Stability of motor endplates is greater in the biceps than in the interossei in a rat model of obstetric brachial plexus palsy
2020-03-07BoLiLiangChenYuDongGu
Bo Li , Liang Chen , , Yu-Dong Gu
1 Department of Hand Surgery, Huashan Hospital and Institutes of Biomedical Sciences, Fudan University, Shanghai, China
2 Shanghai Key Laboratory of Peripheral Nerve and Microsurgery, Shanghai, China
Abstract The time window for repair of the lower trunk is shorter than that of the upper trunk in patients with obstetric brachial plexus palsy. The denervated intrinsic muscles of the hand become irreversibly atrophic much faster than the denervated biceps. However, it is unclear whether the motor endplates of the denervated interosseous muscles degenerate more rapidly than those of the denervated biceps. In this study, we used a rat model of obstetric brachial plexus palsy of the right upper limb. C5-6 was lacerated distal to the intervertebral foramina, with concurrent avulsion of C7-8 and T1, with the left upper limb used as the control. Bilateral interossei and biceps were collected at 5 and 7 weeks. Immunof luorescence was used to assess the morphology of the motor endplates. Real-time quantitative polymerase chain reaction and western blot assay were used to assess mRNA and protein expression levels of acetylcholine receptor subunits (α, β and δ), rapsyn and β-catenin. Immunof luorescence microscopy showed that motor endplates in the denervated interossei were fragmented, while those in the denervated biceps were morphologically intact with little fragmentation. The number and area of motor endplates, relative to the control side, were significantly lower in the denervated interossei compared with the denervated biceps. mRNA and protein expression levels of acetylcholine receptor subunits (α, β and δ) were significantly lower, whereas β-catenin protein expression was higher, in the denervated interossei compared with the denervated biceps. The protein expression of rapsyn was higher in the denervated biceps than in the denervated interossei at 7 weeks. Our findings demonstrate that motor endplates of interossei are destabilized, whereas those of the biceps remain stable, in the rat model of obstetric brachial plexus palsy. All procedures were approved by the Experimental Animal Ethics Committee of Fudan University, China (approval No. DF-187) in January 2016.
Key Words: acetylcholine receptor subunits; biceps; interossei; motor endplates; nerve regeneration; obstetric brachial plexus palsy; peripheral nerve injury
Introduction
Obstetric brachial plexus palsy (OBPP) is a stretch injury of the brachial plexus during delivery (Pondaag et al., 2004; Barsaoui et al., 2017). Approximately 70% of patients with OBPP spontaneously recover by 4-6 months postnatally (Malessy and Pondaag, 2009; Shah et al., 2019); the remainder experience various sequelae of the upper limbs, necessitating reconstruction surgery. For nerve surgery, a longer operative delay is unfavorable for functional recovery (Mackinnon, 1989; Laurent et al., 1993). For OBPP, the clinical consensus is that the C8-T1 spinal nerves innervating the intrinsic muscles of the hand (IMH) should be reconstructed within a maximum of 3 months postnatally (Chuang et al., 2005). In contrast, for the C5-C6-innervated muscles of the upper arm, nerve grafting up to 30 months after birth might still provide functional improvement (Boom et al., 2000). In our previous study of a rat model of OBPP, in which the spinal nerves were reconstructed, we showed that atrophy of the intrinsic muscles of the forepaw (IMF) became irreversible far earlier than atrophy of the biceps, resulting in a shortened operative time window for nerve reconstruction of C8-T1 compared with C5-6 (Wu et al., 2013). Further studies using a rat model of OBPP without nerve reconstruction, in which the IMF are irreversibly atrophied and the biceps are reversibly atrophied, demonstrated that the denervated IMF and the denervated biceps show a unique expression profile of miRNAs (Pan et al., 2015) and mRNAs (Wu et al., 2016), respectively, and that the denervated IMF lacked self-repair potential compared with the denervated biceps (Wu et al., 2016). Furthermore, these findings suggest that the dysregulated miRNAs and mRNAs in these muscles might be associated with the structural and functional changes in the neuromuscular junctions (NMJs) after injury.
The NMJ is composed of the following five components: terminal Schwann cells, nerve endings that release neurotransmitters, a synaptic cleft, a postsynaptic membrane specialization containing the nicotinic acetylcholine receptor (AChR), and a sarcoplasm that provides structural and metabolic support for the postsynaptic region (Sanes and Lichtman, 2001; Wu et al., 2010; Mukund and Subramaniam, 2020). When the peripheral nerve is disconnected, its distal portion undergoes progressive (Wallerian) degeneration, characterized by a breakdown of axons and myelin, which peaks 4-6 weeks post-injury (Beirowski et al., 2004; Mu et al., 2018).
