancer, the specific mechanism underlying its role in CRC is not completely understood[13,14].Forkhead box F2 (FOXF2) is a homeobox protein and an inducer of the EMT. A previous study showed that FOXF2 can promote the EMT of CRC cells by mediating the Wnt/β-catenin signaling pathway[15]. Wnt/β-catenin is a classical signaling pathway in the pathological process of cancer, which is overexpressed in tumor cells and has a very close relationship with their proliferation and apoptosis[16].
We predicted that FOXF2 is probably the target site of miR-19a-3p using online biological prediction software (http://www.targetscan.org/vert_72/). Based on the abovementioned studies, the goal of this study was to determine if miR-19a-3p influences the biological functions of CRC cells by targeting the FOXF2-mediated Wnt/β-catenin signaling pathway.
MATERIALS AND METHODS
Clinical data
Sixty-two CRC patients in the Fourth Affiliated Hospital of Kunming Medical University between March 2017 and July 2018 were enrolled as the study group, and sixty healthy people from the same period were enrolled as the control group. There were 32 males and 30 females (57.8 ± 3.80 years-old) in the study group, and 34 males and 26 females (57.5 ± 3.50 years-old) in the control group. Though comparable, there were no significant differences regarding sex and age between groups. The inclusion criteria of the study were: CRC patients with confirmed disease based on pathological diagnosis[17], and patients with expected survival longer than 3 mo. The exclusion criteria were: Patients who received preoperative chemoradiotherapy and immunotherapy. All patients and their families signed an informed consent form after understanding this study, and the study was conducted with permission from the Ethics Committee of the Fourth Affiliated Hospital of Kunming Medical University.
Experimental reagents and materials
The human normal colonic epithelial cell line and human CRC cell lines (HT29,SW480, SW620, and HCT116) (Nos. BNCC338003, BNCC337731, BNCC100604,BNCC337664, and BNCC337692; Beijing BeNa Culture Collection, Beijing);TransScript/TransScript IIGreen miRNA Two-Step qRT Polymerase Chain Reaction(PCR) SuperMix (AQ202-01/AQ301-01; Beijing TransGen Biotech, Beijing, China);MTT Kit (SY0502; Beijing Biolab Technology Co., Ltd., Beijing, China); Trizol reagent(10296010; Invitrogen Company, Carlsbad, CA, United States); Dual Luciferase Reporter Gene Determination Kit (KFS303; Beijing Biolab Technology); fetal bovine serum (FBS), Transwell Kit, and phosphate-buffered saline (PBS) (1142802, 10010049,10437028; Gibco Company, Gaithersburg, MD, United States), radioimmunoprecipitation assay (JN0190; Beijing Biolab Technology); Bicinchoninic Acid Protein Assay Kit (A53225; Thermo Fisher Scientific, Waltham, MA, United States);FOXF2, glycogen synthase kinase 3 beta (GSK-3β), phosphorylated GSK-3β (p-GSK-3β), β-catenin, p-β-catenin, α-catenin, N-cadherin, E-cadherin, vimentin, and β-actin antibodies (Cell Signaling Technology, Danvers, MA, United States); goat anti-rabbit IgG secondary antibody (Wuhan BOSTER Biological Technology Co., Ltd., Wuhan,China); electrochemiluminescence developer (Thermo Fisher Scientific), and PCR instrument (7500; Applied Biosystems, Waltham, MA, United States). The design and synthesis of all primers were carried out by Shanghai Sangon Biotech Co., Ltd.(Shanghai, China).
Cell culture, passage, and transfection
CRC cell lines were incubated in Dulbecco's Modified Eagle's Medium containing 10% PBS in an incubator with CO2at 37 °C for transfection. When cell growth and fusion under transfection reached 50%, 25% pancreatin was added to the cell lines for digestion, and after digestion, the cell lines were cultured in medium again to complete passage. Cells in the logarithmic phase were grouped and then transfected with an inhibition sequence (miR-19a-3p-inhibitor), overexpression sequence (miR-19a-3p-mimics), miR negative control (miR-NC), targeted inhibited FOXF2 RNA (si-FOXF2), targeted overexpressed FOXF2 RNA (sh-FOXF2), and NC RNA with the Lipofectamine™ 2000 kit, in accordance with the instructions of the corresponding kit.
Determination methods
Quantitative PCR assay: The quantitative PCR (qPCR) assay was employed to determine the level of mRNA in serum and cells. Total serum RNA was extracted from the patients as per the operating instructions of Trizol reagent and dissolved in 20 μL diethylpyrocarbonate water, followed by reverse transcription using a reverse transcription kit in 15 μL total reaction volume containing 1 μL M-MLV, 1 μL Oligo (d T), 0.5 μL RNA enzyme inhibitor, 1 μL NTP, and RNAse-free water to adjust the volume. The RNA was incubated at 38 °C for 60 min. cDNA (1 μL) was sampled and synthesized at 85 °C for 5 s. The synthesized cDNA was adopted as a qPCR amplification template. The PCR reaction system was prepared to a 25 μL total reaction volume with 2.5 μL of 10 × PCR buffer, 1 μL dNTPs, 1 μL upstream and downstream primers, 0.25 μL Taq DNA Polymerase, and dd H2O to adjust the volume. The reaction conditions were as follows: 95 °C for 15 min, 95 °C for 15 s, 58 °C for 30 min (a total of 35 cycles), and then 72 °C for 15 min. Three replicate wells were made for each sample for three repeated experiments. U6 was applied as an internal reference for miR-19a-3p, and β-actin as an internal reference for FOXF2. After reaction, the amplification and dissociation curves of the qPCR were confirmed, and the relative quantity of target gene was calculated based on obtained parameters. The relative quantification of target genes was analyzed using 2-ΔCt.
Western blot assay:Cultured cells were collected from each group, and the total protein was extracted using the radioimmunoprecipitation assay lysis method. The bicinchoninic acid method was adopted to determine the level of total protein, and was adjusted to 4 μg/μL. The total protein was transferred to a PVDF membrane through 6%-12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis.Corresponding protein band was selected according to the target protein, and the membrane was blocked in 5% skim milk for 2 h, followed by the addition of primary antibody (Beyotime Biology Co., Ltd., Jiangsu, China) at a dilution of 1:1000. Then the membrane was washed and stored at 4 °C for one night. The next day, the membrane was incubated with secondary antibody (Beyotime Biology Co., Ltd.) for 1 h with the same procedures, after which developer was added in a dark room. The PVDF membrane was imaged by a Tocan240 motored molecular imaging system (Shanghai Tocan Bio-technology Co., Ltd., Shanghai, China), and the imaging results were analyzed using Image LabTM software.
MTT assay:Cells transfected for 24 h were collected, and seeded into a 96-well plate at 5 × 103cells/well, followed by the addition of 20 μL of MTT solution (5 μg/mL) at 24, 48, and 72 h after being incubated at 37 °C. Then the cells were cultured at 37 °C for 4 h. A total of 200 μL dimethyl sulfoxide was added to each well, and then the optical density of each group of cells under 570 nm was measured using a spectrophotometer.
