乐清湾大型底栖动物群落及其与环境因子之间的关系
2020-03-12王航俊姚炜民林义邹清刘亚林
王航俊,姚炜民*,林义,邹清,刘亚林
( 1. 国家海洋局温州海洋环境监测中心站,浙江 温州 325000)
1 引言
大型底栖动物是海洋环境中的一个重要生态类群,在食物网中扮演着消费者和转移者的角色,在海洋生态系统的物质循环和能量流动中发挥着重要作用[1]。大多数大型底栖动物具有生活相对稳定、活动能力弱、活动范围较小,对海洋环境污染反应敏感等特点。因此,对大型底栖动物群落的生态特征研究不仅能了解底栖动物群落的动态变化,而且为评估海洋生态系统健康状况,合理利用大型底栖动物资源提供理论依据。
乐清湾位于浙江省南部沿岸,瓯江口北侧,自1977 年漩门港一期工程建坝堵截漩门口之后,成为一个半封闭型海湾。随着乐清湾沿岸县市经济的高速发展,乐清湾的海洋经济也得到了全面发展,包括养殖业、港口运输业、临港工业、海洋渔业以及滨海旅游业等。与此同时,大规模的填海造地、港口建设等人类活动,使岸线和滩涂宽度发生了较大的变化;湾两侧的工业废水、生活污水、养殖废水等的大量排放,导致水体中的富营养化程度也呈上升趋势,乐清湾的生态环境正面临着严峻的考验。渔业资源的逐渐萎缩[2],养殖死亡事件的频频出现[3],都说明乐清湾的生态环境已发生了较大的变化。而关于该海域的底栖生物生态研究相对匮乏,其中胡颢琰等[4]、贾海波等[5]等对浙江全省和浙江南部近岸海域的底栖生物研究中有所涉及,但是调查站位和调查季节不够充分;廖一波等[6]在乐清湾西门岛附近的海域开展过4 个季度底栖动物功能群研究,调查区域十分有限。戴泽蘅[7]在1981 年,杨俊毅等[8]在2002-2003 年以及王航俊等[9]在2015 年较为系统的开展了乐清湾底栖生物调查。但是这3 次调查主要集中在底栖生物组成方面的研究,对乐清湾大型底栖生物群落结构特征以及群落与环境因子间关系的研究还未见公开报道。本研究通过分析乐清湾大型底栖动物群落结构及其与环境因子的相关性,综合评定调查海域的生态状况,以期为乐清湾海域生态系统深入研究提供参考。
2 材料与方法
2.1 站位布设及采样方法
2015 年1 月20 日、3 月7-13 日、5 月3-7 日、8 月4-6 日和10 月12-15 日共开展了5 个航次的调查取样,其中1 月共布设了17 个站位,3 月、5 月、8 月和10 月布设了21 个站位。因乐清湾整体呈现葫芦状,可将其分为内湾、中湾和外湾,其中茅埏岛中间线以北海域为内湾,鸡蛋山与胜利塘北片的连线与茅埏岛中间线之间的海域为中湾,其余为外湾。站位均匀布设在内、中、外湾,基本覆盖了整个乐清湾水域。调查站位见图1。大型底栖动物取样采用面积为0.05 m2的抓斗式采泥器重复取样两次,合并为一个样品,具体操作方法按《海洋监测规范》(GB 17378.7–2007)进行。底层水体的温度(Temp)、pH、盐度(Sal)、溶解氧(DO)、化学需氧量(COD)、悬浮物(SS)、叶绿素a(Chl a)、磷酸盐(SRP)、亚硝酸盐–氮(NO2-N)、硝酸盐−氮(NO3-N)、氨氮(NH4-N)和油类(Oil)等环境因子采样和分析方法按《海洋监测规范》(GB 17378.4–2007)和(GB 17378.7–2007)进行;沉积物采样和粒度分析按《海洋调查规范》(GB/T 12763.8–2007)进行。