In rats with sciatic nerve transection, Adams et al. (1995) found that AChR mRNA expression in the gastrocnemius and tibialis anterior muscles peaked 7 days after denervation, and decreased to 10% of the control 6 months later. Ijkema-Paassen et al. (2002) showed that in rats with sciatic nerve transection, a few motor endplate fragments are detectable in the tibialis anterior muscle 4 weeks post-injury, but none by 7 weeks. After denervation, old AChRs are removed faster and replaced by new ones more slowly, while in innervated NMJs, the recycling of AChRs is in balance (Bruneau and Akaaboune, 2006). A recent study showed that early repair of sciatic nerve transection in mice and rats enhances functional reconstruction of NMJs in the target muscles, whereas delayed repair (beyond 1 month post-injury) only provides morphological reconstruction of NMJs without functional improvement (Sakuma et al., 2016).
Given that the IMH have a far shorter operative window than the biceps in OBPP, we hypothesized that degenerative changes occur more rapidly in the motor endplates in the IMH than in the biceps, resulting in the quicker irreversible atrophy of the denervated IMH compared with the biceps. In the present study, we tested this hypothesis using a rat model of OBPP in which the IMF are irreversibly atrophied, but atrophy of the biceps remains reversible (Pan et al., 2015; Wu et al., 2016).
Materials and Methods
Production of the rat model of OBPP and specimen collection
All procedures were approved by the Experimental Animal Ethics Committee of Fudan University, China (approval No. DF-187) in January 2016. Thirty-six 7-day-old male Sprague-Dawley rats weighing 11-13 g were provided by the Animal Science Department, Fudan University, China (license No. SYXK (Hu) 2014-0029).
Each rat was anesthetized intraperitoneally with 1% pentobarbital sodium (Sigma-Aldrich, St. Louis, MO, USA). While in the supine position, the right spinal nerves (C5-T1) were exposed via the supraclavicular route. A common type of OBPP was created. Brief ly, C5-6 was lacerated distal to the intervertebral foramina, with concurrent avulsion of C7-T1, with no possibility of proximal nerve regeneration. To prevent nerve regeneration from C5-6, their proximal stumps were embedded into the surrounding tissue. The brachial plexus on the left remained uninjured to serve as control. All rats were caged under standard room temperature and humidity, with a 12-hour dark-light cycle and ad libitum access to food and fresh water.
The rats were sacrificed, and the bilateral interossei, one of the major IMF, and biceps were collected at 5 and 7 weeks after surgery (18 rats at each time point). This timeframe was selected according to our previous study of the OBPP rat model with nerve grafting, which showed that at 5 weeks after denervation, the IMF became irreversibly atrophic, whereas the biceps remained in a state of reversible atrophy (Wu et al., 2013). The criteria for successful modeling of OBPP were the following: (1) At 5 and 7 weeks after surgery, the right upper limb was thinner than the left upper limb; and (2) it had no function. Harvested specimens were stored at -80°C until analysis. Of the 18 rats analyzed at each time point, the interossei and biceps from 6 rats were used for immunofluorescence analysis of morphological changes in the motor endplates. Samples from another 6 rats were used to assess mRNA expression of AChRα, AChRβ and AChRδ subunits, as well as rapsyn and β-catenin, by real-time quantitative polymerase chain reaction (RT-qPCR). Samples from the remaining 6 rats were used to evaluate protein levels of these NMJ components by western blot assay.
Immunof luorescence labeling
Immunofluorescence labeling was performed to reveal the structure of NMJs at 5 and 7 weeks after surgery. AChR staining was used to assess the morphology of motor endplates, particularly as the postsynaptic muscle membrane is enriched in AChRs (Basu et al., 2015). Neurofilament staining was performed to show the motor nerve endings. The muscle tissue specimens were fixed overnight in 4% paraformaldehyde, dehydrated in 30% sucrose for 12 hours, and embedded in optimal cutting temperature compound. The tissue was sliced longitudinally using a microtome (Leica, Wetzlar, Germany), with 10 sections (20-μm thickness) harvested for morphological analysis of motor endplates. Sections were permeabilized with 0.3% Triton X-100 in phosphate-buffered saline for 30 minutes, and blocked with 3% bovine serum albumin in phosphate-buffered saline for 1 hour at room temperature. The sections were then incubated with rabbit anti-neurof ilament heavy chain antibody (1:1000; Ab8135; Abcam, Cambridge, MA, USA) overnight at 4°C, followed by incubation with donkey polyclonal anti-rabbit IgG H&L (Alexa Fluor 647) (1:200; Ab150075; Abcam) (for neurof ilament heavy chain) and α-Bungarotoxin-ATTO-488 (1:100; ALO-B-100-AG-0.1; Alomone, Jerusalem, Israel) (for AChRs) for 2 hours at 4°C. Finally, sections were mounted on slides with antifade reagents. Confocal f luorescence images were obtained on a Leica TCS SP8 microscope (Leica, Solms, Germany). Projections of maximal pixel intensity of Z-stacks were acquired with LAS-AF software (Leica).
The motor endplates for one section were counted, regardless of the orientation of motor endplates in relation to the focus plane of a microscope. For each muscle block, the number of motor endplates was evaluated semiquantitatively, and was expressed as the mean number of motor plates per section, which was calculated from f ive random sections from a total of 10. The percentage of endplates in the denervated muscle was calculated by dividing this value by that in the contralateral normal counterpart and multiplying by 100%. The area of motor endplates was measured using ImageJ software (NIH, Bethesda, MD, USA). The mean area of each of 50 endplates (10 chosen randomly from each of f ive sections) was calculated for the muscle tissue block. The percentage mean area of endplates in the denervated muscle was calculated by dividing the area by that in the contralateral normal muscle and multiplying by 100%.