Transwell assay:Cells transfected for 24 h were collected and seeded into a 24-well plate at 3 × 104cells/well. The cells were digested with pancreatin, and transferred to the upper compartment. The upper compartment was added with 200 μL of RPMI 1640 culture solution, and the lower compartment was added with 500 mL of RPMI 1640 containing 10% FBS. The plate was cultured at 37 °C for 48 h, and the substrate and cells not passing through the microporous membrane in the upper chamber were removed. The plate was washed three times with PBS, followed by immobilization using paraformaldehyde for 10 min, and then washed three times with double distilled water. Subsequently, 0.5% crystal violet was employed to stain the dry plate,and cells invasion was evaluated using a microscope.
Wound healing assay for migration determination of cells in vitro
The cells were seeded into a 6-well plate after being diluted to 3 × 105cells/mL. When the confluency of the cells was up to 85%, a 200 μL sterile loading gun was used to divide the cells to form a cell-free area in the center of the culture plate. The divided cells were cultured in new culture medium after being washed with PBS. At 0 h (W0)and 24 h (W24) after cell division, the cell migration ability was evaluated according to wounds at three different sites using a microscope.
Xenotransplantation tumor model
Stable miR-19a-3p-inhibitor and 3 × 106HCT116 cells transfected with control plasmid were subcutaneously injected into the left abdomen of female BALB/c nude mice (4 wk old) raised in a sterile environment (5 nude mice in each group). The tumor growth of each mouse was evaluated every 7 d, and at 28 d after injection, the mice were executed by cervical dislocation, and their tumor size and mass in vivo were measured.
Statistical analyses
The collected data were analyzed statistically with SPSS20.0, and visualized with GraphPad 7. Inter-group comparison was conducted using the independent-samples t-test, and multi-group comparisons were conducted by the one-way analysis of variance, and expressed as F. Post hoc pairwise comparison was carried out using the least significant difference t-test, and comparison of data at multiple time points was carried out using the variance of repeated measures, and expressed as F. Post test was performed using Bonferroni. Receiver operating characteristic (ROC) curves of miR-19a-3p and FOXF2 and the value of diagnosing CRC was drawn. Pearson's correlation analysis was carried out to investigate the relationship between serum miR-19a-3p and serum FOXF2. P < 0.05 indicated a significant difference.
RESULTS
Levels of serum miR-19a-3p and FOXF2 and their clinical value
The determination of the levels of serum miR-19a-3p and FOXF2 in the subjects revealed that the study group had a significantly higher level of serum miR-19a-3p,and a significantly lower level of serum FOXF2 than the control group (both P <0.001). Pearson's correlation analysis revealed that the level of serum miR-19a-3p in CRC patients was negatively correlated with that of serum FOXF2 (P < 0.001), and the ROC curves showed that the area under the curves (AUC) of miR-19a-3p and FOXF2 were 0.883 and 0.850, respectively. Analysis of the correlation of miR-19a-3p and FOXF2 with pathological data of the patients revealed that the two indexes were strongly linked to age, sex, tumor size, differentiation, tumor-node-metastasis (TNM)staging, and lymph node metastasis (LNM) (all P < 0.05; Table 1 and Figure 1).
Effects of miR-19a-3p on the proliferation, invasion, migration, and EMT of CRC cells
The determination of miR-19a-3p expression in CRC cells revealed that miR-19a-3p expression in the human normal colonic epithelial cell line was significantly higher than that in human CRC cell lines (HT29, SW480, SW620 and HCT116) (P < 0.05).Transfection of miR-19a-3p-mimics, miR-19a-3p-inhibitor, and miR-NC into HCT116 and HT29 cells led to the following results. HCT116 and HT29 cells transfected with miR-19a-3p-mimics had significantly higher miR-19a-3p expression than those transfected with miR-NC, and HCT116 and HT29 cells transfected with miR-19a-3pinhibitor had significantly lower miR-19a-3p expression than those transfected with miR-NC. Determination of the biological functions of cells in the two groups demonstrated that cells with transfected miR-19a-3p-mimics had significantly more cell proliferation, invasion, and migration abilities than those transfected with miRNC and also had significantly higher expression of p-GSK-3β, β-catenin, N-cadherin,and vimentin proteins, and significantly lower expression of GSK-3β, p-β-catenin, acatenin, and E-cadherin. In addition, cells transfected with miR-19a-3p-inhibitor had significantly weaker cell proliferation, invasion, and migration abilities than those transfected with miR-NC, and also had significantly higher expression of p-GSK-3β,β-catenin, N-cadherin, and vimentin and significantly lower expression of GSK-3β, pβ-catenin, α-catenin,, and E-cadherin (Figure 2).
Effects of FOXF2 on the proliferation, invasion, migration, and EMT of CRC cells
The determination of FOXF2 expression in CRC cells revealed that human CRC cell lines (HT29, SW480, SW620 and HCT116) showed significantly lower expression of FOXF2 than the human normal colonic epithelial cell line (FHC) (P < 0.05).Transfection of si-FOXF2, sh-FOXF2, and miR-NC into HCT116 and HT29 cells transfected with si-FOXF2 showed significantly lower FOXF2 expression than those transfected with NC, while HCT116 and HT29 cells transfected with sh-FOXF2 showed significantly higher FOXF2 expression than those transfected with NC.Determination of the biological functions of cells in the two groups revealed that cells transfected with si-FOXF2 showed significantly stronger cell proliferation, invasion,and migration abilities than those transfected with NC, and also showed significantly higher expression of p-GSK-3β, β-catenin, N-cadherin, and vimentin, and significantly lower expression of GSK-3β, p-β-catenin, α-catenin,, and E-cadherin than those transfected with NC. In addition, cells transfected with sh-FOXF2 showed significantly weaker cell proliferation, invasion, and migration abilities than thosetransfected with NC, and also showed significantly lower expression of p-GSK-3β, βcatenin, N-cadherin, and vimentin and significantly higher levels of GSK-3β, p-βcatenin, α-catenin, and E-cadherin than those transfected with NC (all P < 0.05; Figure 3).
