云南省高原粳稻主要农艺性状与产量的多重分析
2020-03-08蒋聪段玉云杨旭昆吴志刚邹茜
蒋聪 段玉云 杨旭昆 吴志刚 邹茜
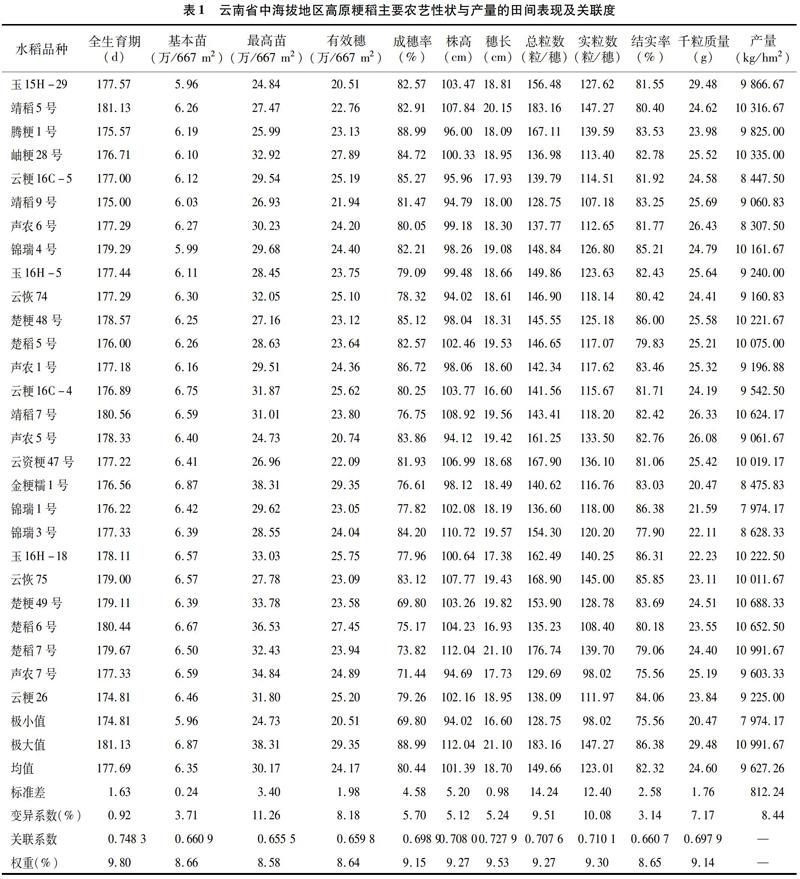
摘要:明确云南省不同海拔稻作区的选择目标,为云南省高原粳稻的品种选育提供一定的理论依据。在云南省中海拔地区种植27个高原粳稻品种,高海拔地区种植13个高原粳稻品种,采集2017年云南省区域试验的12个性状田间表现数据,运用相关分析、通径分析、主成分分析及灰色关联度的方法,研究分析了云南省不同海拔地区的高原粳稻品种主要农艺性状与产量之间的关系。结果表明:(1)在中海拔地区云南省高原粳稻品种的12个性状的变异系数在0.92%~11.26%之间;产量与全生育期呈极显著正相关(r=0.601 1* *),与株高、总粒数和实粒数呈显著正相关(r=0.427*、r=0.425*、r=0.393*),与其余7个农艺性状相关但均不显著;通径分析表明,全生育期、最高苗、成穗率、株高、实粒数和千粒质量对产量的直接作用为正值,基本苗、有效穗、穗长、总粒数和结实率对产量的直接作用为负值;主成分分析表明,对产量贡献大小排序为总粒数因子>基本苗因子>千粒質量因子>结实率因子>有效穗因子>穗长因子;产量与11个性状的关联度大小依次为全生育期>穗长>实粒数>株高>总粒数>成穗率>千粒质量>基本苗>结实率>有效穗>最高苗。(2)在高海拔地区云南高原粳稻品种的12个性状的变异系数在1.82%~9.99%之间;产量与株高呈极显著正相关(r=0.738 8* *),与成穗率、总粒数和实粒数呈显著正相关(r=0.618 2*、r=0.615 4*、r=0.556 2*),与其余7个农艺性状相关但均不显著;通径分析表明,最高苗、成穗率、株高、总粒数和结实率对产量的直接作用为正值,全生育期、基本苗、有效穗、穗长、实粒数和千粒质量对产量的直接作用为负值;主成分分析表明,对产量贡献大小排序为总粒数因子>基本苗因子>结实率因子>成穗率因子;产量与农艺性状的关联度大小依次为总粒数>株高>实粒数>穗长>结实率>成穗率>基本苗>最高苗>有效穗>全生育期>千粒质量。结果表明,在云南省高原稻区选育适宜云南中海拔地区的常规粳稻品种时应注重选择生育期偏长、株高偏高及千粒质量与有效穗适中的大穗多粒型品种,而在云南省高原稻区选育适宜云南高海拔地区的常规粳稻品种时应注重选择生育期适中、株高偏高、成穗率高及千粒质量与有效穗适中的大穗多粒型的品种类型,并注意性状间的协调发展,以充分表现品种潜力,选育出优异的高原粳稻品种。
关键词:高原粳稻;农艺性状;相关分析;通径分析;主成分分析;灰色关联度分析
中图分类号:S511.2+20.4 文献标志码:A 文章编号:1002-1302(2020)21-0074-10
中国65%的人口都以水稻为主食,在农作资源贫乏的地区还将水稻作为生计作物。至2018年统计,我国水稻种植面积9 968.5万hm2,比2017年减少107.9万hm2,下降1.1%[1-3]。近10年来,我国水稻产量一直处于停滞状态。2018年的水稻总产量比2017年减少了9%。影响水稻总产量的外因:耕地面积下降、水资源和气候危机、农村劳动力短缺、市场对优质大米的需求。水稻生产系统面临的关键问题是水稻遗传背景狭窄、肥料和杀虫剂过度使用致使土壤被破坏、农村农作物管理过于简单化[4]。