2.2 数据分析与处理
2.2.1 群落聚类分析
利用PRIMER6.0 软件进行群落结构的分析。去除在整个研究海域相对密度小于1%的物种,但保留在任何一个站位相对密度大于3%的物种,以减少稀有种对群落划分的干扰。为平衡优势种和稀有种在群落中的作用,对种类丰度数据进行4 次方根转换,得到Bray-Curtis 相似性矩阵,利用CLUSTER 进行聚类分析,ANOSIM 来检验各聚类分组之间的差异显著性,通过SIMPER 分析获得表征不同群落或造成群落差异的特征种。
2.2.2 群落稳定性分析
利用PRIMER6.0 软件绘制丰度/生物量曲线(ABC曲线),用以表征大型底栖动物群落稳定性水平。如果生物量曲线始终位于丰度曲线之上,则表征群落未受到扰动较为稳定;反之,则表征群落受到严重扰动不稳定;当两曲线有交叉或重叠,则表征群落受到中度扰动而不够稳定。
2.2.3 群落结构与环境因子的相关性关系
将原始环境因子数据进行标准化操作后计算欧式距离(Euclidean)获得一个不相似矩阵,种类丰度数据进行4 次方根转换,得到Bray-Curtis 相似性矩阵,通过BVSTEP 分析选择能够解释群落结构的最佳环境变量组合[10]。

图 1 乐清湾大型底栖动物取样站位Fig. 1 Sampling stations of macrobenthos in the Yueqing Bay
2.2.4 群落特征种与环境因子的关系
利用CANOCO5.0 软件对群落结构聚类分析获得的群落特征种与环境因子的典范对应分析(Canonical Correspondence Analysis, CCA),利用Monte Carlo 检验CCA 排序中所有轴的显著性。
3 结果
3.1 海域环境状况
乐清湾海域环境参数的月份变化如表1 所示,其中水温、pH、盐度、溶解氧、叶绿素a、磷酸盐和硝酸盐−氮的月度之间存在极显著差异P<0.01,悬浮物和油类的月度之间存在显著差异P<0.05,水深、化学耗氧量、亚硝酸盐−氮和氨−氮的月度之间不存在显著性差异。
3.2 群落聚类分析
对乐清湾5 个航次的大型底栖动物进行聚类分析,结果见图2。1 月份在相似度19%的水平上,可以将乐清湾大型底栖动物划分为5 个群落,ANOSIM 检验分析表明,各群落间的差异显著(Global R=0.804,P<0.01);3 月份在相似度24%的水平上,可以将乐清湾大型底栖动物划分为7 个群落,ANOSIM 检验分析表明,各群落间的差异显著(Global R =0.768,P<0.01);5 月份在相似度23%的水平上,可以将乐清湾大型底栖动物划分为5 个群落,ANOSIM 检验分析表明,各群落间的差异显著(Global