RT-qPCR
AChR subunits (α, β and δ) are the main components of motor endplates, and rapsyn and β-catenin are key regulators of these subunits. cDNA templates were synthesized from 1 μg of RNA from each muscle sample at 5 and 7 weeks after surgery using SuperScript III Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA). mRNA expression levels were determined by RT-qPCR using ABI SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) and a CFX96 thermocycler (Bio-Rad, Hercules, CA, USA). Primer pairs, synthesized by Sangon Biotech (Shanghai, China), are listed in Table 1. Relative mRNA expression levels were assessed for candidate genes from both denervated and contralateral normal muscles using the 2-ΔΔCTmethod and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as the internal control for normalization. The percentage expression of AChRα, AChRβ, AChRδ, rapsyn and β-catenin in the denervated muscle was calculated by dividing expression by the corresponding expression in the contralateral normal muscle and multiplying by 100%.
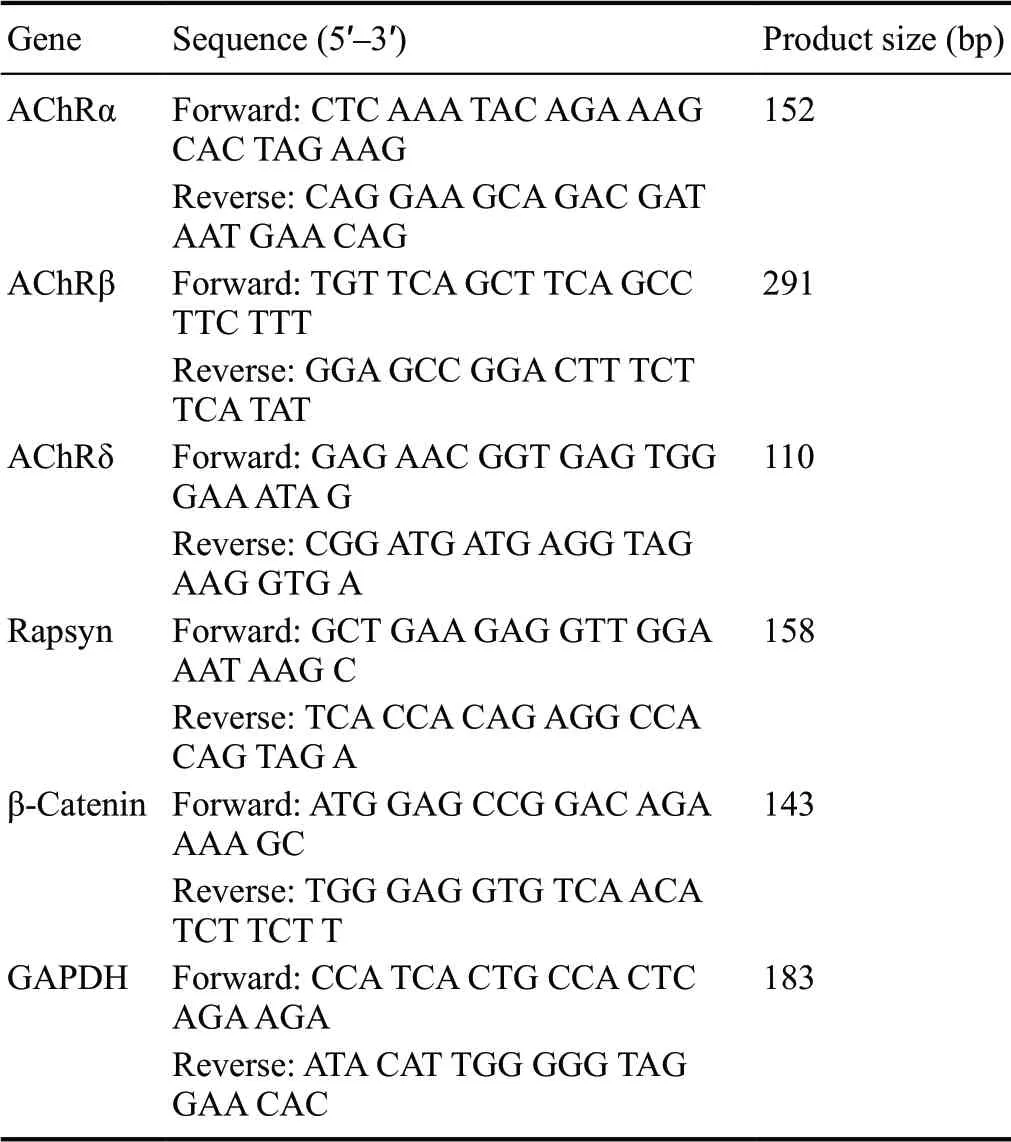
Table 1 Sequences of the primers used in the study
Western blot assay
Proteins were extracted from the bilateral interossei and biceps at 5 and 7 weeks after surgery at 4°C in radioimmunoprecipitation assay buffer containing protease and phosphatase inhibitors. Following centrifugation, the protein concentration in the supernatants was quantif ied using a bicinchoninic acid assay kit (Thermo Scientific, Waltham, MA, USA). For each sample, 30 μg of total protein was separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and transferred to a polyvinylidene f luoride membrane. The membrane was blocked with 5% dried skim milk and incubated at 4°C for 12 hours with one of the following primary antibodies: rabbit anti-AChRα (1:1000; PAB12331; Abnova, Walnut, CA, USA), rabbit anti-AChRβ (1:1000; ab76159; Abcam), mouse anti-AChRδ (1:5000; NB120-2804; Novus Biologicals, Littleton, CO, USA), rabbit anti-rapsyn (1:1000; ab156002; Abcam), rabbit anti-β-catenin (1:5000; ab32572; Abcam) or rabbit anti-GAPDH (1:2500; ab9485; Abcam). After three washes with Tris-buffered saline-Tween for 15 minutes each, blots were incubated at room temperature for 2 hours with the corresponding horseradish peroxidase-conjugated secondary antibody (goat anti-mouse or goat anti-rabbit; 1:5000; Millipore, Burlington, MA, USA). After three washes with Tris-buffered saline-Tween for 15 minutes each, immunoreactive bands were detected by chemiluminescence (Thermo Scientific) and quantified using ImageJ software, with GAPDH as an internal control. Finally, the percentage expression of AChRα, AChRβ, AChRδ, rapsyn and β-catenin in the denervated muscle was calculated by dividing the expression level by the corresponding expression level in the contralateral normal muscle and multiplying by 100%.
Statistical analysis
The median (p25-p75) was recorded for expression data. The Mann-Whitney U-test was used to measure differences between denervated interossei and denervated biceps using SPSS 19.0 software (SPSS, Armonk, NY, USA). P < 0.05 was considered statistically significant.
