扰动方式对湖泊营养盐及藻类生物量影响的模拟研究
2020-03-07赵家敏杨桂军张宏亮汤祥明
赵家敏,田 伟,杨桂军,张宏亮,汤祥明
(1.中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,江苏 南京 210008;2.安阳师范学院资源环境与旅游学院,河南 安阳 455000;3.安徽师范大学环境科学与工程学院,安徽 芜湖 241003;4.江南大学环境与土木工程学院,江苏 无锡 214122)
20世纪以来,伴随工业化进程的发展及城市化进程的加快,湖泊富营养化日趋严重[1-2]。蓝藻水华的暴发不仅会影响供水危机,死亡后的蓝藻还会消耗大量的氧气,从而促使水中生物窒息死亡,威胁着水环境生态系统的完整性和可持续性[3]。
富营养化水体中蓝藻水华的暴发与蓝藻本身的生理特征有关,如伪空泡、固碳、光吸收及营养盐利用能力,使其在藻类竞争中保持优势[4-5]。在蓝藻细胞生长阶段,营养盐、温度、光照等环境因素对蓝藻生物量影响较为显著,为蓝藻水华暴发奠定了物质基础;蓝藻水华暴发阶段主要受蓝藻细胞(团)浮力作用与水动力湍流作用的共同影响[6-7]。太湖受亚热带季风影响,特别是夏季和秋季,湖水在风浪扰动的作用下始终处于一个动态环境。蓝藻水华在太湖中的发生往往是伴随着一场大风过程结束之后出现的[8], 因此水动力在蓝藻水华形成过程中起着重要作用。适当的风浪扰动会使太湖微囊藻小群体碰撞聚集形成大群体,从而获得较快的上浮速度,待扰动停止时漂浮于水面上[9]。扰动条件可以在一定程度上影响营养盐的分布, 避免营养盐在环境中出现分布不均现象[10]。
目前已有关于扰动强度对湖泊营养盐及藻类生长影响的研究[11],但是太湖中由风浪引起的扰动通常属于间歇扰动,而不是持续性扰动,表明水华暴发与扰动方式之间可能存在着某种联系。而国内外鲜见关于扰动方式对水体营养盐和蓝藻水华影响的研究。为了解不同扰动方式对水体营养盐及蓝藻水华的影响,笔者对比了不扰动、间歇扰动和持续扰动3种条件下水体氮、磷营养盐和叶绿素a(Chl-a)浓度的变化规律,以期为蓝藻水华的发生及控制提供理论依据。
1 材料与方法
1.1 实验设置与样品采集
1.1.1实验设置
实验为半野外大桶实验,在中国科学院太湖湖泊生态系统研究站生态室水池内进行。以9只用纯水洗净晾干的100 L有盖塑料水桶(上口截面积为0.163 m2,桶高约0.62 m)为实验容器,向每只水桶内抽入约100 L太湖水。在太湖湖边打捞发生水华时的蓝藻,蓝藻藻浆用64 μm孔径尼龙网浓缩(经鉴定,蓝藻的比例占85%以上,其中微囊藻占80%以上),添加适量蓝藻藻浆于上述水桶内,混匀后测定水桶中Chl-a浓度,保持各水桶内ρ(Chl-a)初始值为110 μg·L-1左右。