黄土丘陵区生物结皮土壤温度年内动态特征及其对土壤水分的响应
2020-03-10王闪闪赵允格张子辉
王闪闪, 赵允格, 杨 光, 任 伟, 杨 凯, 张子辉
(1.中国科学院 水利部 水土保持研究所 黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨陵 712100;2.中国科学院大学, 北京 100049; 3.吴起县气象局, 陕西 吴起 717600; 4.西北农林科技大学 资源环境学院, 陕西 杨凌 712100)
生物土壤结皮(生物结皮),是由蓝绿藻、地衣和苔藓植物等自养生物和异养细菌、古菌及真菌等与土壤矿物质颗粒复合而形成的复杂复合体[1],是干旱半干旱地区土壤—大气间广泛存在的界面层,具有重要的生态功能,显著影响土壤—大气间的物质与能量的交换,以至于有研究者将其比喻为土壤的活性“皮肤”[2],是土壤温度重要的影响因子。在生物结皮及其生态功能研究的早期阶段,人们已经认识到生物结皮可影响土壤温度。如20世纪90年代,Belnap[3]在美国科罗拉多高原最早发现,以鞘藻属为主的藻结皮在夏季的温度可高于大气温度23 ℃,冬季高于大气温度14 ℃。此后,Kidron[4]在以色列内盖夫沙漠,以除去生物结皮的裸土为对照,采用便携式温度计观测证实,不同类型生物结皮可增加土壤温度3.9~6.9 ℃,增加程度与土层深度和含水量有关。同期,杨永胜等[5]在我国毛乌素沙地用曲管地温计测定了生物结皮土壤温度,结果表明生物结皮在气温和降水量较高时降低了地表土壤温度,而气温和降水量较低时增加了土壤温度,以此推测出生物结皮对土壤温度的影响受到气温和含水量的影响。肖波等[6]在黄土高原水蚀风蚀交错区的砂质土壤上通过定位监测,研究了藓结皮对土壤温度的影响,结果证实,夏季湿热条件下生物结皮对土壤温度的影响主要受大气温度和土壤水分调控,生物结皮降低了土壤温度;冬季干冷条件下主要受气温的影响,生物结皮可增加土壤温度。纵观已有研究可见,生物结皮对土壤温度的影响非常复杂,是多因素综合作用的结果,但目前有关生物结皮对土壤温度影响的研究仍非常有限,且主要在砂质土壤上展开,生物结皮对不同质地土壤温度的影响尚未可知,其对土壤温度的影响及其与土壤质地、土壤含水量及生物结皮类型等的相关性仍有研究的必要性,以进一步深化生物结皮生态功能的研究。土壤温度是土壤系统中诸多过程的重要影响因素,是土壤物理重要的研究内容。已有研究[7]表明,土壤温度不仅与土壤含水量有关,同时还与土壤质地密切相关,不同质地的土壤热特性不同。通常,在相同含水率条件下,砂粒含量越高,土壤的导热率越大,土壤导热能力越强[8-9]。生物结皮作为干旱半干旱地区土壤温度重要的影响因素,目前仍鲜有研究关注不同土壤质地土壤上生物结皮对土壤温度的影响,是生物结皮生态功能研究中的薄弱点。