水稻生产持续增长对实现国家粮食安全有重要作用。水稻产量的增加主要依靠单位面积产量的提高而不是种植面积的扩大需要通过整合中国种质、基因组资源和分子技术,发展绿色超级稻。这些绿色超级稻品种将提高对主要病虫害的抗性、养分利用的高度科学性、主要非生物胁迫(如干旱、盐分、异常温度的抗性)、产量和粮食品质。可以预见,绿色超级稻的发展将使中国水稻产量增加。
云南省自海拔100 m以下至海拔2 700 m均有水稻种植,南北跨越维度将近80°。由于境内地形复杂、地势高低悬殊,垂直气候分布差异非常显著。云南省常规粳稻区域试验通常鉴定粳稻品种的丰产性、稳定性、抗逆性等指标,针对云南省“立体气候”筛选出适合的栽培品种对促进云南粳稻生产具有重要的意义[5-7]。相关研究结果表明,水稻的单株产量与水稻的千粒质量、结实率、有效穗数、每穗的实粒数之间具有非常复杂的关系,为了研究主要性状与产量的关系,以往大多科研工作者都采用简单的相关或多元回归分析法。然而简单相关分析结果并不能很全面体现变数间的相互关系,致使分析结果带有一定的片面性,而且在品种选育过程中,品种(品系)的优劣本身就是基因和环境互作的外在体现,本研究运用相关分析、通径分析、主成分分析及灰色关联度的方法对2016—2017年度云南省中海拔常规粳稻区域试验的27个水稻品种,2016—2017年度云南省高海拔常规粳稻区域试验的13个水稻品种进行综合评估,以期明确其间的关系,为品种选育、推广和水稻生产提供科学依据。
1 材料与方法
1.1 试验材料
试验材料为参加2016—2017年度云南省中海拔常规粳稻区域试验的27个水稻品种(玉15H-29、靖稻5号、腾粳1号、岫粳28号、云粳16C-5、靖稻9号、声农6号、锦瑞4号、玉16H-5、云恢74、楚粳48号、楚稻5号、声农1号、云粳16C-4、靖稻7号、声农5号、云资粳47号、金粳糯1号、锦瑞1号、锦瑞3号、玉16H-18、云恢75、楚粳49号、楚稻6号、楚稻7号、声农7号和云粳26),参加2016—2017年度云南省高海拔常规粳稻区域试验的13个水稻品种(云粳46、昆粳10号、云粳16K-5、云粳37号、靖稻6号、靖稻8号、凤12-17、昭粳13号、丽粳22号、丽粳23号、云资粳47号、师粳8号和凤稻23号)。
1.2 试验设计与方法
中海拔常规粳稻于2016—2017年度种植于云南省陆良县(1 840 m)、麒麟区(1 840 m)、泸西县(1 705 m)、峨山县(1 553 m)、牟定县(1 780 m)、弥渡县(1 720 m)、澜沧县(1 456 m)、隆阳区(1 650 m)和腾冲市(1 648 m)的试验田,选择肥力均匀一致、中等偏上的田块。高海拔常规粳稻于2016—2017年度种植于云南省寻甸县(1 874 m)、昭阳区(1 900 m)、马龙区(2 032 m)、会泽县(1 940 m)、大理市(1 984 m)、祥云县(1 940 m)、剑川县(2 200 m)和永胜县(2 240 m)试验田,选择肥力均匀一致、中等偏上的田块。采用随机区组排列,重复3次,安排在同一块田上,小区面积统一为13.33 m2(长方形),播种、移栽时节及栽植密度、施肥量、田间管理要求以当地最佳节令和方式进行,旱育秧或湿润育秧,栽培技术略高于当地生产水平,同一生产措施当天完成。试验期间防虫、不防病,各试点基本按照云南省水稻品种区域试验观察记载项目与标准方法及标准的要求进行观察记载。