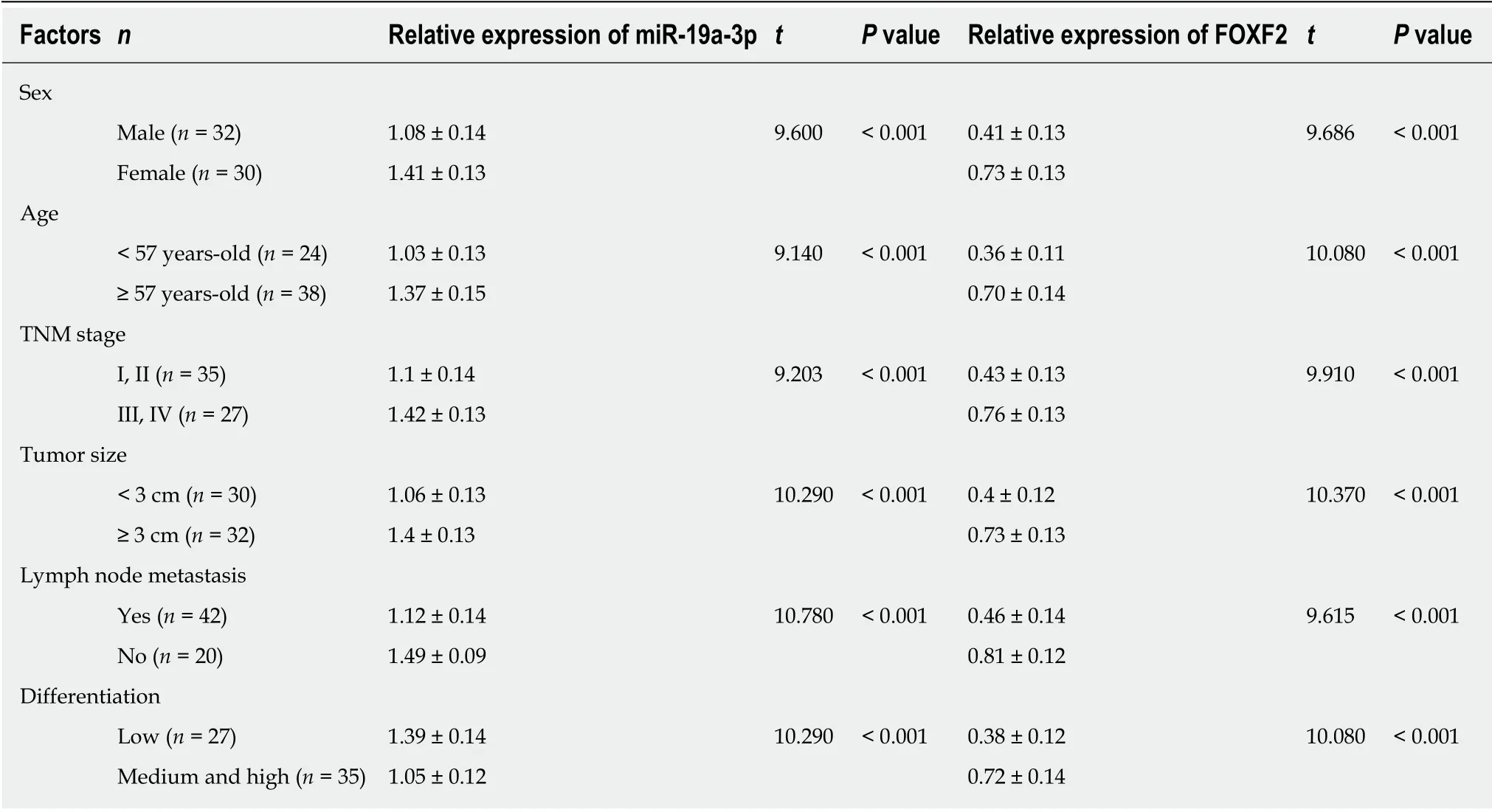
Table 1 Correlation of miR-19a-3p and Forkhead box F2 with pathological data of colorectal cancer patients
Identification of target genes of miR-19a-3p
In order to verify the relationship between miR-19a-3p and FOXF2, we predicated the downstream target genes of miR-19a-3p through Targetscan7.2, and found targeted binding sites between FOXF2 and miR-19a-3p. Therefore, we conducted dual luciferase detection, and found that inhibiting miR-19a-3p significantly lowered the luciferase activity of pmirGLO-FOXF2-3'UT Wt luciferase activity (P < 0.001), but had no effect on pmirGLO-FOXF2-3'UTR Mut. WB analysis revealed that HT29 and HCT116 cells transfected with miR-19a-3p-inhibitor had significantly upregulated FOXF2 expression, and those transfected with miR-19a-3p-mimics had significantly downregulated FOXF2 expression (both P < 0.05; Figure 4).
Rescue experiment
We transfected miR-19a-3p-mimics + sh-FOXF2 and miR-19a-3p-inhibitor + si-FOXF2 into HT29 and HCT116 cells, and detected their biological functions. There were no significant differences between these transfected cells and those transfected with miRNC regarding cell proliferation, invasion, and migration. Cells transfected with one of those constructs showed significantly stronger proliferation, invasion, and migration abilities than those transfected with miR-19a-3p-mimics, and showed significantly weaker proliferation, invasion, and migration abilities than those transfected with miR-19a-3p-inhibitor (all P < 0.05). The EMT-related protein assay revealed that there were no significant differences between cells transfected with miR-19a-3p-mimics +sh-FOXF2 or miR-19a-3p-inhibitor + si-FOXF2 and those transfected with miR-NC regarding the expression of E-cadherin, N-cadherin, and vimentin, while cells transfected with miR-19a-3p-mimics + sh-FOXF2 or miR-19a-3p-inhibitor + si-FOXF2 showed a significantly lower level of E-cadherin, and significantly higher levels of Ncadherin and vimentin than those transfected with miR-19a-3p-mimics, and showed a significantly higher level of E-cadherin, and significantly lower levels of N-cadherin and vimentin than those transfected with miR-19a-3p-inhibitor (all P < 0.05; Figure 5).
Inhibition of tumor growth in nude mice by silenced miR-19a-3p expression
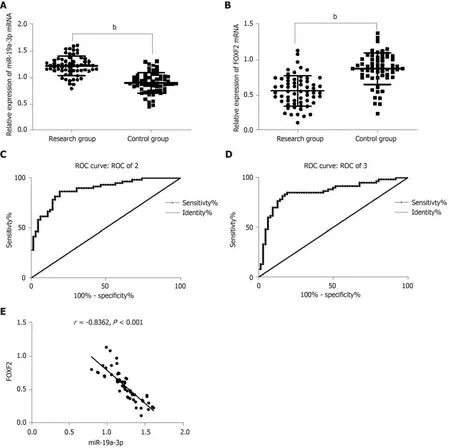
Figure 1 Expression of serum miR-19a-3p and Forkhead box F2 in colorectal cancer patients and their clinical value. A: The study group showed significantly higher miR-19a-3p expression than the control group, and miR-19a-3p was highly expressed in serum of colorectal cancer (CRC) patients. bP < 0.001; B: The study group showed significantly lower expression of Forkhead box F2 (FOXF2) than the control group, and FOXF2 had low expression in the CRC patients. bP < 0.001; C:The area under the curve of miR-19a-3p was 0.883; D: The area under the curve of FOXF2 was 0.850; E: The expression of serum miR-19a-3p was negatively related to that of FOXF2 in CRC patients (r = -0.836, bP < 0.001). CRC: Colorectal cancer; FOXF2: Forkhead box F2.
We constructed a tumor formation model by injecting HCT116 cells transfected with miR-19a-3p-inhibitor or miR-NC into the abdomen of nude mice. Nude mice injected with miR-19a-3p-inhibitor had a significantly slower growth rate than those injected with miR-NC, and after the mice were sacrificed, the tumor size and weight of mice with miR-19a-3p-inhibitor were significantly smaller than those with miR-NC (all P <0.001). In addition, mice infected with miR-19a-3p-inhibitor had significantly lower miR-19a-3p expression than those infected with NC (P < 0.001), which suggested that inhibiting miR-19a-3p expression can suppress tumor growth in vivo (Figure 6).
DISCUSSION
The biological characteristics of CRC cells affect the development of CRC. For example, the changes in proliferation, invasion, migration, and apoptosis of tumor cells affect and control tumor growth[17]. A large number of studies have concluded that cell proliferation is an important vital sign of biological organisms, and consistent proliferation, invasion and metastasis of tumor cells are the main causes of death for patients with malignant tumor[18]. Recent studies have indicated that miRNAs are strongly linked to various tumors, and play important roles in carcinogenesis and tumor progression, because they can affect the proliferation, invasion, migration, and apoptosis of tumor cells. One study confirmed that miR-19a-3p is abnormally and increasingly expressed in CRC tissues, and the changes in miR-19a-3p expression can affect the EMT of CRC cells[19], but the specific mechanism is not completely understood. This study determined that one biological function of miR-19a-3p in CRC cells is to regulate the FOXF2-mediated Wnt/β-catenin signaling pathway, thereby providing a novel theoretical basis for diagnosing and treating CRC in molecular biology.