黄土丘陵区退耕还林草生态工程实施后,生物结皮在该区退耕地大面积发育,平均盖度可达60%~70%[10],是该区土壤温度的重要影响因素,进而影响退耕地土壤系统中微生物的活动、养分的转化、根系的生长发育等诸多过程,但目前黄土丘陵区生物结皮影响下土壤温度的研究多集中在砂土地区,其他土质地区的研究鲜见报道,妨碍了人们对退耕地生态过程的准确认识。为此,本文以黄土丘陵区粉砂质壤土上发育至稳定阶段的生物结皮为研究对象,通过野外定位监测,观测生物结皮层和结皮层下5 cm土壤温度,及结皮层下5 cm土壤体积含水量动态。以2016年1月1日至2018年12月31日共3 a的数据为依据,分析生物结皮层和结皮层下5 cm土壤温度的年内动态特征以及对土壤含水量变化的响应,为进一步理解生物结皮在黄土丘陵区生态系统中的功能提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于黄土高原腹地,陕西省吴起县合沟小流域(北纬36°53′32″,东经108°13′26″),该小流域属于典型的黄土丘陵沟壑区,属暖温带半干旱季风气候[11],海拔1 450 m。年均气温为7.8 ℃。其中最高温7月,平均温度为22.7 ℃,最低温1月,平均温度-7.0 ℃。年均日照时数为2 400 h,多年年均降雨量为480 mm,主要集中在7—9月。研究区土壤类型为黄土母质上发育而成的黄绵土,植被覆盖度为40%~65%,常见植被有铁杆蒿(Artemisiagmelinii)、达乌里胡枝子(Lespedezadavurica)、匍匐委陵菜(Potentillareptans)、菊叶委陵菜(Potentillatanacetifolia)、狭裂白蒿(Artemisiakanashiroi)、硬质早熟禾(Poasphondylodes)、糙隐子草(Cleistogenessquarrosa)和长芒草(Stipabungeana)等。该流域自1999年退耕初期便实施严格封山禁牧制度,地表生物结皮广泛发育,平均盖度75.0%,主要类型包括藻结皮、藓结皮和藻、藓及地衣混生结皮[12],其中藓结皮优势种为土生对齿藓(Didymodonvinealis)和短叶对齿藓(Didymodontectorus),常见的地衣种类有Collemaspp.,Endocarponpusillum,Fulgensiabracteatad,Psoraspp.。样地土壤理化性质详见表1。
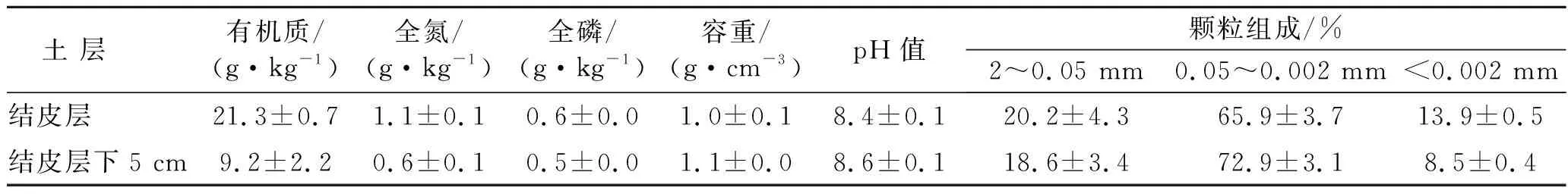
表1 样地土壤理化性质
1.2 研究方法