Results
Morphological and quantitative evaluation of motor endplates
Immunof luorescence staining for AChRs in muscle sections revealed that in normal interossei and biceps (Figure 1A1-A3 and C1-C3), the motor endplate, a pretzel-like structure, was located in the center of the muscle fiber, and the motor nerve endings, with neurof ilament labeling, were localized around the motor endplate. At 5 weeks after denervation, no motor nerve endings could be detected in interossei or biceps. In denervated interossei (Figure 1B1-B3), AChR clusters at the postsynaptic membrane were fragmented, and their contours were difficult to identify. However, in denervated biceps (Figure 1D1-D3), AChR clusters were morphologically intact with little fragmentation, although they were shrunken. In interossei and biceps that had been denervated for 7 weeks, fragmentation of the interossei motor endplates worsened (Figure 2B1-B3), while in comparison, the biceps motor endplates remained mostly intact (Figure 2D1-D3).
Compared with the normal side, the number and area of the motor endplates were markedly reduced in the denervated interossei and biceps. The percentage mean number of endplates per section relative to the contralateral normal counterpart, was significantly lower in the denervated interossei than in the denervated biceps, 5 weeks (P < 0.01) and 7 weeks (P < 0.01) after denervation (Table 2). Furthermore, the percentage mean area per endplate relative to the contralateral normal counterpart was significantly lower in the denervated interossei than in the denervated biceps, 5 weeks (P < 0.01) and 7 weeks (P < 0.01) after denervation (Table 2).
mRNA expression levels of AChRα, AChRβ, AChRδ, rapsyn and β-catenin
The expression levels of AChRα, AChRβ and AChRδ mRNA (relative to the corresponding normal control) were significantly lower in denervated interossei than in denervated biceps, 5 weeks (P < 0.01) and 7 weeks (P < 0.01) after denervation. The expression of rapsyn mRNA was not significantly different between the two denervated muscles at 5 weeks (P > 0.05) or 7 weeks (P > 0.05). The expression of β-catenin mRNA was significantly higher in denervated interossei compared with denervated biceps at 5 weeks (P < 0.01), but not at 7 weeks (P > 0.05) (Figure 3).
Protein expression levels of AChRα, AChRβ, AChRδ, rapsyn and β-catenin
The expression levels of AChRα, AChRβ and AChRδ, relative to the contralateral normal muscles, were significantly lower in the denervated interossei than in the denervated biceps at 5 weeks (P < 0.01 or P < 0.05) and 7 weeks (P < 0.01) after denervation. The expression of rapsyn protein, relative to the corresponding normal control, was significantly lower in the denervated interossei than in the denervated biceps at 5 weeks (P < 0.01), but not at 7 weeks (P > 0.05). The expression of β-catenin protein, relative to the normal control, was significantly higher in the denervated interossei than in the denervated biceps at 5 weeks (P < 0.01) and 7 weeks (P < 0.01) (Figure 4).