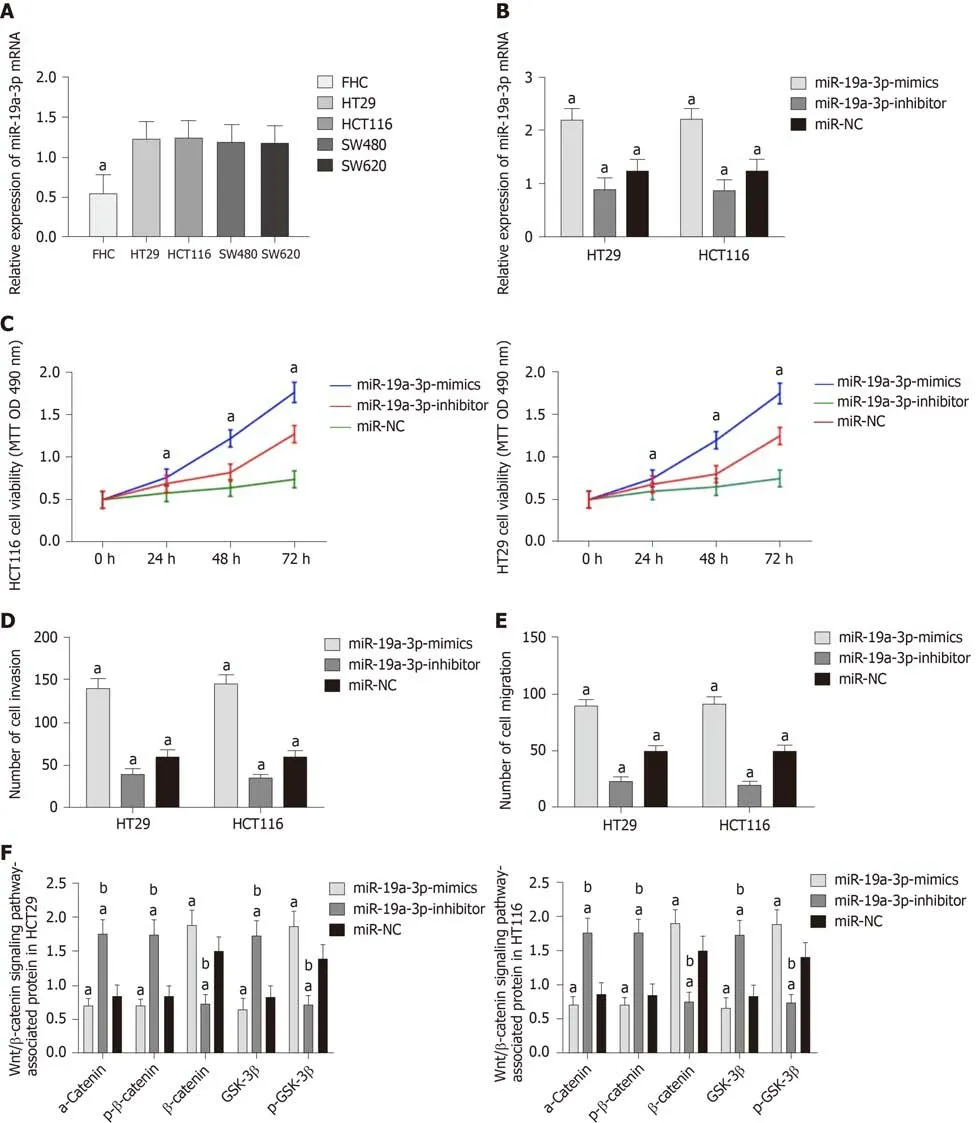
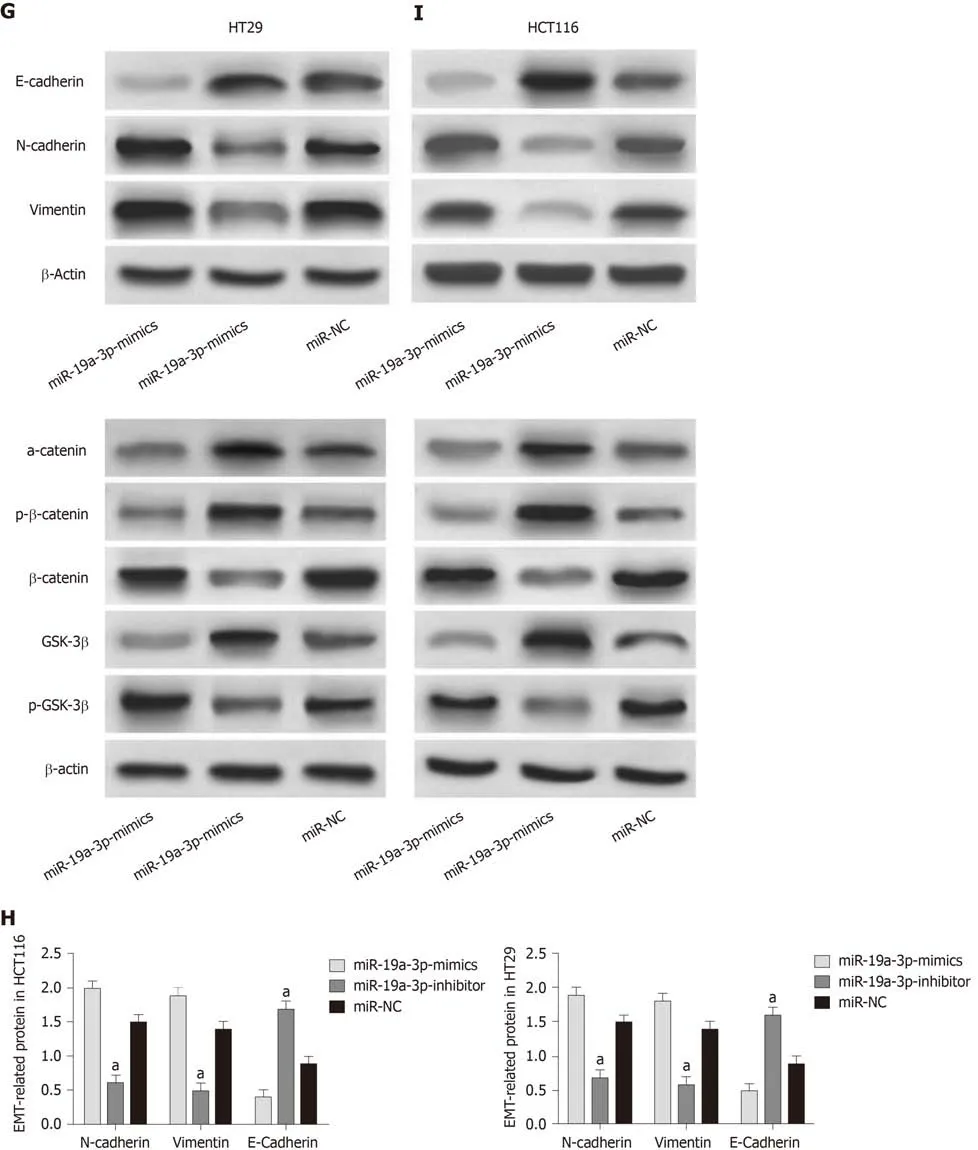
Figure 2 Effects of miR-19a-3p on the proliferation, invasion, migration, and epithelial-mesenchymal transition of colorectal cancer cells. A: miR-19a-3p was highly expressed in colorectal cancer cells; B: Compared with cells transfected with miR negative control (miR-NC), cells transfected with miR-19a-3p-mimics showed significantly upregulated expression of miR-19a-3p, and cells transfected with miR-19a-3p-inhibitor showed significantly downregulated expression of miR-19a-3p; C: Compared with cells transfected with miR-NC, cells transfected with miR-19a-3p-mimics showed significantly increased cell proliferation ability, and cells transfected with miR-19a-3p-inhibitor showed significantly decreased cell proliferation ability; D: Compared with cells transfected with miR-NC, cells transfected with miR-19a-3p-mimics showed significantly increased cell invasion ability, and cells transfected with miR-19a-3p-inhibitor showed significantly decreased cell invasion ability; E: Compared with cells transfected with miR-NC, cells transfected with miR-19a-3p-mimics showed significantly increased cell migration ability, and cells transfected with miR-19a-3p-inhibitor showed significantly decreased cell migration ability; F: Compared with cells transfected with miR-NC, cells transfected with miR-19a-3p-mimics showed significantly upregulated expression of p-GSK-3β and β-catenin, and significantly downregulated expression of GSK-3β and p-β-catenin, and cells transfected with miR-19a-3p-inhibitor showed significantly downregulated expression of p-GSK-3β and β-catenin, and significantly upregulated expression of GSK-3β and p-β-catenin; G: Western blot map; H: Compared with cells transfected with miR-NC, cells transfected with miR-19a-3p-mimics showed significantly upregulated expression of N-cadherin and vimentin, and significantly downregulated expression of E-cadherin, and cells transfected with miR-19a-3p-inhibitor showed significantly downregulated expression of N-cadherin and vimentin and significantly upregulated expression of E-cadherin; I: Western blot results. Note: a indicates that in comparison with cells transfected with miR-NC, bP < 0.001; b indicates that in comparison with cells transfected with miR-19a-3p-inhibitor, bP < 0.001. miR-NC:MiR negative control.