R =0.831,P<0.01);8 月份在相似度13%的水平上,可以将乐清湾大型底栖动物划分为7 个群落,ANOSIM 检验分析表明,各群落间的差异显著(Global R =0.682,P<0.01);10 月份在相似度15%的水平上,可以将乐清湾大型底栖动物划分为8 个群落,ANOSIM 检验分析表明,各群落间的差异显著(Global R =0.916,P<0.01)。
3.3 群落空间分布
根据聚类分析结果获得各群落的空间分布及其主要特征种如图3 所示,仅包含一个站位的群落由于密度和生物量值极低,故不进行SIMPER 特征种分析。
1 月份的优势群落为群落Ⅳ,该群落占据了35%的站位,主要特征种为薄云母蛤(Yoldia similis),分布在中湾和外湾。其他群落组别还有群落Ⅱ、群落Ⅲ和群落Ⅴ,群落Ⅱ的主要特征种为寡鳃齿吻沙蚕(Nephtys oligobranchia)和索沙蚕属一种(Lumbrinerissp.),分布在内湾和中湾;群落Ⅲ的主要特征种为寡鳃齿吻沙蚕,在整个乐清湾均有分布;群落Ⅴ的主要特征种为中蚓虫属一种(Mediomastus sp.)和沙沟虾属一种(Byblis sp.),在整个乐清湾均有分布。

表 1 乐清湾海域环境参数(平均值±标准差)的月份变化Table 1 Monthly change of environmental parameters (mean±standard deviation) in the Yueqing Bay
3 月份的优势群落为群落Ⅶ,该群落占据了57%的站位,主要特征种为薄云母蛤、寡鳃齿吻沙蚕、不倒翁虫(Sternaspis sculata)和中蚓虫属一种,在整个乐清湾均有分布。其他群落组别还有群落Ⅳ和群落Ⅵ,群落Ⅳ的主要特征种为光滑河篮蛤(Potamocorbula laevis)、冠奇异稚齿虫(Paraprionospio cristata)和全刺沙蚕属一种(Nectoneanthes sp.),仅在内湾有分布;群落Ⅵ的主要特征种为寡鳃齿吻沙蚕和钩虾一种(Gammaridae),仅在内湾有分布。
5 月份的优势群落为群落Ⅴ,该群落占据了62%的站位,主要特征种为薄云母蛤和不倒翁虫,在整个乐清湾均有分布。其他群落组别还有群落Ⅲ和群落Ⅳ,群落Ⅲ的主要特征种为寡鳃齿吻沙蚕,在内湾和中湾有分布;群落Ⅳ的主要特征种为小头虫(Capitella capitata)和寡鳃齿吻沙蚕,在内湾和中湾有分布。
8 月份的优势群落为群落Ⅵ和群落Ⅴ,分别占据了33%和29%的站位,群落Ⅵ的主要特征种为不倒翁虫、异毛虫一种(Paraonidae)和多丝独毛虫(Tharyx multifilis),在整个乐清湾均有分布;群落Ⅴ的主要特征种为双形拟单指虫(Cossurella dimorpha),在内湾和中湾有分布。其他群落组别还有群落Ⅲ和群落Ⅳ,群落Ⅲ的主要特征种为双唇索沙蚕(Lumbrineris cruzensis),在内湾和外湾有分布;群落Ⅳ的主要特征种为纽虫一种(Nemertea)和背毛背蚓虫(Notomastus cf.aberans),在内湾和外湾有分布。