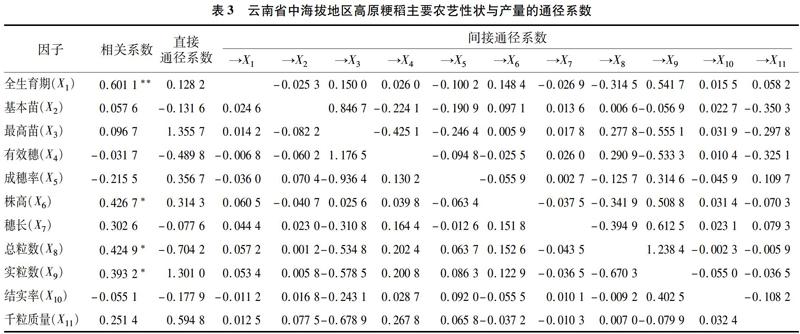
成穗率对产量的直接通径系数为0.356 7;成穗率通过基本苗、有效穗、穗长、实粒数和千粒质量对产量间接地起正向作用(0.627 6),通过全生育期、最高苗、株高、总粒数和结实率对产量间接地起负向作用(-1.199 9),但正向效应远小于负向效应。综上成穗率对产量总效应较小,为-0.215 5,因此在育种选择中应尽量选择成穗率适中的类型。
株高对产量的直接通径系数为0.314 3;株高通过全生育期、最高苗、有效穗、实粒数和结实率对产量间接地起正向作用(0.666 1),通过基本苗、成穗率、穗长、总粒数和千粒质量对产量间接地起负向作用(-0.553 8),但正向效应大于负向效应。综上株高对产量总效应较小,为0.426 7,因此在育种选择中应尽量选择株高偏高的类型。
穗长对产量的直接通径系数为-0.077 6;穗长通过全生育期、基本苗、有效穗、株高、实粒数、结实率和千粒质量对产量间接地起正向作用(1.098 5),通过最高苗、成穗率和总粒数对产量间接地起负向作用(-0.718 3),但正向效应大于负向效应。综上穗长对产量总效应较小,为-0.215 5,因此在育种选择中应尽量选择成穗率适中的类型。
总粒数对产量的直接通径系数为-0.704 2;总粒数通过全生育期、基本苗、有效穗、成穗率、株高和实粒数对产量间接地起正向作用(1.715 5),通过最高苗、穗长、结实率和千粒质量对产量间接地起负向作用(-0.586 5),但正向效应远远大于负向效应。综上总粒数对产量总效应较大,为0.302 6,因此在育种选择中应尽量选择总粒数偏大的类型。
实粒数对产量的直接通径系数为1.301 0;实粒数通过全生育期、基本苗、有效穗、成穗率和株高对产量间接地起正向作用(0.469 2),通过全生育期、最高苗、株高、总粒数和结实率对产量间接地起负向作用(-1.376 8),但正向效应远小于负向效应。综上实粒数对产量总效应较大,为0.393 2,因此在育种选择中应尽量选择实粒数偏多的类型。
结实率对产量的直接通径系数为-0.177 9,结实率通过基本苗、有效穗、穗长、实粒数和千粒质量对产量间接地起正向作用(0.550 1),通过全生育期、最高苗、株高、总粒数和结实率对产量间接地起负向作用(-0.427 2),但正向效应大于负向效应。综上成穗率对产量总效应较小,为-0.055 1,因此在育种选择中应尽量选择结实率适中的类型。
千粒质量对产量的直接通径系数为0.594 8,基本苗通过全生育期、基本苗、有效穗、成穗率、总粒数和实粒数对产量间接地起正向作用(0.463 0),通过最高苗、株高、穗长和实粒数对产量间接地起负向作用(-0.806 3),但正向效应小于负向效应。综上千粒质量对产量总效应较小,为0.251 4,因此在育种选择中应尽量选择千粒质量适中的类型。
2.1.5 云南省中海拔地区高原粳稻主要农艺性状
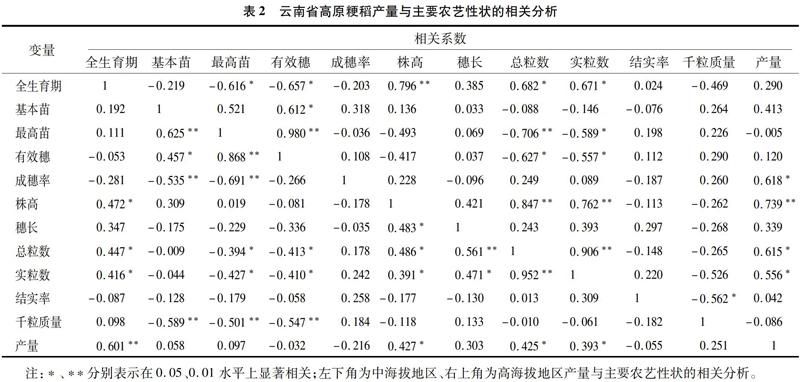