1.2.1 仪器安装及数据采集 于2015年9月10日,在合沟小流域的梁峁坡上选择发育稳定期的藻藓混合的生物结皮,垂直挖取10 cm深土壤剖面,将温度热电偶探头TT-T-24(Omega Engineering, Inc., Miami, FL, USA)垂直土壤剖面插入生物结皮层下1 cm(结皮厚度为6—8 mm,距离地表约2 cm深度处)和结皮层下5 cm土壤内。温度探头是T型探头,量程为-200~350 ℃,精度为±0.2 ℃。同时,在生物结皮层下5 cm土壤采用CS616-LC(Campbell Scientific, Inc., Logan, UT, USA)探头测定土壤体积含水量。含水量传感器分辨率为0.1%,精度为0.05%,量程为0%~50%。土壤温度和水分的测量频率是每10 s进行一次,水温监测传感器连接数据采集器CR1 000(Campbell Scientific, Inc., Logan, UT, USA),将数据保存到设定好的表格中,每10 min计算土壤温度(℃)和含水量(V/V%)的平均值,并将数据进行采集和存储。其后,每两个月下载数据。气象参数及大气温度由吴起气象局提供,该局测定点距离研究样地监测点3.9 km,气象局提供的年降雨数据与试验地雨量计监测数据一致。
1.2.2 数据处理 所用数据的记录时段为2016年1月1日至2018年12月31日。
(1) 为明确生物结皮层和结皮层下5 cm土壤温度年内变化,将相同月份内监测的温度数据进行平均,分析生物结皮层和结皮层下5 cm土壤温度的年内动态;将相同月份同一时间所监测的温度数据进行平均,以此分析不同月份生物结皮层及结皮层下5 cm土壤温度日动态。
(2) 为明确最冷月和最热月土壤含水量对生物结皮层及结皮层下5 cm土壤温度的影响,根据土壤导热方式的差异(土壤体积含水量小于4%以固体、气体及辐射参与导热。随着含水量的增加,水分逐渐填充土壤颗粒的间隙,土壤体积含水量大于12%时以固体、液体导热为主[13])。选取7月(最热月,雨热同期)晴朗天气、大气平均温度为20~25 ℃的自然天内日均土壤含水量≥12%和土壤含水量≤4%的生物结皮层和结皮层下5 cm土壤温度数据,分析了7月这两个土壤含水量对生物结皮及结皮层下5 cm土壤温度的影响;选择1月(最冷月)天气晴朗的自然天内日均土壤含水量≥12%和土壤含水量≤4%的生物结皮层和结皮层下5 cm土壤温度数据,分析1月土壤含水量对生物结皮层及结皮层下5 cm土壤温度的影响。需要指出的是,冬季土壤冻结后,水分探头测量结果显示在0%~5%之间变化,可能不是实际土壤含水量,但是日间升温后,土壤解冻,水分含量增加,因此我们根据一天的平均土壤含水量来选择土壤含水量≥12%和土壤含水量≤4%的生物结皮土壤温度数据。选择的天数及日期详见表2。
1.3 统计分析
采用Excel 2010(Microsoft Corp., Redmond, WA, USA)和SPSS 19.0(SPSS Inc., Chicago, IL, USA)软件对数据进行统计分析。其中,大气和生物结皮层及结皮层下5 cm土壤温度年内变化的差异显著性进行One-way ANOVA方差分析,并用LSD法进行多重比较(α=0.05)。采用Pearson相关分析研究不同月份生物结皮土壤日均温与大气日均温、日均土壤含水量之间的相关性。利用Origin Pro 9.1(Origin Lab Corp., Northampton, MA, USA)进行作图分析,数据表示为平均值±标准误。