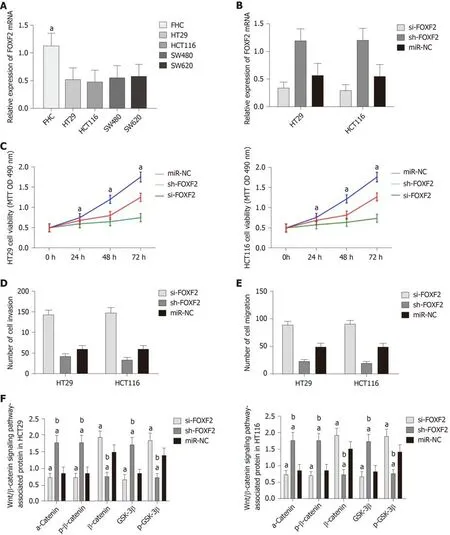
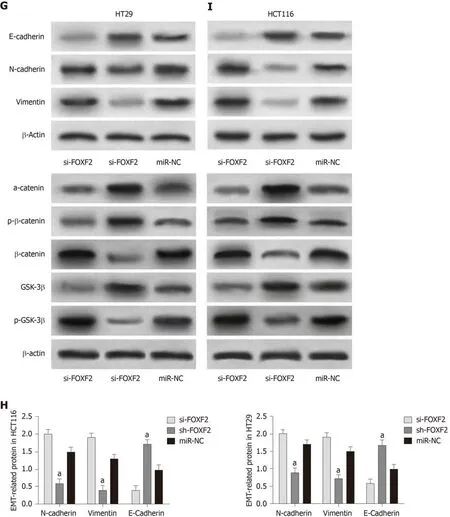
Figure 3 Effects of Forkhead box F2 on proliferation, invasion, migration and epithelial-mesenchymal transition of colorectal cancer cells. A: Forkhead box F2 (FOXF2) had low expression in colorectal cancer cells; B: Compared with cells transfected with MiR negative control (miR-NC), cells transfected with sh-FOXF2 showed significantly upregulated expression of FOXF2, and cells transfected with si-FOXF2 showed significantly down-regulated expression of FOXF2; C: Compared with cells transfected with miR-NC, cells transfected with si-FOXF2 showed significantly stronger cell proliferation ability, while cells transfected with sh-FOXF2 showed significantly weaker cell proliferation ability; D: Compared with cells transfected with miR-NC, cells transfected with si-FOXF2 showed significantly stronger cell invasion ability, while cells transfected with sh-FOXF2 showed significantly weaker cell invasion ability; E: Compared with cells transfected with miR-NC, cells transfected with si-FOXF2 showed significantly stronger cell invasion ability, while cells transfected with sh-FOXF2 showed significantly weaker cell invasion ability; F:Compared with cells transfected with miR-NC, cells transfected with si-FOXF2 showed significantly up-regulated expression of p-GSK-3β and β-catenin, and significantly down-regulated expression of GSK-3β and p-β-catenin, and cells transfected with Sh-FOXF2 showed significantly downregulated expression of p-GSK-3β and β-catenin, and significantly upregulated expression of GSK-3β and p-β-catenin; G: Western Blot results; H: Compared with cells transfected with miR-NC, cells transfected with si-FOXF2 showed significantly up-regulated expression of N-cadherin and vimentin, and significantly down-regulated expression of E-cadherin, and cells transfected with sh-FOXF2 showed significantly down-regulated expression of N-cadherin and vimentin and significantly up-regulated expression of E-cadherin; I:Western blot results. Note: a indicates that in comparison with cells transfected with miR-NC, bP < 0.001; b indicates that in comparison with cells transfected with sh-FOXF2, bP < 0.001. FOXF2: Forkhead box F2; miR-NC: MiR negative control.