图 2 乐清湾大型底栖动物群落结构聚类分析Fig. 2 Cluster analysis of macrobenthic community structure in the Yueqing Bay
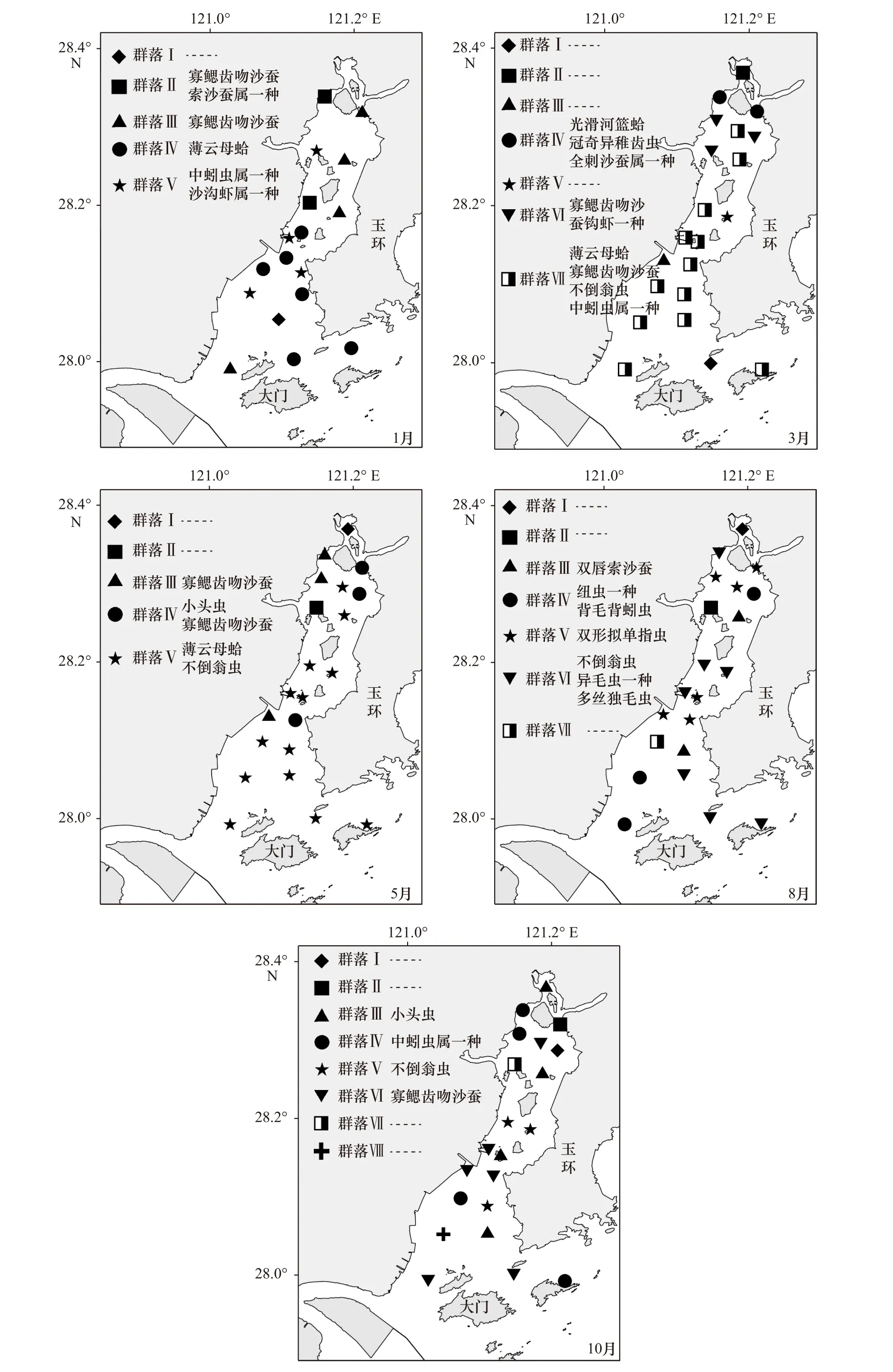
图 3 乐清湾大型底栖动物群落空间分布Fig. 3 Spatial distribution of macrobenthic community in the Yueqing Bay
10 月份的优势群落为群落Ⅵ、群落Ⅲ和群落Ⅳ,分别占据了29%、19%和19%的站位,群落Ⅵ的主要特征种为寡鳃齿吻沙蚕,在整个乐清湾均有分布;群落Ⅲ的主要特征种为小头虫,在整个乐清湾均有分布;群落Ⅳ的主要特征种为中蚓虫属一种,在内湾和外湾有分布。其他群落组别还有群落Ⅴ,主要特征种为不倒翁虫,在中湾和外湾有分布。
3.4 群落稳定性分析
利用ABC 曲线分别对5 个月份全部站位以及各月份不同群落组别进行稳定性分析,其中仅包含一个站位的群落因物种数极少故不进行ABC 曲线的稳定性分析[11]。如图4,1 月份群落Ⅳ和群落Ⅴ处于严重扰动的不稳定状态;3 月份群落Ⅶ处于中度扰动的不稳定状态;5 月份全站位和各群落均处于较稳定状态;8 月份全站位、群落Ⅲ、群落Ⅳ、群落Ⅴ和群落Ⅵ均有不同程度的扰动而处于不稳定状态;10 月份群落Ⅲ和群落Ⅵ处于中度扰动的不稳定状态。
3.5 群落结构与环境因子之间的关系
通过BVSTEP 分析年度和各月份的大型底栖动物种类丰度与环境因子的关系,结果显示群落结构与多个环境因子组合相关性比单个环境因子的相关性高(表2)。年度、3 月和5 月的最佳环境因子组合与群落结构存在显著性相关(P<0.05),1 月、8 月和10 月的最佳环境因子组合与群落结构不存在显著性相关(P>0.05)。不同月份及年度之间的最佳环境因子组合存在较大的差异,出现次数最多的环境因子为氨氮和磷酸盐。
3.6 群落特征种与环境因子的关系
乐清湾各月份的大型底栖动物群落结构特征种与13 个共有环境因子的CCA 分析结果见表3。前两个排序轴的特征值分别为0.426 和0.315,特征种和环境因子前两个排序轴的相关性系数为0.885 和0.810,Monte Carlo 显著性检验结果表明,第一排序轴和所有轴均呈极显著性差异(第1 轴:F=4.9,P=0.002;所有轴:F=2.1,P=0.002),说明排序轴能较好地反映出特征种和环境因子之间的关系。单个环境因子对乐清湾大型底栖群落结构特征种的解释率和显著性检验结果见表4,其中水温、盐度、溶解氧、硝酸盐−氮、亚硝酸盐−氮、pH 与群落结构特征种之间均存在极显著性相关(P<0.01)。

图 4 乐清湾大型底栖动物群落ABC 曲线Fig. 4 ABC curves of macrobenthic community in the Yueqing Bay