与产量的主成分分析 为了进一步解析云南中海拔地区高原粳稻主要农艺性状与产量的关系 对其主要农艺性状与产量进行主成分分析,从表4可以看出,分析结果显示:利用主成分分析法将中海拔地区的27个粳稻品种的12个主要性状与产量转化为6个主分量所构成的信息量为总信息量的89.653 3%(>85%),因此提取的6個主分量可以概括云南中海拔地区高原粳稻主要农艺性状与产量的的绝大部分信息,说明这6个因子可以作为主分量。这6个主分量中,第1主分量可代表总粒数、实粒数和最高苗,载荷量分别为0.413 1、0.420 5和-0.373 4;第2主分量可代表基本苗、株高和全生育期,载荷量分别为0.402 0、0.369 0、0.366 6;第3主分量可代表千粒质量,载荷量为-0.562 9;第4主分量可代表结实率和产量,载荷量分别为0.549 9和0.510 3;第5主分量可代表有效穗和成穗率,载荷量分别为0.596 2和0.588 6;第6主分量可代表穗长,载荷量为0.726 1。这6个主分量中,仅仅第1、2、3主分量的特征值大于1,且累计贡献率为71.037 2%,说明第1、2、3主分量含有大部分信息,相对贡献较大。说明云南中海拔地区高原粳稻的总粒数、实粒数、最高苗、基本苗、株高、全生育期和千粒质量7个性状相对品种的综合表现影响更大,在6个性状中仅仅千粒质量的载荷量为负值,其余均为正值,说明在云南省中海拔地区高原粳稻育种时,应尽量选择蘖多粒多、株高、全生育期偏长但千粒质量适中的品种。
2.2 云南省高海拔地区高原粳稻主要农艺性状与产量的多重分析
2.2.1 云南省高海拔地区高原粳稻主要农艺性状与产量田间表现 在云南省高海拔地区8个地点种植的13个高原粳稻常规品种的12个性状表现平均值见表5,其中产量变幅为9 304.69~11 784.38 kg/hm2,产量最高的为丽粳23号,最低的是靖稻6号。变异系数通常作为评价所考察性状受环境条件影响而发生变异程度的指标,因此在考察高原粳稻主要农艺性状与产量的变异系数的大小可反映品种在实际生产中的丰产性及稳产性。从表5可以看出,在中海拔地区的13个高原粳稻的12个性状的变异系数在1.82%~9.99%之间,变异系数大小顺序为实粒数>总粒数>有效穗>最高苗>产量>株高>结实率>千粒质量>穗长>成穗率>全生育期>基本苗,可见实粒数的变异系数最大,总粒数、有效穗、最高苗、产量、株高、结实率、千粒质量、穗长、成穗率和全生育期次之,基本苗的变异系数最小,表明选育云南省高海拔地区高原粳稻时其实粒数、总粒数、有效穗和最高苗改良潜力较大,其次是产量、株高、结实率和千粒质量,而穗长、成穗率、全生育期和基本苗改良潜力较小。
2.2.2 云南省高海拔地区高原粳稻主要农艺性状与产量的灰色关联度分析 以高海拔云南高原常规粳稻的农艺性状作为子序列,以产量性状作为母序列进行灰色关联度分析(表5)。从表5可以看出,13个高海拔云南高原常规粳稻品种农艺性状与产量的关联度大小依次为总粒数>株高>实粒数>穗长>结实率>成穗率>基本 苗> 最高苗>有效穗>全生育期>千粒质量。由此看出,在高海拔地区云南高原常规粳稻的总粒数与其产量关联度最大,在高海拔地区云南高原常规粳稻的总粒数是影响云南高原常规粳稻产量的最主要因素,次要因素依次是株高、实粒数、穗长、结实率、成穗率、基本苗、最高苗、有效穗、全生育期和千粒质量。而高海拔地区云南高原常规粳稻的产量3要素实粒数、有效穗和千粒质量与产量关联度依次为0.730 9、0.628 0和0.587 0,说明高海拔地区云南高原常规粳稻的实粒数对产量贡献较大,其次为有效穗和千粒质量。