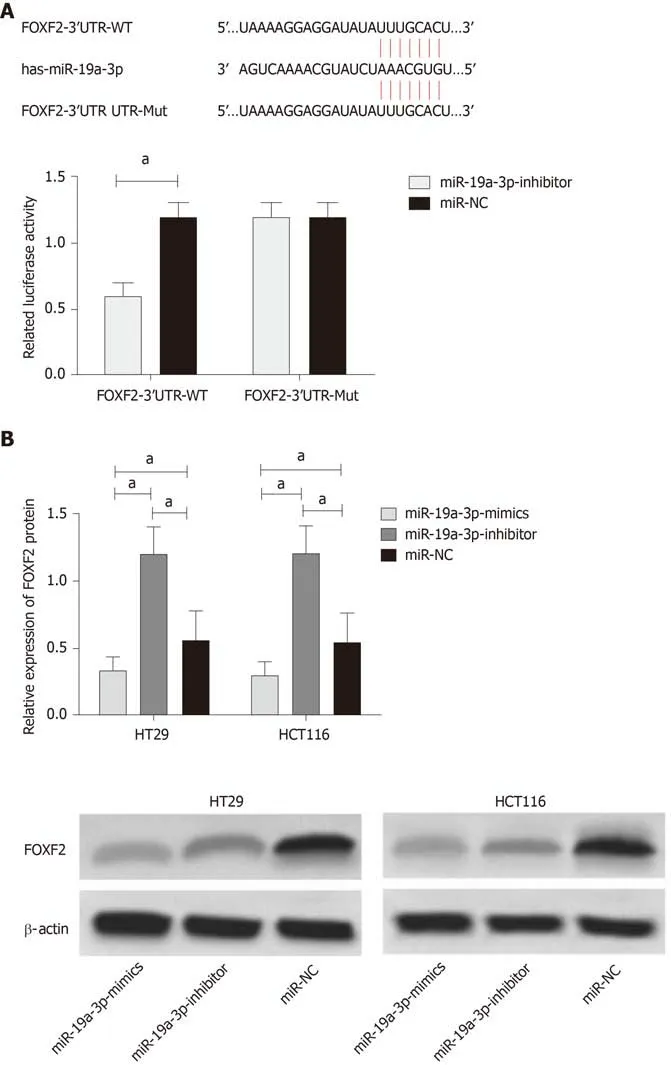
Figure 4 Luciferase detection. A: Binding sites between miR-19a-3p and Forkhead box F2, and relative luciferase activity - dual luciferase reporter assay; B: Expression of Forkhead box F2 in HT29 and HCT116 cells after transfection. aP < 0.001.
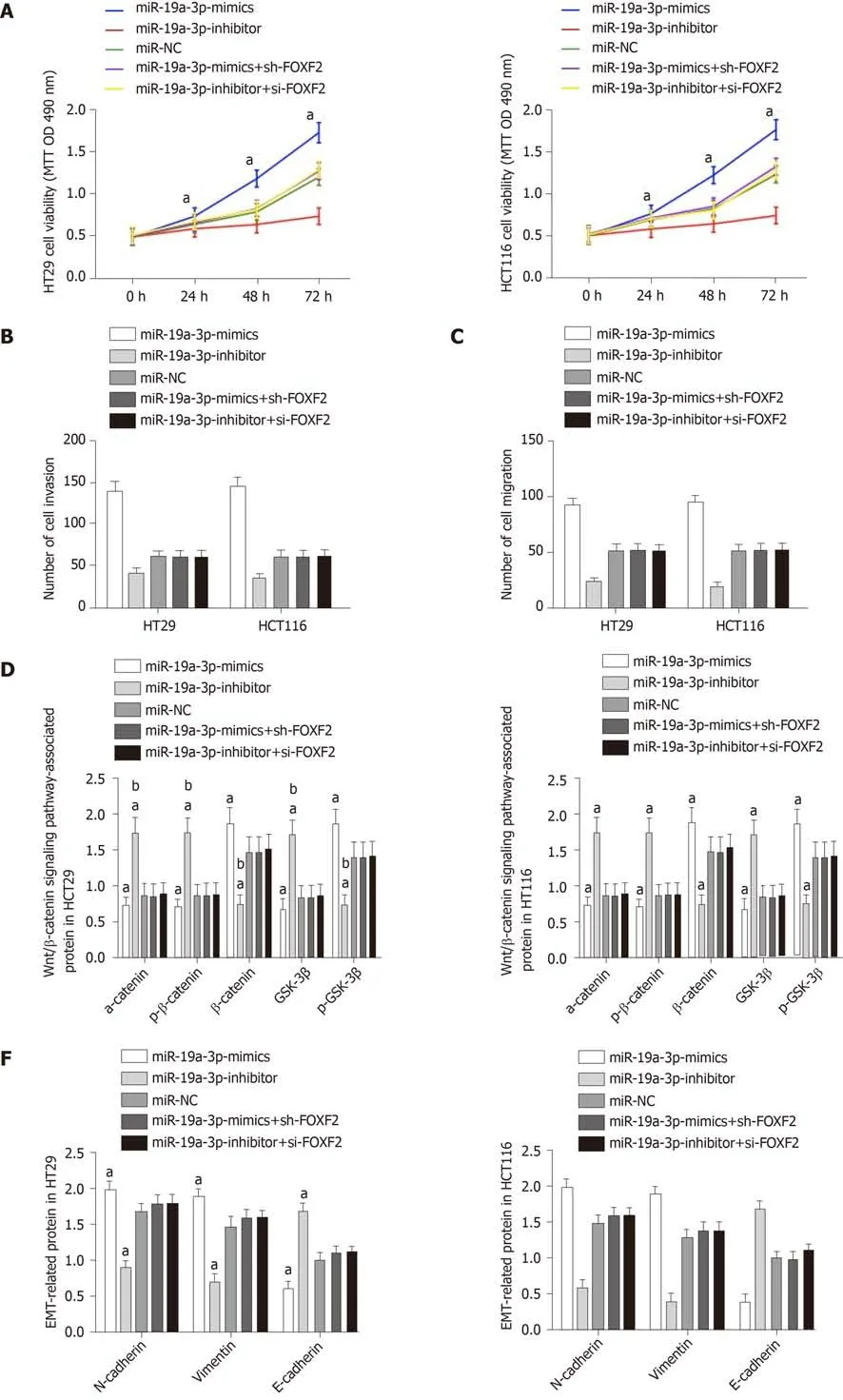
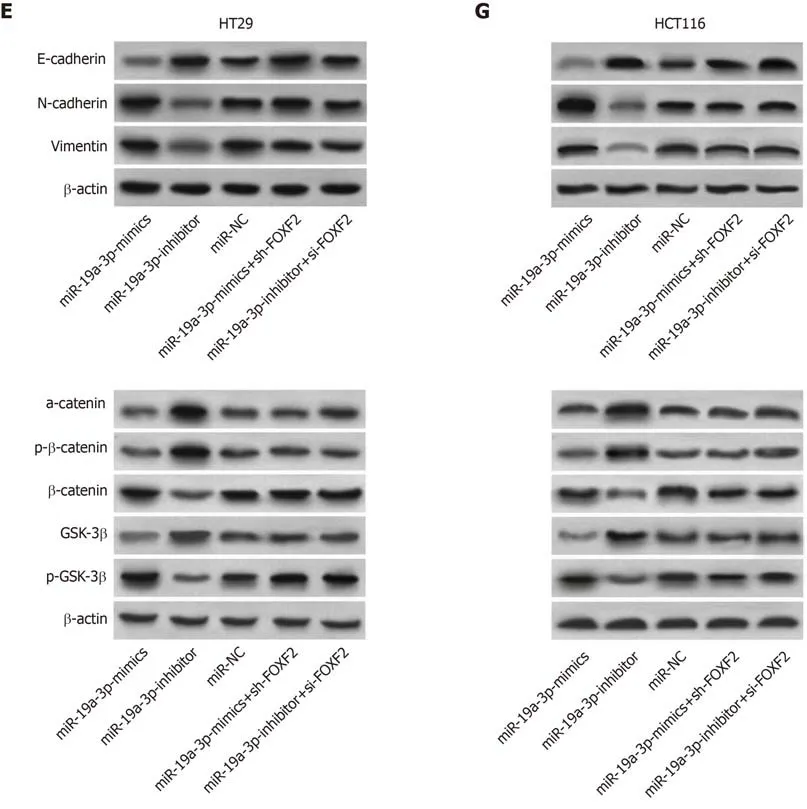
Figure 5 Rescue experiment. A: Cell proliferation ability after transfection with miR-19a-3p-mimics + sh-Forkhead box F2 (FOXF2) or miR-19a-3p-inhibitor + si-FOXF2; B: Cell invasion ability after transfection with miR-19a-3p-mimics + sh-FOXF2 or miR-19a-3p-inhibitor + si-FOXF2; C: Cell migration ability after transfection with miR-19a-3p-mimics + sh-FOXF2 or miR-19a-3p-inhibitor + si-FOXF2; D: Expression of p-GSK-3β, β-catenin, α-catenin,, GSK-3β and p-β-catenin in cells transfected with miR-19a-3p-mimics + sh-FOXF2 or miR-19a-3p-inhibitor + si-FOXF2; E: Western blot map; F: Expression of E-cadherin, N-cadherin, and vimentin in cells transfected with miR-19a-3p-mimics+sh-FOXF2 or miR-19a-3p-inhibitor+si-FOXF2; G: Western blot map; aP < 0.05 vs cells transfected with miR- negative control or miR-19a-3p-mimics + sh-FOXF2 or with cells transfected with miR-19a-3p-inhibitor+si-FOXF2, bP < 0.001 vs cells transfected with miR-19a-3p-inhibitor.FOXF2: Forkhead box F2.