由图5 可见,在沿轴1 水平方向上,位于右边的群落特征种,特别是冠奇异稚齿虫、全刺沙蚕属一种、腺袋才女虫与含氮营养盐呈正相关,而与盐度呈负相关,这些种类主要分布在近岸和河口有淡水注入的地方。
在沿轴2 垂直分布上,位于下方的群落特征种,特别是后指虫、日本细莱毛虫、沙钩虾属一种、蛇尾幼体与溶解氧、pH 等呈正相关,这些种类需要较高的溶解氧含量,而位于轴2 上方的群落特征种如中华后指虫、异蚓虫和双唇索沙蚕等则更加适应溶解氧含量较低的环境。
位于第二象限内的群落特征种,特别是纽虫一种、双唇索沙蚕、异毛虫一种与水温、盐度和叶绿素a呈正相关,与硝酸盐−氮呈负相关,这些种类主要集中在8 月份水温和盐度较高的时候出现。
在排序图原点附近分布的种类主要受各种环境因子的综合作用而非个别因子的作用,这些种类包括寡鳃齿吻沙蚕、中蚓虫属一种、不倒翁虫、多丝独毛虫、索沙蚕属一种。由于各种环境因子对这些种类的生态分布都有或多或少以及直接间接的影响,因此它们的生态位较大,能适应多种不同的环境。这些种类在多数月份和站位都有出现,是乐清湾大型底栖动物的优势种和常见种。
4 讨论
4.1 乐清湾大型底?栖动物群落特征
基于Bray-Curtis 相似性矩阵的等级聚类和SIMPER 特征种分析显示,1 月、3 月和5 月的优势群落明显,优势群落特征种组成较为一致,主要为薄云母蛤和不倒翁虫,其中3 月和5 月的优势群落的站位空间分布也较为一致,85%以上站位均为相同站位,覆盖了外湾和中湾大部分站位以及少量内湾站位。该优势群落的结构特征与乐清湾南面的洞头列岛附近海域[12]和乐清湾北面的象山港海域[13]的群落结构特征较为一致。8 月和10 月的优势群落不明显,群落特征种差异大。

表 2 乐清湾大型底栖动物群落结构与环境因子的BVSTEP 相关性分析Table 2 BVSTEP analysis of community structure and environmental factors of macrobenthos in the Yueqing Bay

表 3 乐清湾大型底栖动物群落结构特征种与环境因子之间CCA 分析结果Table 3 CCA results of structural characteristics and environmental factors of macrobenthic community in the Yueqing Bay
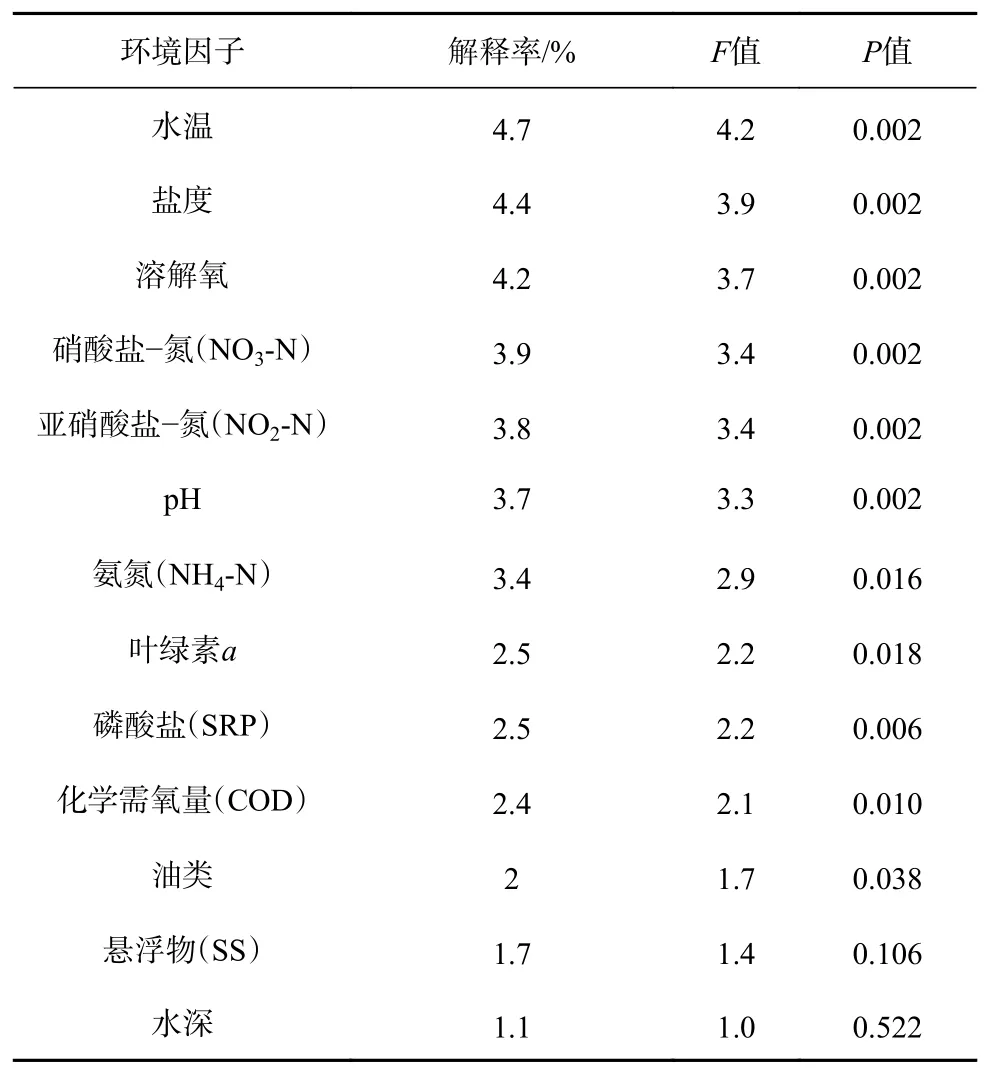
表 4 环境因子的解释率和显著性检验结果Table 4 Interpretation rate of environmental factors and significance test results
SIMPER 特征种分析显示除薄云母蛤在3 个月份中均有出现外,还有多个特征种在多个月份中出现,如寡鳃齿吻沙蚕在1 月、3 月、5 月和10 月均有出现;不倒翁虫在3 月、5 月、8 月和10 月均有出现;中蚓虫属一种在1 月、3 月和10 月均有出现。各月份之间的群落特征种相似性较高,表明乐清湾各月份之间的群落结构差异不明显,相类似的情况在烟台近海[14]、象山港[15]、杭州湾[16]等近岸海域也有出现。