2.2.3 云南省高海拔地区高原粳稻主要农艺性状与产量的相关分析 以云南高海拔地区高原粳稻的产量为因变量,11个性状为自变量进行相关分析,结果见表2(右上角)。云南高海拔地区高原粳稻11个性状与产量的相关系数大小排序依次为株高>成穗率>总粒数>实粒数>基本苗>穗长>全生育期>有效穗>千粒质量>结实率>最高苗,其中株高与产量呈极显著正相关(r=0.739* *),成穗率、总粒数和实粒数与产量呈显著正相关(r=0.618、r=0.615、r=0.556),其余7个农艺性状与产量相关但均不显著。结果表明,在选育适宜云南高海拔地区的高原粳稻品种时,应注重株高、成穗率、总粒数和实粒数等目标性状的优化,即选育株高偏高、成穗率高的多粒型品种,但是在关注目标性状的同时还应考虑到性状间相互作用,促进各性状间协调发展,选育综合性状优良的高原粳稻品种。
2.2.4 云南省高海拔地区高原粳稻主要农艺性状与产量的通径分析 为进一步比较云南高海拔地区高原粳稻各主要农艺性状对其产量的贡献大小及相对重要性,对11个主要农艺性状与产量进行了通径分析(表6)。从表6可以看出,云南高海拔地区高原粳稻的11个主要农艺性状对产量直接贡献的大小依次为是最高苗>有效穗>株高>总粒 数> 全生育期>实粒数>结实率>成穗率>千粒质量>穗长>基本苗,但是最高苗、成穗率、株高、总粒数和结实率5个性状的直接效应对产量有正向作用,全生育期、基本苗、有效穗、穗长、实粒数和千粒质量6个性状的直接效应对产量有负向作用;最高苗(P=4.262 1)、有效穗(P=3.752)和株高(P=1.870 6)的通径系数较大即对产量的直接贡献较大,在云南高海拔地区高原粳稻优良品种的育种过程中应重视对这最高苗、有效穗和株高3个性状的改良且兼顾其他性状,更易获得综合性状较好的高产品种。
全生育期对产量的直接通径系数为-1.369 9;全生育期通过基本苗、有效穗、株高、总粒数、结实率和千粒质量6个性状对產量间接地起正向作用(5.175 5),通过最高苗、成穗率、穗长和实粒数对产量间接地起负向作用(-3.515 6),但正向效应大于负向效应。综上全生育期对产量总效应较小,为0.289 9,因此在育种选择中应尽量选择全生育期适中的品种。
基本苗对产量的直接通径系数为-0.111 5;基本苗通过全生育期、最高苗、成穗率、株高和实粒数对产量间接地起正向作用(3.042 3),通过有效穗、穗长、总粒数、结实率和千粒质量对产量间接地起负向作用(-2.517 6),但正向效应大于负向效应。综上基本苗对产量总效应较小,为0.413 1,因此,在育种选择中应尽量选择基本苗适中的类型。
最高苗对产量的直接通径系数为4.262 1,对产量的直接贡献最大;最高苗通过全生育期、实粒数和结实率对产量间接地起正向作用(1.591 9),通过基本苗、最高苗、有效穗、成穗率、实粒数和千粒质量对产量间接地起负向作用,综合看正向效应远低于负向效应(-5.858 8)。综上最高苗对产量总效应较小,为-0.004 9,因此在育种选择中分蘖力适中的类型。
有效穗对产量的直接通径系数为-3.752 0;有效穗通过全生育期、最高苗、成穗率、实粒数和结实率对产量间接地起正向作用(5.792 1),通过基本苗、株高、穗长、总粒数和千粒质量对产量间接地起负向作用(-1.920 4),但正向效应远大于负向效应。综上有效穗对产量总效应较小,为0.119 6,因此在育种选择中应尽量选择有效穗适中的类型。