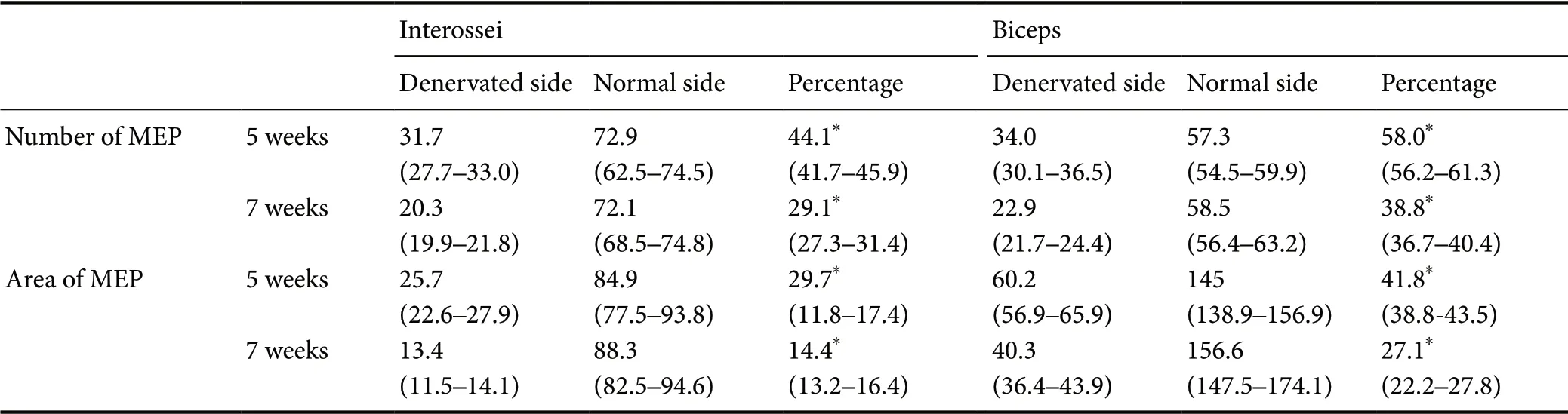
Table 2 Comparison of percentage of number and area of motor endplates between denervated interossei and denervated biceps in rats
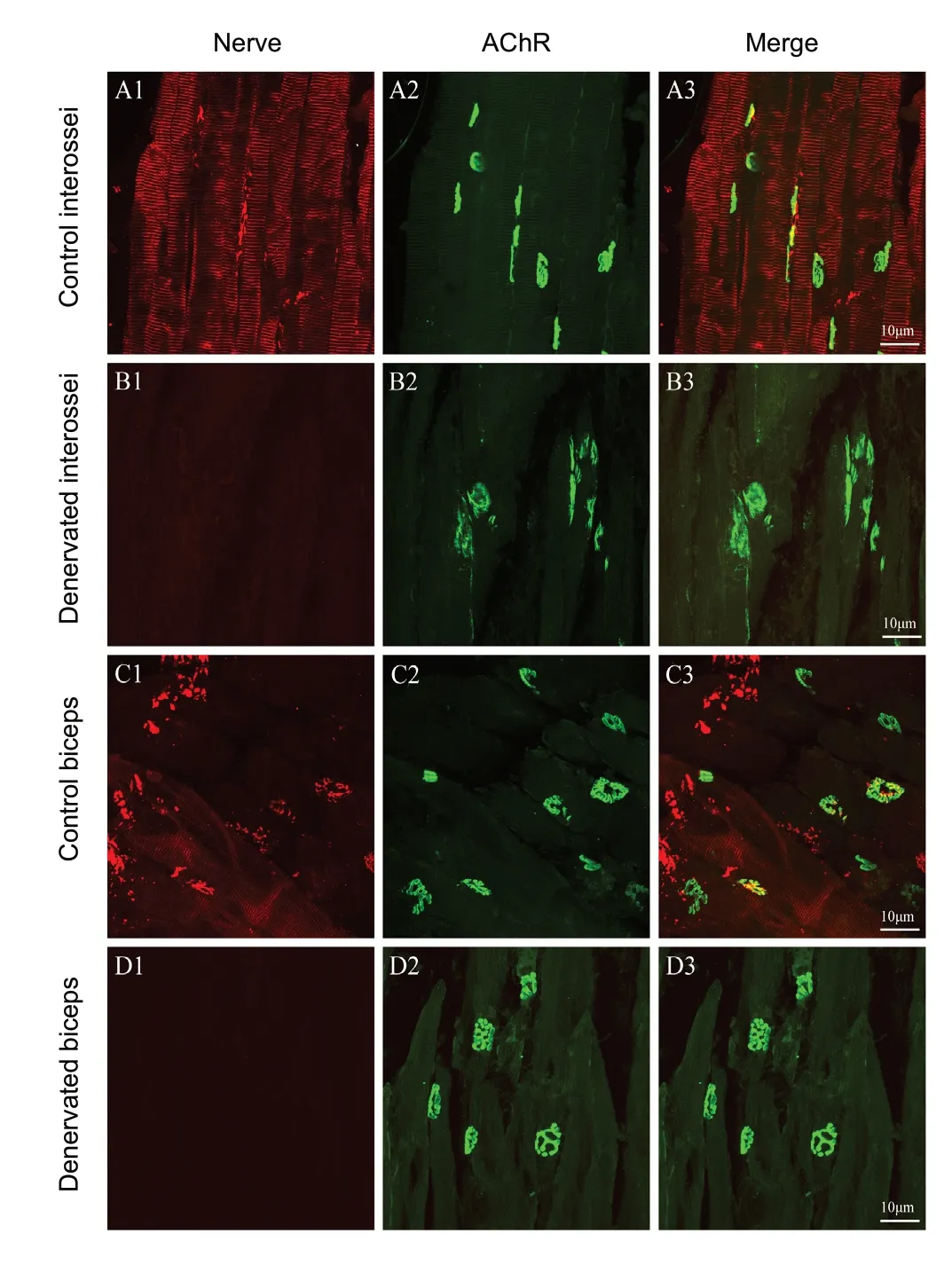
Figure 1 Confocal images of motor endplates of interossei and biceps 5 weeks after denervation.