5 个月份共计18 个群落的SIMPER 特征种分析,累积出现28 种群落特征种,其中多毛类占绝对优势,有23 种占比82.1%,其他类群还有软体动物2 种,占比7%,棘皮动物、甲壳类和纽形动物各1 个种,分别占比3.6%。这表明多毛类已经成为乐清湾大型底栖动物群落结构的优势类群,类似的群落结构特点在渤海[17–18]、黄海[19–21]、长江口[22–23]等人类活动频繁的海域也有出现。
4.2 乐清湾大型底栖动物群落稳定性
Warwick[24]提出用ABC 曲线来反映环境污染对大型底栖动物群落稳定所带来的影响,并在后续的相关研究中得到了进一步的验证[25]。对于任何理化性质改变、生物扰动以及污染所引起的大型底栖动物群落稳定性变化,ABC 曲线均能灵敏地指示出来。本研究中,5 月份的群落稳定性最好,全站位和各群落组别均未受到明显的扰动;8 月份的群落稳定性最差,全站位和各群落组别均受到不同程度的扰动;1 月、3 月和10 月群落稳定性一般,虽然全站位未表现出扰动,但是优势群落组别均呈现不同程度的扰动。经初步分析8 月份乐清湾大型底栖动物的群落稳定性与海水质量密切相关。温州市夏季(6–8 月)的降雨量明显大于其他3 个季节[26],夏季充沛的雨量使得瓯江、清江、大荆溪等江河携带更多的污染物冲刷进入乐清湾,加上8 月份的高温加速了有机质等有害物质的分解和有机体的代谢,降低了水体中溶解氧的含量,导致其海水质量差。同时乐清湾内的两座燃煤电厂的热污染可能进一步加剧了海水质量的恶化。当然,影响乐清湾底栖动物群落稳定的原因还有很多,如人类活动导致的富营养化污染、过度捕捞导致的生态失衡、海底施工、船舶航道等都会造成乐清湾底栖动物群落的不稳定。
4.3 乐清湾大型底栖动物群落与环境因子的关系
大型底栖动物的群落结构很大程度上依赖于水文环境和理化条件[27]。周然等[28]、Glockzin 和Zettler[29]研究渤海湾和波美拉尼亚海湾表明水深与大型底栖动物群落结构呈显著性相关;李宝泉等[30]研究胶州湾表明软体动物的数量分布和季节变化与底质和水温存在较密切的关系;Jayaraj 等[31]对印度洋西北大陆架的大型底栖动物研究表明水温和盐度是影响大型底栖动物群落结构的主要环境因子;刘晓收等[32]、Carvalho等[33]研究认为沉积物中粉砂黏土含量与大型底栖动物群落结构显著相关。

图 5 乐清湾大型底栖动物群落特征种与环境因子的CCA 二维排序图Fig. 5 CCA biplot of macrobenthic community characteristic species and environmental factors in the Yueqing Bay