成穗率对产量的直接通径系数为0.313 5;成穗率通过全生育期、株高、穗长和总粒数对产量间接地起正向作用(1.118 2),通过基本苗、最高苗、有效穗、实粒数、结实率和千粒质量对产量间接地起负向作用(-0.813 5),但正向效应大于负向效应。综上成穗率对产量总效应较大,为0.618 2,因此在育种选择中应尽量选择成穗率高的类型。
株高对产量的直接通径系数为1.870 6;株高通过有效穗、成穗率、总粒数和千粒质量对产量间接地起正向作用(3.053 3),通过全生育期、基本苗、最高苗、穗长、实粒数和结实率对产量间接地起负向作用(-4.185 2),但正向效应小于负向效应。综上株高对产量总效应较大,为0.738 8,因此在育种选择中应尽量选择株高偏高的类型。
穗长对产量的直接通径系数为-0.134 2;穗长通过最高苗、株高、总粒数、结实率、千粒质量对产量间接地起正向作用(1.626 8),通过全生育期、基本苗、有效穗、成穗率和实粒数对产量间接地起负向作用(-1.153 9),但正向效应大于负向效应。综上穗长对产量总效应较小,为0.338 6,因此在育种选择中应尽量选择成穗率适中的类型。
总粒数对产量的直接通径系数为1.613 0;总粒数通过基本苗、有效穗、成穗率、株高和千粒质量对产量间接地起正向作用(4.076 1),通过全生育期、最高苗、穗长、实粒数和结实率对产量间接地起负向作用(-5.073 9),但正向效应小于负向效应。综上总粒数对产量总效应较大,为0.615 4,因此在育种选择中应尽量选择总粒数偏大的类型。
实粒数对产量的直接通径系数为-1.157 5;实粒数通过基本苗、有效穗、成穗率、株高、总粒数、结实率和千粒质量对产量间接地起正向作用(5.194 4),通过全生育期、最高苗和穗长对产量间接地起负向作用(-3.480 6),但正向效应大于负向效应。综上实粒数对产量总效应较大,为0.556 2,因此在育种选择中应尽量选择实粒数偏多的类型。
结实率对产量的直接通径系数为0.340 4,结实率通过基本苗、最高苗和千粒质量对产量间接地起正向作用(0.957 2),通过全生育期、有效穗、成穗率、株高、穗长、总粒数和实粒数对产量间接地起负向作用(-1.255 9),但正向效应小于负向效应。综上成穗率对产量总效应较小,为0.042 0,因此在育种选择中应尽量选择结实率适中的类型。
千粒质量对产量的直接通径系数为-0.190 0,千粒质量通过全生育期、最高苗、成穗率、穗长和实粒数对产量间接地起正向作用(2.330 9),通过基本苗、有效穗、株高、总粒数和结实率对产量间接地起负向作用(-2.226 4),但正向效应小于负向效应。综上成穗率对产量总效应较小,为-0.085 7,因此在育种选择中应尽量选择千粒质量适中的类型。