Figure 2 Confocal images of motor endplates of interossei and biceps 7 weeks after denervation.
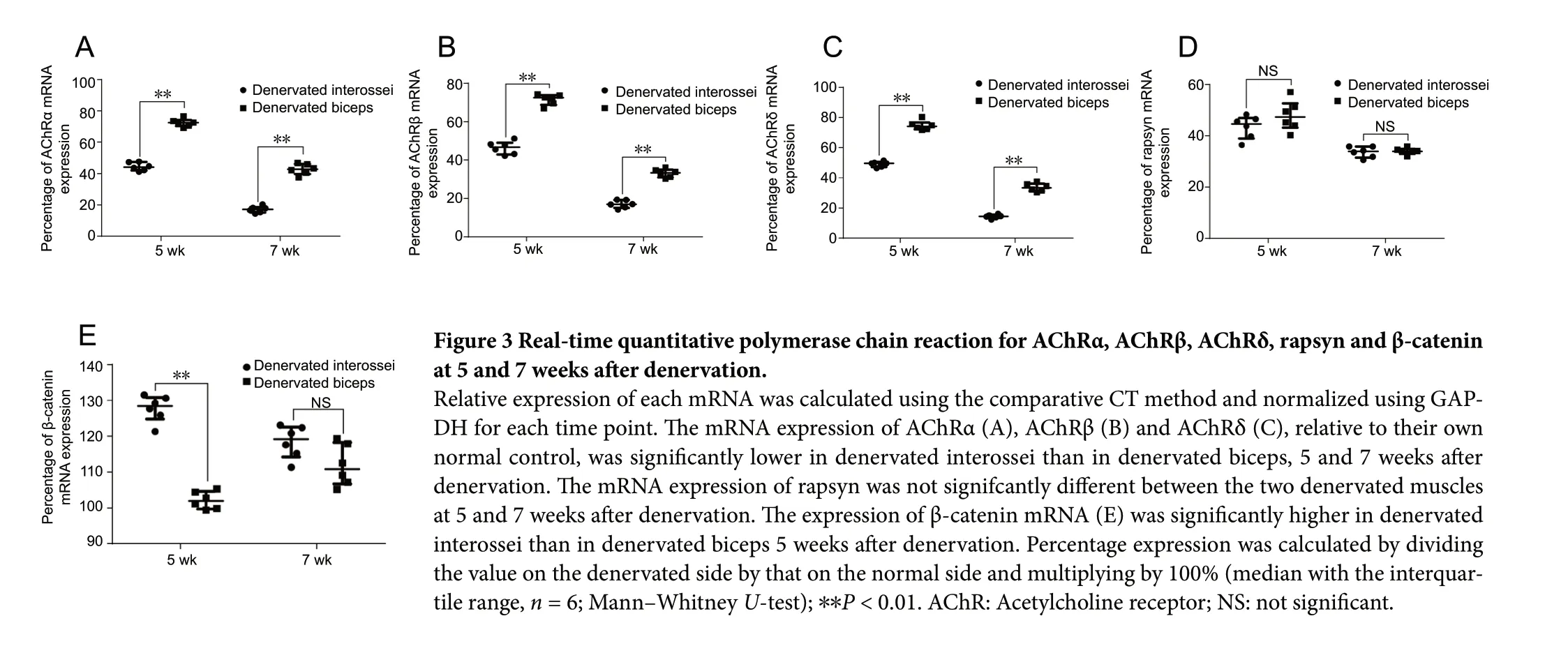
Figure 3 Real-time quantitative polymerase chain reaction for AChRα, AChRβ, AChRδ, rapsyn and β-catenin at 5 and 7 weeks after denervation.