因桶中没有添加沉积物,为保证实验期间水体中营养盐充足,向水桶中添加氮磷营养盐,其中TN和TP分别用NaNO3和K2HPO4·3H2O配制,保持各水桶内水的ρ(TN)为10 mg·L-1,ρ(TP)为0.5 mg·L-1,这是2000—2008年太湖梅梁湾TN和TP浓度平均值的4倍。水体静置1 d后开始实验。在6只水桶的水面下10 cm处固定变频造浪泵(中山市捷宝电子电器有限公司生产,型号为WP-60,功率为60 W),使用W1经典造浪模式,使扰动时波浪约5 cm高,接近太湖平均波高[12]。实验时3只水桶不做扰动(对照组),3只水桶每天扰动时间为24 h(持续扰动组),其余3只每天扰动6 h,时间为10:00—13:00以及16:00—19:00(间歇扰动组)。水桶悬挂在有机玻璃房内的大型水池内,实验时玻璃房内室温维持在29 ℃(夏季太湖平均水温)。实验持续19 d (2018年7月11—29日)。
1.1.2样品采集
实验开始第1、4、7、10、13、16、19天上午08:00用柱状采样器进行水柱均匀采样,采集约1 L水样,用于测定水体中的总氮(TN)、总磷(TP)、溶解性总磷(TDP)、溶解性总氮(TDN)、磷酸盐(PO43--P)、叶绿素a(Chl-a)、悬浮颗粒物(SS)、硝态氮(NO3--N)及氨氮(NH4+-N)。每天上午08:30用多参数水质检测仪(YSI)在水深约20 cm处现场测定水体的水温(T)、溶解氧(DO)和pH值等水质参数。
1.2 水质分析方法
Chl-a浓度参照陈宇炜等[13]的热乙醇提取、分光光度法测定。测定Chl-a时将水样过滤(混纤微孔滤膜,孔径0.45 μm)后,滤膜避光冷冻保存 24 h 以上。在暗光条件下用一定量w=90%的热乙醇研磨提取滤膜上的色素,再用GF/C玻璃纤维滤膜过滤提取液,采用分光光度法测定。
测定TN和TP浓度时用过硫酸钾氧化及消解水样,测定TDN、TDP浓度时先将水样经GF/F滤膜过滤后再进行硫酸钾氧化及消解。TN和TDN浓度采用紫外分光光度法测定, TP和TDP浓度采用钼锑抗显色分光光度法测定。NH4+-N浓度用纳氏试剂光度法测定,NO3--N和PO43--P浓度采用流动分析、分光光度法进行测定。SS采用GF/C玻璃纤维滤膜(0.45 μm)过滤后,105 ℃烘干30 min称重而得[14-15]。
1.3 数据分析
实验数据通过Microsoft Excel 2013录入,分别采用RStudio 3.5.2和Origin 2018软件分析和作图。利用R软件平台的Performance Analytics包进行理化因子的可视化相关性分析,采用非参数Kruskal-Wallis秩和检验分析不同组别之间是否存在显著性差异。
2 结果与分析
2.1 水体中物理因子的变化