2.2.5 云南省中海拔地区高原粳稻主要农艺性状与产量的主成分分析 为了进一步解析云南省高海拔地区高原粳稻主要农艺性状与产量的关系,对其主要农艺性状与产量进行主成分分析,结果见表4。从表4可以看出,利用主成分分析法将高海拔地区的13个粳稻品种的12个主要性状与产量转化为4个主分量所构成的信息量为总信息量的 87.231 4%(>85%),因此提取的4个主分量可以概括云南中海拔地区高原粳稻主要农艺性状与产量的的绝大部分信息,说明这4个因子可以作为主分量。第1主分量可代表总粒数、实粒数、株高、全生育期和最高苗,载荷量分别为0.413 2、0.408 1、0.383 4、0.387 1和-0.343 1;第2主分量可代表产量、基本苗和有效穗,载荷量分别为0.511 6、0.491 6 和0.350 2;第3主分量可代表结实率,载荷量为0.595 5;第4主分量可代表成穗率、穗长和千粒质量,载荷量分别为-0.528 5、0.430 9和 0.285 4;这4个主分量中,仅第1、2、3主分量的特征值大于1,且累计贡献率为79.582 1%,说明第1、2、3主分量含有大部分信息,相对贡献较大。说明云南高海拔地区高原粳稻的总粒数、实粒数、株高、全生育期、最高苗、产量、基本苗、有效穗和结实率8个性状相对品种的综合表现影响更大,在8个性状中仅最高苗的载荷量为负值,其余均为正值,说明在云南中海拔地区高原粳稻育种时,应尽量选择多实多粒、株高偏高、分蘖适中且成穗高的高产品种。
3 讨论
中国人口众多且不断增长、在耕地面积不断减少的情况下,继续提高水稻总产只能依靠提高单产,品种改良成为最行之有效的手段[8]。如果将品种演变规律加以分析,对品种改良有着深远的意义。云南粳稻的高产育种始于20世纪50年代初,经历了3个阶段,第1阶段是20世纪50年代至20世纪60年代初,主要是评选推广农家良种;第2阶段是20世纪60年代中期至20世纪70年代,外引了一批植株较矮、株形优良的平原粳稻适种推广;第3阶段是20世纪80年代至20世纪90年代后期,此时期也是云南粳稻高产育种的黄金时代,侧重于大穗大粒稻种遗传资源的株型改良,也就是注重品种的穗数、粒数和千粒质量。由此可以看出,这3个阶段所对应的品种更新换代都是以提高产量为基础,以解决水稻生产中面临的问题,但这些选种目标都过于笼统。
由于云南省的地理和气候条件比较多样,相关研究表明,云南省的选种工作,要选出适应不同环境条件的良种,特别是希望在少数几个选种的中心,能够照顾到不同地区的需要,不从生态观点出发,是很难建立一套适用的工作方法[9-15]。本研究在云南省中海拔地区种植27个高原粳稻品种,高海拔地区种植13个高原粳稻品种,采集2017年云南省区域试验的12个性状田间表现数据,将水稻育种的传统评价方法与相关分析、通径分析、主成分分析及灰色关联度的分析相结合,得知与育种目标关联度较大的性状,在水稻品种选育过程中,品系(组合)的优劣其实就是其综合性状的外在表现,采用此方法可以克服人为的主观因素,提高品种(组合)评价的准确性和育种效率。为明确主要农艺性状之间的关系,并给云南省高原粳稻的品种选育提供一定的理论依据,研究结果表明,此方法可以快速计算出每个材料的综合评分和排名,有助于提高育种材料的筛选效率。所建立方法的评价结果与品种区域试验意见趋于一致,证明此方法能有效筛选出优良品种,接下来的工作可以通过此法分析了解各主要性状对产量影响的相对重要性,对品种的各性状进行综合评估,把田间的表现型与室内考种数据结合起来,使人们对后代的选择更加合理,有效提高育种材料的筛选效率。