根据本研究年度BVSTEP 分析结果,在时间尺度上,亚硝酸盐是乐清湾大型底栖动物群落各月份差异的主要环境驱动因子。根据本研究各月份BVSTEP分析结果,在空间尺度上,盐度和氨氮分别是影响乐清湾3 月和5 月大型底栖动物的主要环境因子。本研究的水深和沉积物粒组含量与乐清湾的大型底栖动物群落结构不存在显著的相关性,盐度和水温在3 月份与乐清湾的大型底栖动物群落结构有较显著相关性,值得注意是营养盐(磷酸盐、亚硝酸盐、氨氮和硝酸盐)对乐清湾的大型底栖动物群落结构有较显著的影响,特别是磷酸盐和氨氮在时间和空间尺度上与乐清湾大型底栖动物群落结构均有较大相关性,见表2。
徐勇等[34]发现长江口大型无脊椎动物群落结构与磷酸盐存在显著性相关;彭松耀等[35]研究珠江口大型底栖动物群落结构发现磷酸盐和氨氮等环境因子与大型底栖动物群落结构存在较显著的相关性;刘坤等[36]研究厦门近岸海域大型底栖动物摄食功能群的分布与磷酸盐存在较大的相关性。磷酸盐对大型底栖群落结构的影响机理尚不清楚,生长速率假说(Growth Rate Hypothesis)指出环境中氮、磷的不同浓度变化会影响生物体的生长速率,从而影响种群的分布特征[37–38]。乐清湾受瓯江、清江、大荆溪等江河的长期持续影响,其磷酸盐、硝酸盐等营养盐输入丰富[39],因此可能是通过控制水体中营养盐的氮磷比例影响了浮游植物的生长状态,进而通过食物链的上行效应对大型底栖动物产生影响。
8 月份的BVSTEP 分析结果表明,沉积物粒组含量与大型底栖动物群落结构均不存在显著的相关性。这是由于乐清湾海域底质基本上都为黏土质粉砂,空间分布差异小,因此两者不存在显著性相关。通常生境异质性较高的区域沉积物粒组含量差异大,可以形成多种不同的小生境,具有较多的底栖动物物种数。乐清湾的沉积物空间分布差异小且颗粒较细,导致像一些双壳类动物由于受到较高含量悬浮物颗粒的影响而较难以生存,另外较细的沉积物环境下,一些在沉积物间隙活动的底栖动物分布也受到限制[40]。
4.4 乐清湾大型底栖动物群落特征种与环境因子的关系
根据本研究群落结构特征种与环境因子的CCA分析结果,水温对乐清湾大型底栖群落特征种时空分布影响最大。这是由于受太阳辐射影响,乐清湾海水水温在8 月份的平均温度可以达到(29.2±1.3)℃,1 月份的平均温度仅有(10.8±0.6)℃,季节变化显著。水温的变化直接影响着各种底栖动物的生长发育状况,如薄云母蛤在水温较低的月份可以形成较大的种群而成为优势物种,在水温较高的月份如8 月夏季则难以生存,相反另外一些物种如纽虫一种、双唇索沙蚕、异毛虫一种则在水温较高的月份才能得到较好的生长。
里加湾的长期研究表明,富营养化对大型底栖动物群落产生显著影响[41],渤海湾的研究表明大型底栖动物主要分布于含氮营养盐较低的水域[28]。在本研究中,除了冠奇异稚齿虫、全刺沙蚕属一种、腺袋才女虫等少数几种多毛类分布在含氮营养盐较高的水域,其他大部分群落特征种均分布在含氮营养盐较低的水域,表明乐清湾水体的无机氮浓度已经对该海域的大型底栖群落特征种产生了负面影响。因此,控制含氮营养盐的输入是维持乐清湾生态平衡的重要举措。