实验期间各组水体中DO浓度均有减小的趋势(图1)。实验结束时,对照组、间歇扰动组和持续扰动组的ρ(DO)分别从初始值的9.06、9.36、9.78 mg·L-1减少到7.86、3.17和7.49 mg·L-1。间歇扰动组的ρ(DO)在4 d内迅速下降至3.20 mg·L-1左右,此后保持稳定;而对照组的DO浓度在4 d内呈上升趋势。总体而言,间歇扰动组DO浓度显著低于对照组和持续扰动组(P<0.01)。
实验期间3组的水温变化趋势相同,在1~13 d水温变化比较平稳,呈先升高后降低的趋势。随后水温迅速升高,在第19天时对照组、间歇扰动组和持续扰动组水温分别增加到31.7、31.7和32.2 ℃。持续扰动组水温高于对照组和间歇扰动组,差异不显著(P> 0.05)。

箱体的上下线代表上下四分位数;箱中横线代表中位数;线段上下线代表最大值和最小值。**表示P<0.01。
2.2 水体中氮浓度的变化
实验过程中水体TN、TDN及NO3--N 浓度的变化趋势如图2所示。TN、TDN和NO3--N 浓度在3种处理下均有所减少。间歇扰动和持续扰动的TN浓度略低于对照组,其中间歇扰动的TN浓度减少得最多。对照组、间歇扰动组和持续扰动组ρ(TN)从实验前的11.94、11.66和11.7 4 mg·L-1分别减少到实验后的8.81、6.33和6.50 mg·L-1。对照组与间歇扰动组、持续扰动组之间存在显著性差异(P<0.05)。
实验期间持续扰动组的ρ(TDN)和ρ(NO3--N)低于对照组,而间歇扰动组在1~7 d略高于对照组。对照组、间歇扰动组、持续扰动组的ρ(TDN)从实验前的10.33、10.45、10.86 mg·L-1分别减少到实验后的6.61、5.29、4.43 mg·L-1,但3种处理之间TDN浓度差异不显著(P>0.05)。3组中的ρ(NO3--N)从实验前的9.88、9.64、9.68 mg·L-1分别降至实验后的4.28、1.64、2.58 mg·L-1,但3种处理之间NO3--N浓度差异也不显著(P>0.05)。
3种处理下的NH4+-N浓度在实验结束时均有增加,持续扰动组的NH4+-N浓度略高于其他2组。NH4+-N浓度变化可以分为3个阶段:第1阶段(1~7 d)对照组、间歇扰动组、持续扰动组的ρ(NH4+-N)分别从0.20、0.18、0.19 mg·L-1上升到0.31、0.37、0.39 mg·L-1;第2阶段分别降至0.21、0.19、0.27 mg·L-1(第10天),随后升至0.34、0.36、0.44 mg·L-1(第13天);第3阶段分别降至0.28、0.25、0.34 mg·L-1(第16天),随后升至0.38、0.28、0.32 mg·L-1(第19天)。总体而言,3种扰动方式之间NH4+-N浓度无显著性差异(P> 0.05)。

箱体的上下线代表上下四分位数;箱中横线代表中位数;线段上下线代表最大值和最小值。*表示P<0.05。
2.3 水体中磷浓度的变化