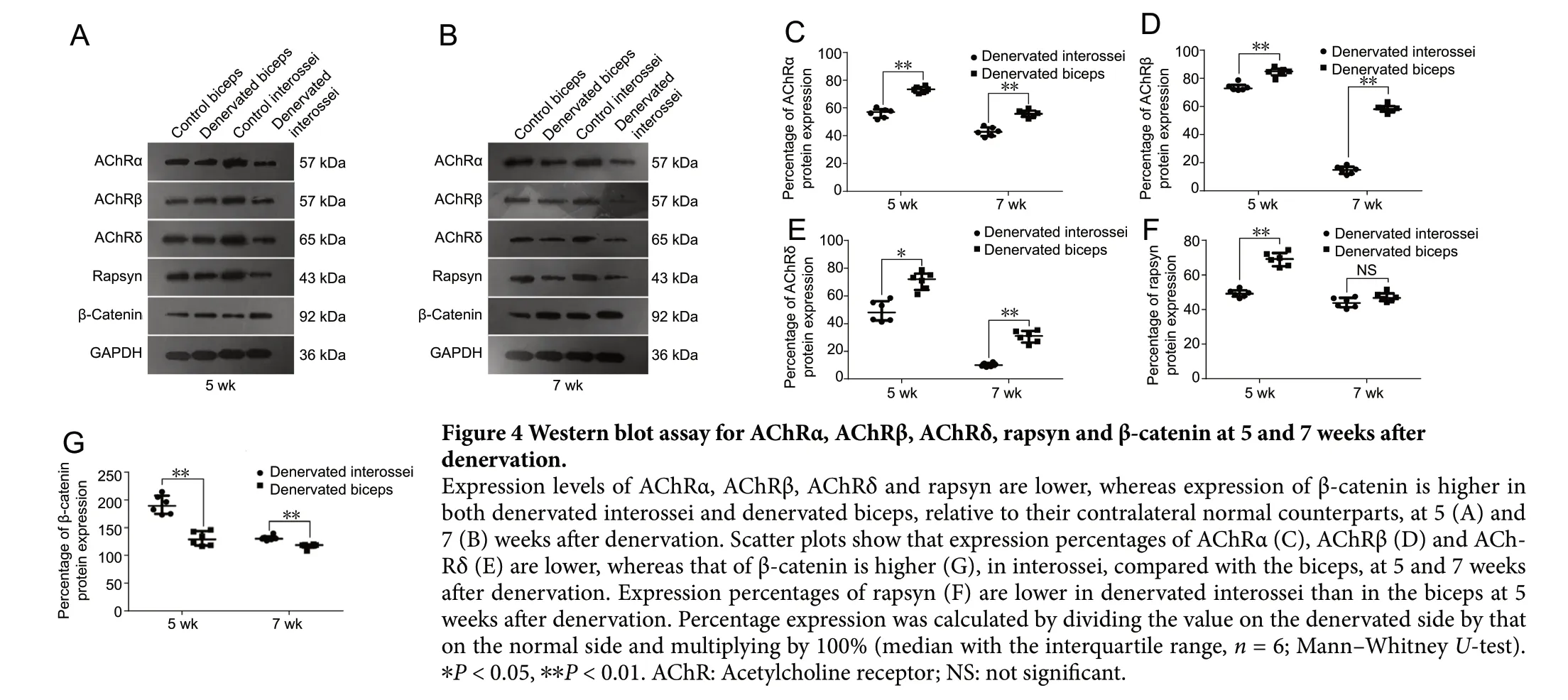
Figure 4 Western blot assay for AChRα, AChRβ, AChRδ, rapsyn and β-catenin at 5 and 7 weeks after denervation.
Discussion
At the early stage after denervation, motor endplates remain intact anatomically because of their intrinsic ability to prepare for re-innervation. For example, in mice with transection of the sciatic nerve, motor endplates in the target muscles can remain morphologically intact 1 month after denervation, although degenerative changes appear at 2 months (Kurimoto et al., 2015). In the present study, the motor endplates were imaged by immunof luorescence with α-bungarotoxin, which binds AChRs. It was previously reported that AChRs may diffuse throughout the entire muscle fiber after denervation (Miledi and Slater, 1968; Stanley and Drachman, 1981), and that the extra-junctional AChRs are diffusely distributed and no longer localized to motor endplates (Hartzell and Fambrough, 1972), whereas regenerating axons always reoccupy the original synaptic sites (Purves and Lichtman, 1987; Kang and Lichtman, 2013). Here, we found that the motor endplates of the denervated biceps displayed an intact morphology, while those in the denervated interossei appeared fragmented. This suggests that AChRs in the biceps remain localized to their original synaptic sites. Furthermore, the number and area of the motor endplates, relative to the uninjured side, were much lower in the denervated interossei than in the denervated biceps. Thus, together with our previous study, the current findings suggest that the motor endplates of the denervated IMH exhibit a lower ability for self-repair compared with the denervated biceps, resulting in a relatively reduced potential for re-innervation.