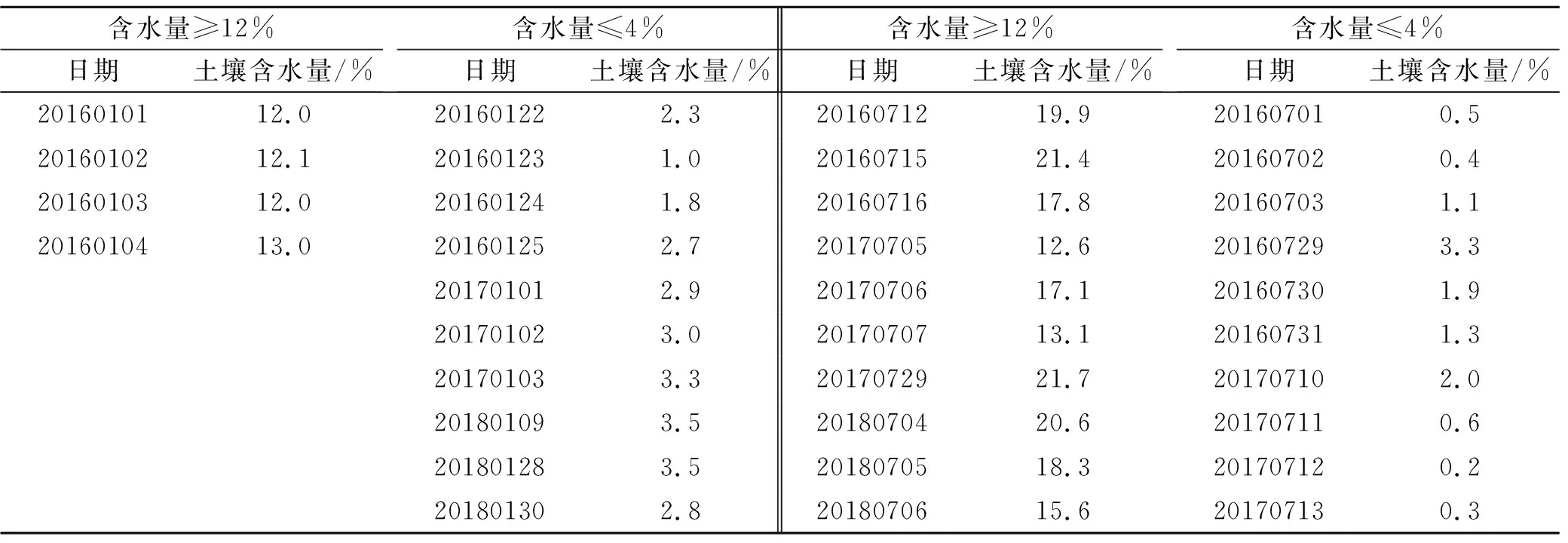
表2 1月和7月所选择的土壤含水量≥12%和土壤含水量≤4%的日期
2 结果与分析
2.1 生物结皮层及下层土壤温度年内动态
统计分析3 a的数据表明,3 a中有1 096 d中大气日均温高于生物结皮层日均温的天数约50 d,其余1 046 d大气温度均低于生物结皮层温度。根据3 a年均温可知(图1),3 a研究区大气年均温为8.7 ℃,生物结皮层及结皮层下5 cm土壤年均温分别为13.6 ℃和12.9 ℃。与气温相比,生物结皮层和结皮层下5 cm土壤年均温分别高于大气4.9 ℃和4.3 ℃。图2为2016—2018年生物结皮层及结皮层下5 cm深度的月均土壤温度、气温以及降水量随月份的变化特征,生物结皮层和结皮层下5 cm土壤温度的变化与气温、降水量的变化均呈正弦波型,生物结皮层月均温最高在6月,温度为27.1 ℃,结皮层下5 cm土壤和大气月均温最高在7月,温度分别为22.7和25.1 ℃。大气、生物结皮层和结皮层下5 cm土壤月均温最低均在1月,分别为-6.9 ℃,-1.7 ℃和-1.4 ℃。

注:不同小写字母表示差异显著(p<0.05)

图2 月均降水量以及生物结皮层、结皮层下5 cm土壤和大气月均温年内动态
2.2 不同月份生物结皮层及下层土壤温度日动态
图3为不同月份生物结皮层和结皮层下5 cm土壤与大气温度日动态。由图3可知,生物结皮层和结皮层下5 cm土壤温度的日变化特征明显,呈正弦曲线变化,与气温的日变化基本一致,有一个谷值和一个峰值。生物结皮层和结皮层下5 cm土壤温度日较差在雨季前(4—6月)变化最大,最大日较差出现在6月,生物结皮层和结皮层下5 cm土壤分别为32.0 ℃和17.2 ℃。7月之后,生物结皮层和结皮层下5 cm土壤温度日较差逐渐变小,10月生物结皮层温度日较差最小,为17.3 ℃,1月结皮层下5 cm土壤温度日较差最低为8.5 ℃。同时,生物结皮与大气的温差在雨季前最大,7月之后(雨季中后期)逐渐缩小。
表3为生物结皮土壤温度与气温和土壤含水量的相关性分析结果。一年12个月中,生物结皮土壤温度与大气温度均呈极显著正相关;3—10月生物结皮土壤温度与土壤含水量呈极显著负相关,3月的相关系数最小,7月的相关系数最大;11和12月生物结皮土壤温度与土壤含水量呈极显著正相关。由此可见,不同月份生物结皮土壤温度受水分影响的程度和结果不同。

图3 大气和生物结皮层及结皮层下5cm土壤温度不同月份日动态
2.3 1月和7月土壤含水量对生物结皮层及下层土壤温度的影响