研究证明此法在云南省品种选育中的运用是行之有效的。结合各组合性状的相关分析,说明在云南省高原稻区选育适宜云南省中海拔地区的常规粳稻品种时应注重选择生育期偏长、株高偏高及千粒质量与有效穗适中的大穗多粒型品种,而在云南省高原稻区选育适宜云南省高海拔地区的常规粳稻品种时应注重选择生育期适中、株高偏高、成穗率高及千粒质量与有效穗适中的大穗多粒型品种的品种类型,并注意性状间的协调发展,以充分表现品种潜力,选育出优异的高原粳稻品种。
参考文献:
[1]Peng S B,Tang Q Y,Zou Y B. Current status and challenges of rice production in China[J]. Plant Production Science,2009,12(1):3-8.
[2]王宏广. 中国粮食安全研究[M]. 北京:中国农业出版社,2005.
[3]张洪程,张 军,龚金龙,等. “籼改粳”的生产优势及其形成机理[J]. 中国农业科学,2013,46(4):686-704.
[4]秦志列,王 术,王伯伦. 不同穗型水稻产量形成及物质生产分析[J]. 中国农学通报,2006,22(4):181-184.
[5]吴伟明,程式华. 水稻根系育种的意义与前景[J]. 作物育种信息,2005,19(8):27-28.
[6]程式華,曹立勇,陈深广,等. 后期功能型超级杂交稻的概念及生物学意义[J]. 中国水稻科学,2005,19(3):280-284.
[7]张 旭. 水稻生态育种[M]. 北京:农业出版社,1991.
[8]吴文革,张洪程,吴桂成,等. 超级稻群体籽粒库容特征的初步研究[J]. 中国农业科学,2007,40(2):250-257.
[9]张 耗,谈桂露,薛亚光,等. 江苏省粳稻品种近60年演进过程中产量与形态生理特征的变化[J]. 作物学报,2010,36(1):133-140.
[10]Kim D M,Lee H S,Kwon S J,et al. High-density mapping of quantitative trait loci for grain-weight and spikelet number in rice[J]. Rice,2014,7(1):14.
[11]Zhuang J Y,Fan Y Y,Rao Z M,et al. Analysis on additive effects and additive-by-additive epistatic effects of QTLs for yield traits in a recombinant inbred line population of rice[J]. Theoretical and Applied Genetics,2002,105(8):1137-1145.
[12]钟 楚,朱颖墨,朱 勇,等. 云南不同类型一季稻产量形成及其与气象因子的关系[J]. 应用生态学报,2013,24(10):2831-2842.
[13]段 骅,杨建昌. 高温对水稻的影响及其机制的研究进展[J]. 中国水稻科学,2012,26(4):393-400.
[14]刘向蕊,陈小荣,杨 军,等. 冷害隶属函数在水稻耐低温性状的评价[J]. 江西农业大学学报,2013,35(4):675-681.
[15]刘奇华,周学标,杨连群,等. 生育前期遮光对水稻灌浆期剑叶生理特性及籽粒生长的影响[J]. 应用生态学报,2009,20(9):2135-2141.