In this study, we conducted qPCR to evaluate the expression of serum miR-19a-3p in CRC patients and healthy individuals, and found that serum miR-19a-3p was abnormally increased in CRC patients, in line with a study conducted by Jiang et al[20].We also carried out relevant analyses on the clinicopathological features of the patients, finding that high miR-19a-3p expression was correlated with age, sex, tumor size, differentiation, TNM stage, and LNM of CRC patients. We drew a ROC curve for miR-19a-3p, and found that its AUC was more than 0.8, suggesting that miR-19a-3p had a high diagnostic value for CRC. Those findings indicated that miR-19a-3p is correlated with the development and progression of CRC. A large number of studies have confirmed that miRNA can affect the biological functions of cells by regulating their target genes. For example, miR-19a-3p is reportedly able to target downstream gene pairs to regulate the biological functions of cells[21,22]. We carried out our analyses based on the TargetScan database, finding that there was a targeted relationship between miR-19a-3p and FOXF2. We also analyzed the relationship between FOXF2 and clinicopathological characteristics of the patients, finding that low expression of FOXF2 was correlated with clinicopathologic feature of CRC patients. Furthermore,we drew a ROC curve for FOXF2, and found that its AUC was more than 0.8. FOXF2,as a transcription factor, regulates the progression and differentiation of tumors, and is involved in the changes of biological functions of various tumor cells[23]. It can affect the cycle and other biological functions of CRC cells by regulating CRC cell genes[24].In this study, we analyzed FOXF2 expression in CRC cells based on the data from TCGA, and found that CRC cells showed increased miR-19a-3p and decreased FOXF2 expression, which was consistent with the experimental results of this study,suggesting that both may play regulatory roles in CRC. However, it remains unclear whether there is a regulatory relationship between the two and their effects on the EMT of CRC cells.
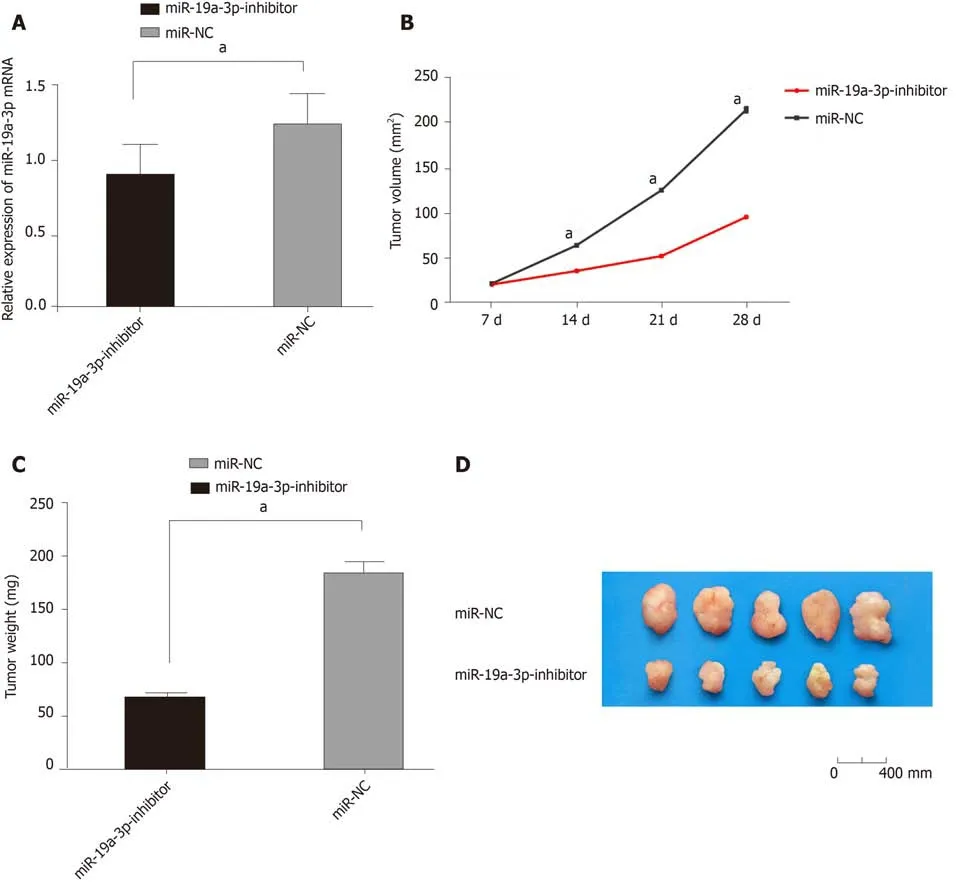
Figure 6 Effects of inhibiting miR-19a-3p expression on tumor growth in nude mice. A: Comparison between the two groups revealed that tumor tissues with miR-19a-3p-inhibitor showed lower expression of miR-19a-3p; B: Comparison between the two groups revealed that the mice with miR-19a-3p-inhibitor showed slower tumor growth rate; C: Comparison between the two groups revealed that the mice with miR-19a-3p-inhibitor showed lighter tumor weight; D: Comparison between the two groups revealed that the mice with miR-19a-3p-inhibitor showed smaller tumor size. aP < 0.001.
In order to verify these results, we performed additional cell experiments. We compared human CRC cell lines (HT29, SW480, SW620, and HCT116) with the human normal colonic epithelial cell line (FHC), and found that CRC cells showed higher miR-19a-3p expression and lower FOXF2 expression, consistent with our previous results. After the comparison, we regulated the expression of miR-19a-3p and FOXF2 in HT29 and HCT116 cells, and transfected miR-19a-3p-mimics, miR-19a-3p-inhibitor,sh-FOXF2, and si-FOXF2 into HT29 and HCT116 cells, finding that cells with inhibited miR-19a-3p expression or overexpressed FOXF2 expression showed significantly suppressed cell proliferation and invasion abilities, and also showed decreased expression of N-cadherin and vimentin, and significantly increased E-cadherin expression during EMT. However, HT29 and HCT116 transfected with miR-19a-3pmimics and FOXF2-inhibitor showed opposite results. The occurrence and development of tumors are closely related to the EMT, and can promote proliferation,invasion, and migration of tumor cells. Therefore, the above results indicate that miR-19a-3p acts as a potential target for CRC, and silencing of miR-19a-3p can suppress the proliferation, invasion, migration, and EMT of CRC cells. Tumorigenicity in nude mice in vitro revealed that miR-19a-3p overexpression was able to strongly promote tumor formation and growth, which further indicated the function of miR-19a-3p changes in CRC. However, it is not clear how miR-19a-3p affects the biological functions of CRC cells and EMT.