图4为1月土壤含水量≥12%和土壤含水量≤4%时大气和生物结皮层以及结皮层下5 cm土壤温度日动态。当含水量≥12%时,生物结皮层、结皮层下5 cm土壤和大气温度日较差分别是15.9 ℃,5.6 ℃和18.8 ℃。当含水量≤4%时,生物结皮层、5 cm土壤和大气温度日较差分别是26.8 ℃,12.3 ℃和11.4 ℃。
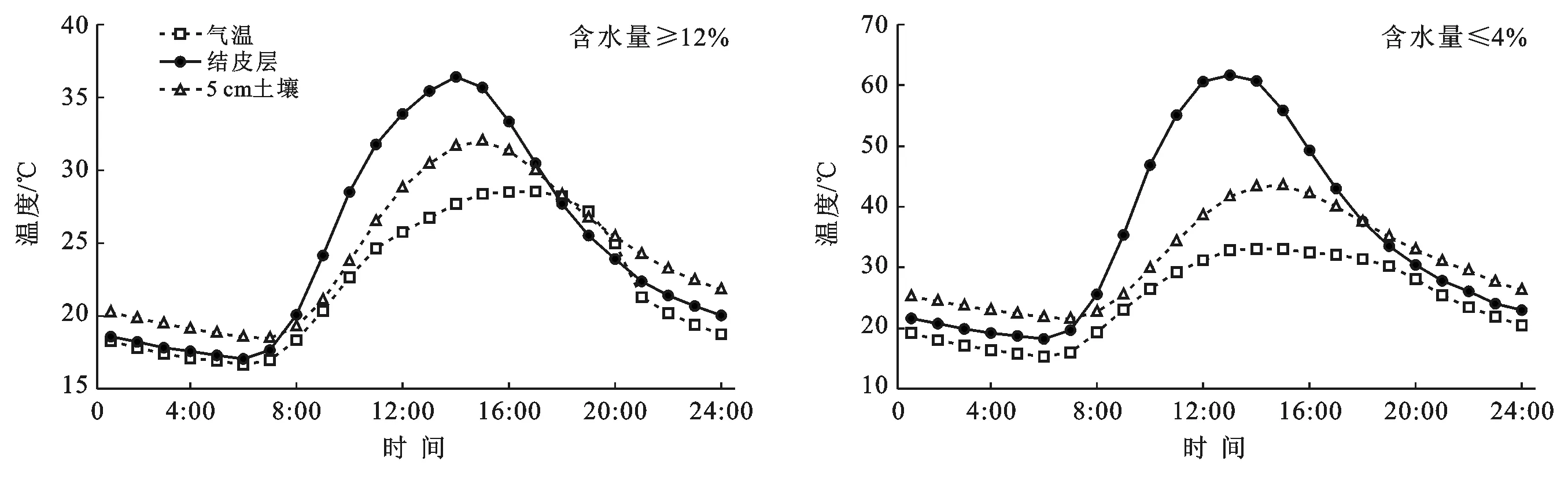
图4 1月土壤含水量≥12%和土壤含水量≤4%时大气和生物结皮层及结皮层下5 cm土壤温度日动态

表3 不同月份生物结皮土壤温度与土壤含水量和气温之间的Pearson相关性分析
图5为7月土壤含水量≥12%和土壤含水量≤4%时大气和生物结皮层以及结皮层下5 cm土壤温度日动态。当含水量≥12%时,生物结皮层、结皮层下5 cm土壤和大气温度日较差分别是19.4 ℃,13.5 ℃和11.8 ℃。当含水量≤4%时,生物结皮层、结皮层下5 cm土壤和大气温度日较差分别是35.2 ℃,20.8 ℃和17.8 ℃。
3 讨 论
生物结皮是土壤—大气之间物质、能量交换的界面层,显著影响土壤温度。本研究以黄土丘陵区发育至稳定阶段的生物结皮为研究对象,研究生物结皮土壤温度的年内动态以及对土壤水分的响应。结果表明,受太阳辐射的影响,气温、生物结皮层与结皮层下5 cm土壤温度年内动态均呈正弦曲线,与该区表层土壤温度变化趋势一致[14]。结皮层下5 cm土壤和大气温度月均温最高在7月,而生物结皮层在6月。一方面,可能是因为该区降雨主要在7—9月,6月降水较少,土壤含水量也较低,生物结皮土壤热容量和导热率较小,吸收相同的热量后,6月生物结皮层温度升高较快[7],进而导致生物结皮土壤温度在该月最高。另一方面,6月生物结皮层多数时间并未湿润颜色多为黑灰色,有利于吸收太阳辐射[3],使得生物结皮土壤升温较快。肖波等在黄土高原水蚀风蚀交错区的砂土上的研究表明夏季生物结皮对土壤温度的影响主要与气温和降水有关,而冬季主要受气温调控[6]。但是本文结果表明,黄土丘陵区冬季的部分月份土壤水分与生物结皮土壤温度呈显著正相关,可见该区冬季土壤含水量也是影响生物结皮温度的重要因素。

图5 7月土壤含水量≥12%和土壤含水量≤4%时大气和生物结皮层及结皮层下5 cm土壤温度日动态
通过分析冬季1月黄土丘陵区粉砂质壤土上的生物结皮土壤温度日动态与土壤水分的关系,与砂土区同为1月的结果略有不同。主要体现在黄土丘陵区土壤含水量较低时生物结皮层及结皮层下5 cm土壤温度日较差是土壤含水量较高时的1.7倍以上。两个土壤含水量条件下的生物结皮土壤最高温相差不大,而差异主要在最低温处。土壤含水量较高时,夜间土壤中的水会结冰放热[15],使得生物结皮土壤最低温为-3 ℃,而同时段土壤含水量较低时生物结皮土壤最低温为-17 ℃,进而使得在土壤含水量较高时生物结皮土壤温度日振幅较小。可见1月土壤含水量也是影响生物结皮土壤温度的重要因子。