The AChR in motor endplates of skeletal muscles is a pentamer composed of 2α, β, γ and δ subunits (Witzemann et al., 1989; Adams et al., 1995; Foucault-Fruchard et al., 2018; Weng et al., 2018; Das et al., 2019). In innervated muscles, the expression of the AChR subunits may be reduced by neuronal factors and muscular activity induced by nerve f iring, in a negative feedback effect (Witzemann et al., 1991). In rats with nerve transection, denervation results in upregulation of most AChR subunits in the target muscle, peaking at 7 days post-injury, possibly by interrupting the negative feedback. However, the AChR subunits are downregulated below baseline from one month onwards, likely because the progressive atrophy of the denervated muscles may reduce the availability of AChR subunits (Ma et al., 2005, 2007; Shen et al., 2006). In the present study, using rats at 7 days of age, we only examined the expression of AChRα, β and δ subunits, because they are expressed more stable than the γ and δ subunits shortly after birth (Witzemann et al., 1989). The α, β and δ subunits were downregulated in both the interossei and biceps at 5 and 7 weeks after denervation, consistent with previous studies (Adams et al., 1995; Ma et al., 2007). The relative mRNA and protein expression levels of the α, β and δ subunits were reduced, while β-catenin protein expression was increased in the denervated interossei, compared with the denervated biceps, at 5 and 7 weeks. Rapsyn, a synaptic protein, is necessary for the clustering of AChRs and the formation of NMJs (Apel et al., 1997; Phillips et al., 1997; Huebsch and Maimone, 2003; Chen et al., 2016). β-Catenin is a key signaling molecule in the Wnt pathway. Activation of β-catenin induces its aggregation in the cell membrane and cytoplasm, as well as its translocation to the nucleus to activate various target genes (Cisternas et al., 2014). A study of mice with sciatic nerve laceration showed that β-catenin mRNA was upregulated around ACh-Rs in denervated target muscles (Kurimoto et al., 2015) and inhibited AChR clustering by suppressing rapsyn expression (Wang et al., 2008). Our current findings suggest that in denervated interossei, the AChR is downregulated because of a lack of subunit availability, compared with the denervated biceps. Furthermore, the subunits are less likely to combine to form active receptors in denervated interossei, compared with denervated biceps. These changes likely contribute to the destabilization of endplates in the denervated interossei.
In our previous study using the OBPP rat model, in which the proximal stump of the C6 spinal nerve was grafted to the ulnar nerve and that of C5 to the musculocutaneous nerve, we found that the IMF remained uninnervated at 5 weeks, while the biceps were reinnervated even at 10 weeks (Wu et al., 2013). Accordingly, regenerated motor endplates were much fewer in the IMF than in the biceps.
In a study using a mouse model of motor neuron disease, NMJs of IIb (fast-twitch) fibers were found to be lost more easily than NMJs of the IIa (fast-twitch) and I (slow-twitch) fibers, and botulinum toxin A-induced plasticity was much poorer in IIb fibers compared with the other fibers (Frey et al., 2000). Lumbrical muscles, one of the major muscles of the IMF, whose function is very similar to that of the interossei, are almost all composed of type II fibers (Russell et al., 2015), of which IIb fibers account for 80% of the total (Ridge and Rowlerson, 1996). In contrast, in the biceps, the type I and IIa fibers account for up to 55% of total muscle fibers (Fuentes et al., 1998). Thus, the much greater ratio of IIb fibers in the interossei might be the major factor contributing to the comparatively rapid degeneration of motor endplates in the interossei after denervation. Functionally, the IMH are still immature a year postnatally, whereas the biceps is mature at birth (Zafeiriou, 2004). Immature synapses during development are more vulnerable to damage caused by the blocking of nerve-evoked activities, compared with mature synapses (Pun et al., 2002). We conjecture that the relatively immature status of the IMH may also contribute to the comparatively rapid motor endplate degeneration and the irreversible atrophy in these muscles in OBPP.
In summary, the motor endplates of denervated interossei were rapidly destabilized, while those of the denervated biceps remained comparatively stable, in a rat model of OBPP. AChR mRNA and protein expression levels were significantly lower, whereas β-catenin protein expression was higher, in the denervated interossei compared with the denervated biceps. These findings suggest that in OBPP, the self-repair potential of motor endplates is lower in the denervated IMH than in the denervated biceps.
Acknowledgments:We thank Li Yao from Howard Hughes Medical Institute (Bethesda, MD, USA) for her assistance with the OBPP model illustrations.
Author contributions:Study design: LC; experimental implementation: BL; data analysis and paper writing: BL and LC; analysis with constructive discussions: YDG. All authors approved the final version of the paper.
Conf licts of interest:The authors declare that there are no conf licts of interest associated with this manuscript.
Financial support:This study was supported by the National Natural Science Foundation of China, No. 81672240 (to LC). The funding source had no role in study conception and design, data analysis or interpretation, paper writing or deciding to submit this paper for publication.
Institutional review board statement:All procedures were approved by the Experimental Animal Ethics Committee of Fudan University, China (approval No. DF-187) in January 2016. The experimental procedure followed the United States National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1996).
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewer:Annas Al-Sharea, Baker IDI Heart and Diabetes Institute, Australia.
Additional file:Open peer review report 1.
杂志排行

中国神经再生研究(英文版)的其它文章
- Recovery of an injured ascending reticular activating system with recovery from a minimally conscious state to normal consciousness in a stroke patient: a diffusion tensor tractography study
- The role of vascularization in nerve regeneration of nerve graft
- New insights into Wnt signaling alterations in amyotrophic lateral sclerosis: a potential therapeutic target?
- Advanced diffusion magnetic resonance imaging in patients with Alzheimer’s and Parkinson’s diseases
- Modulation of autophagy for neuroprotection and functional recovery in traumatic spinal cord injury
- Decoding epigenetic codes: new frontiers in exploring recovery from spinal cord injury