One study demonstrated that activating the Wnt/β-catenin signaling pathway exerts great effects on promoting EMT[25], and some studies have shown that, by regulating the Wnt/β-catenin signaling pathway, FOXF2 can inhibit the proliferation,migration, invasion, and EMT of cervical cancer Hela cells[26]. In this study, we observed the Wnt/β-catenin signaling pathway-related proteins after inhibiting or overexpressing miR-19a-3 in CRC cells, finding that cells with underexpressed miR-19a-3p or overexpressed FOXF2 showed significantly downregulated levels of p-GSK-3β, β-catenin, N-cadherin, and vimentin, and significantly upregulated levels of GSK-3β, p-β-catenin, α-catenin, and E-cadherin. However, cells with overexpressed miR-19a-3p or underexpressed FOXF2 presented with the opposite results. It suggests that miR-19a-3p is able to suppress the activation of the Wnt/β-catenin signaling pathway by regulating FOXF2, thereby inhibiting the EMT of cells. Studies on the Wnt signaling pathway have also revealed that activating the Wnt/β-catenin signaling pathway can promote the invasion and proliferation of esophageal squamous cell carcinoma, nasopharyngeal carcinoma, as well as breast cancer cells, and induce their EMT[27-29].
At the end of the study, we performed a rescue experiment, and found no significant differences between HT29 and HCT116 cells transfected with miR-19a-3pmimics + sh-FOXF2 and miR-19a-3p-inhibitor + si-FOXF2 and those transfected with miR-NC regarding cell proliferation, invasion and migration, but cells transfected with miR-19a-3p-mimics + sh-FOXF2 and miR-19a-3p-inhibitor + si-FOXF2 showed significantly stronger abilities than those transfected with miR-19a-3p-inhibitor regarding cell proliferation, invasion and migration, and significantly weaker cell proliferation, invasion and migration abilities than those transfected with miR-19a-3pmimics. The results suggested that miR-19a-3p can target FOXF2. Therefore, the dual luciferase reporter assay was conducted to verify the correlation of miR-19a-3 with FOXF2. The results showed that overexpressing miR-19a-3 significantly increased the luciferase activity of FOXF2-3'UT Wt, but had no effect on the activity of FOXF2-3'UTR Mut. Moreover, cells transfected with miR-19a-3p-inhibitor showed significantly upregulated FOXF2 expression, which indicated that there was a targeted regulatory relationship between miR-19a-3p and FOXF2. Therefore, we believe that inhibiting the expression of miR-19a-3p can affect the biological functions of CRC cells by promoting the expression of FOXF2.
This study proved that with high expression in CRC cells, miR-19a-3p can inhibit FOXF2-mediated cell proliferation. However, this study had some limitations. For example, other factors that may affect the growth of tumors in mice were not been further analyzed. The regulatory network of miR-19a-3p is still under investigation,and whether it can affect the development and progression of tumors through other mechanisms requires more research. Therefore, bioinformatics analysis is still ongoing to explore the regulatory network, in order to provide more references for our experiments. To summarize, miR-19a-3p can mediate changes of CRC cells by regulating the FOXF2 expression, and it is of great significance to study molecular targeted treatments for CRC.
ARTICLE HIGHLIGHTS
Research background
Colorectal cancer (CRC) has a high rate of mortality, and patients with this disease often miss the optimal treatment period due to the lack of clinical symptoms of early CRC, which affects their prognosis. At present, CRC is extremely difficult to prevent and treat.
Research motivation
In this study, we studied the effects of regulating the Forkhead box F2 (FOXF2)-mediated Wnt/β-catenin signaling pathway by miR-19a-3p on the biological functions of CRC cells from the perspective of the mechanism of CRC, so as to explore the changes in biological functions of CRC cells.
Research objectives
This study evaluated the expression of miR-19a-3p and FOXF2 in patients with CRC and the relevant mechanisms.
Research methods
Elbow venous blood was sampled from CRC patients and healthy individuals, and blood serum was saved for later analysis. MiR-19a-3p-mimics, miR-19a-3p-inhibitor, miR-NC, si-FOXF2, and sh-FOXF2 were transfected into HT29 and HCT116 cells. Then quantitative polymerase chain reaction was applied to determine the expression of miR-19a-3p and FOXF2 in HT29 and HCT116 cells, and Western blotting was conducted to determine the expression of FOXF2, GSK-3β, p-GSK-3β, β-catenin, p-β-catenin, α-catenin, N-cadherin, E-Cadherin, and vimentin. The MTT, Transwell, and wound-healing assays were applied to detect cell proliferation, invasion,and apoptosis, respectively, and the dual luciferase reporter assay was used to determine the relationship between miR-19a-3p and FOXF2.
Research results
MiR-19a-3p was highly expressed in the serum of the patients, while FOXF2 was lowly expressed in them. MiR-19a-3p and FOXF2 were related to age, sex, tumor size, tumor, node,metastasis staging, lymph node metastasis, and differentiation of patients with CRC. Silencing of miR-19a-3p and over-expression of FOXF2 suppressed epithelial-mesenchymal transition,proliferation, invasion, and migration of cells, and Western blot assay supported that silencing of miR-19a-3p and over-expression of FOXF2 significantly suppressed epithelial-mesenchymal transition. Dual luciferase reporter assay confirmed that there was a targeted relationship between miR-19a-3p and FOXF2. Therefore, inhibiting the expression of miR-19a-3p can affect the biological functions of CRC cells by promoting the expression of FOXF2.
Research conclusions
Inhibiting the expression of miR-19a-3p can affect the biological functions of CRC cells by promoting the expression of FOXF2.
Research perspectives
It has been confirmed that inhibiting the expression of miR-19a-3p can up-regulate the FOXF2-mediated Wnt/β-catenin signaling pathway, thus affecting the epithelial-mesenchymal transition, proliferation, invasion, and migration of cells, so miR-19a-3p is expected to be a potential therapeutic target for CRC.
杂志排行

World Journal of Gastroenterology的其它文章
- Flushing as atypical initial presentation of functional gallbladder neuroendocrine carcinoma: A case report
- Expression of miR-1304 in patients with esophageal carcinoma and risk factors for recurrence
- Validation and evaluation of clinical prediction systems for first and repeated transarterial chemoembolization in unresectable hepatocellular carcinoma: A Chinese multicenter retrospective study
- Bacterial infection triggers and complicates acute-on-chronic liver failure in patients with hepatitis B virus-decompensated cirrhosis: A retrospective cohort study
- Characterizing the composition of intestinal microflora by 16S rRNA gene sequencing
- Histone deacetylase inhibitor pre-treatment enhances the efficacy of DNA-interacting chemotherapeutic drugs in gastric cancer