而肖波等在黄土高原砂土上的结果显示土壤含水量在冬季整体较低,对生物结皮土壤温度的影响有限,而黄土丘陵区粉砂质壤土的土壤水分状况要优于砂土区,因此两个区域在冬季1月土壤水分对生物结皮温度的影响存在差异。这可能是因为两个地区土壤质地不同,砂土的土壤孔隙较大,水分入渗和蒸发量都高于粉砂质壤土[4],而粉砂质壤土的土壤颗粒较细,土壤孔隙较小,对水的吸持能力较大,土壤水分状况相对较好。另外,在比较两个区域同为低含水量条件且气温日较差一致的情况下,砂土区生物结皮土壤的日较差低于黄土丘陵区5 ℃,经过对比发现,砂土上生物结皮土壤最高温低于黄土丘陵区粉砂质壤土最高温约5 ℃,这可能是由于以下两个原因导致的:一方面是因为黄土丘陵区纬度低于砂土区,冬季接受的太阳辐射多于砂土区[16]。另一方面,砂土的导热率高于黄土丘陵区的粉砂质壤土,吸收的热量易传导到下层[7],使表层生物结皮土壤温度不易升高,因此砂土上生物结皮土壤温度低于粉砂质壤土上生物结皮土壤温度。以上结果表明,黄土丘陵区粉砂质壤土上生物结皮土壤温度动态对土壤水分的响应程度不同于黄土高原水蚀风蚀交错区的砂土,进而对土壤温度的影响亦不同,因此有关黄土丘陵区生物结皮对土壤温度的影响有待进一步研究。
生物结皮作为黄土丘陵区土壤微生物和小型节肢动物的栖息地,其温度变化对微生物活性以及小型节肢动物的生存至关重要[1]。同时,雨季前(4—6月)生物结皮土壤日较差高于雨季中后期,这一现象对储存于生物结皮的植物种子的萌发有促进作用[17]。明确生物结皮土壤温度动态特征及与土壤水分的关系,对下伏土壤中的微生物活性、土壤动物生存及植被生长具有重要意义。
4 结 论
(1) 通过3 a野外定位监测发现,受降水和气温的影响,生物结皮土壤年内动态和日动态均按正弦曲线变化。生物结皮层月均温最高出现在6月,为27.1 ℃;结皮层下5 cm土壤月均温最高在7月,为22.7 ℃;生物结皮层和结皮层下5 cm土壤月均温均在1月最低,分别为-1.7 ℃和-1.4 ℃。6月生物结皮层和结皮层下5 cm土壤温度日较差最大,分别为32.0 ℃和17.2 ℃;10月生物结皮层温度日较差最小为17.3 ℃,1月结皮层下5 cm土壤温度日较差最低为8.5 ℃。
(2) 土壤水分在最冷月(1月)和最热月(7月)均会影响黄土丘陵区粉砂质壤土上生物结皮土壤温度。土壤含水量的增加导致生物结皮层和结皮层下5 cm土壤温度日较差下降。1月土壤含水量≥12%时生物结皮层和结皮层下5 cm土壤温度日较差分别是15.9 ℃和5.6 ℃;当含水量≤4%时生物结皮层和结皮层下5 cm土壤温度日较差分别是26.8 ℃和12.3 ℃。7月受气温和降雨的影响,当土壤含水量≥12%时,生物结皮层和结皮层下5 cm土壤温度日较差分别是19.4 ℃和13.5 ℃;当含水量≤4%时,生物结皮层和结皮层下5 cm土壤温度日较差分别是35.2 ℃和20.8 ℃。