实验过程中水体TP和TDP浓度的变化趋势如图3所示。在整个实验过程中3组的TP和TDP浓度均在下降,其中1~10 d 的TP和TDP浓度下降很快,第10天后两者的下降速率均放缓。实验结束后对照组、间歇扰动组、持续扰动组的ρ(TP)分别从初始的0.44、0.44、0.43 mg·L-1降至0.17、0.10、0.13 mg·L-1,3种处理下TP浓度无显著性差异(P>0.05)。间歇扰动组的TDP浓度始终占优势,3种条件下的ρ(TDP)分别从实验前的0.158、0.186、0.166 mg·L-1降至实验第19天的0.03、0.033、0.028 mg·L-1,间歇扰动组与持续扰动组之间存在显著性差异(P<0.05)。
2.4 水体中悬浮物与叶绿素浓度的变化
实验过程中水体SS和Chl-a的变化趋势如图4所示。间歇扰动组的SS浓度始终低于其余2组,3组中的SS浓度在实验结束时均有所增加。实验开始时对照组和间歇扰动组ρ(SS)分别为27.9和20.7 mg·L-1,2组ρ(SS)在第7天均下降至最低,分别为12.5和20.7 mg·L-1,随后分别上升至52.4和31.4 mg·L-1(第19天)。而持续扰动组ρ(SS)在实验期间从20.5 mg·L-1上升至43.7 mg·L-1(1~19 d)。间歇扰动组与对照组、持续扰动组之间存在显著性差异(P<0.05)。
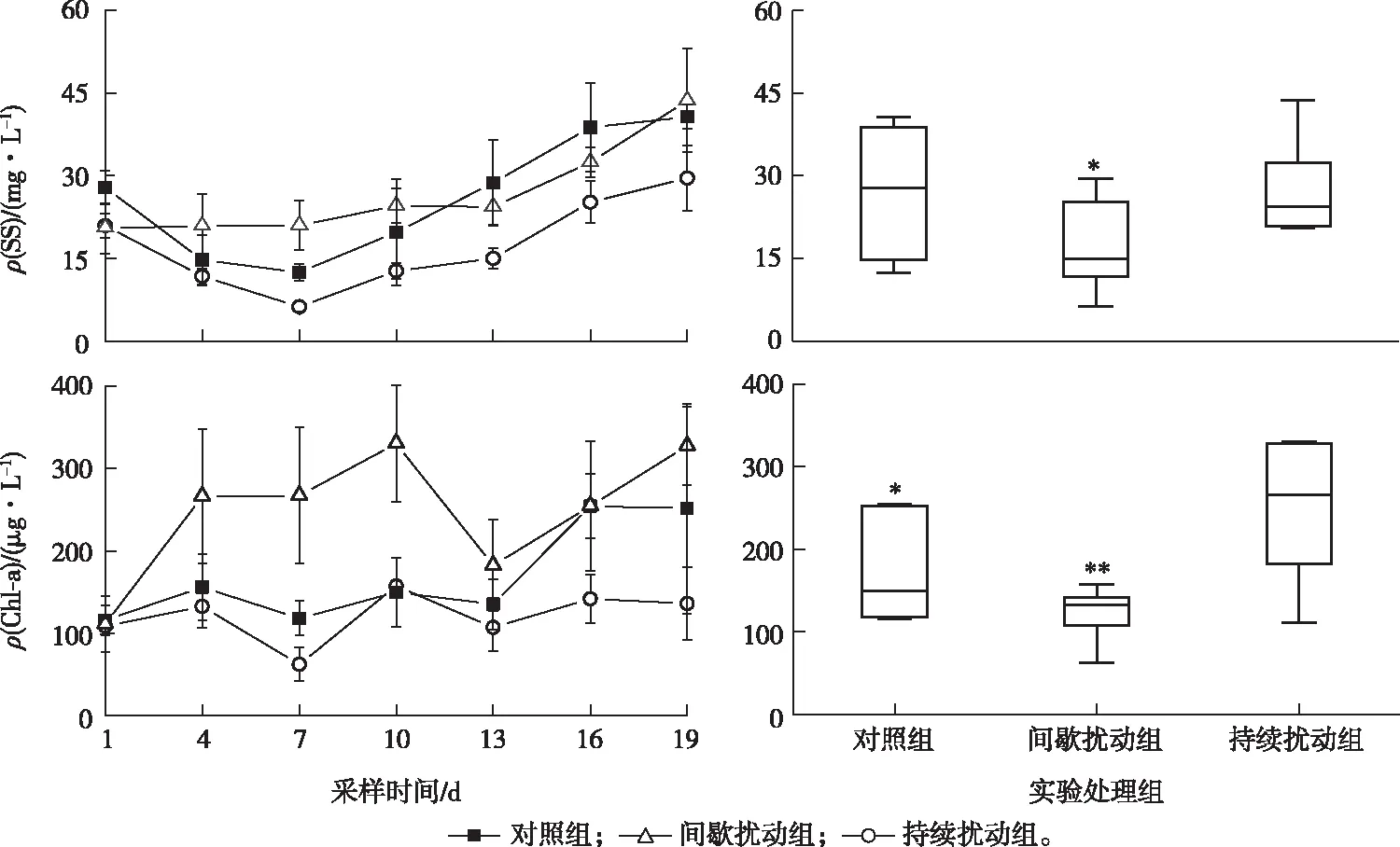
箱体的上下线代表上下四分位数;箱中横线代表中位数;线段上下线代表最大值和最小值,*表示P<0.05,**表示P<0.01。
实验时Chl-a浓度出现了较为明显的波动,3组Chl-a浓度在实验结束时均有所增加,持续扰动组的Chl-a浓度增幅最大,间歇扰动组增加最少。在实验初期(1~10 d)持续扰动组的ρ(Chl-a)持续增加,在第10天达到最大值(330.5 μg·L-1),第14天减小至182.5 μg·L-1,实验结束时又增至327.2 μg·L-1。不扰动组与间歇扰动组ρ(Chl-a)的变化趋势相同,初始值分别为115.9和108.8 μg·L-1,第4天增至156.3和132.6 μg·L-1,随后出现2次波动,第1次波动的最小值分别为118.3和62.4 μg·L-1(第7天),最大值为150.0和157.4 μg·L-1(第10天)。第2次波动在第16天出现最大值,分别为254.5和142.1 μg·L-1,随后分别下降至251.8和136.2 μg·L-1(第19天)。对照组与间歇扰动组、持续扰动组之间存在显著性差异(P<0.05),间歇扰动组与持续扰动组之间存在极显著差异(P<0.01)。
2.5 水体中理化因子的相关性
水体中理化因子的相关性分析结果如表1所示。水体中除NH4+-N外的其余形态的氮与磷之间存在显著相关性,Chl-a浓度与SS浓度呈显著正相关,与TN、TDN、NO3--N浓度之间存在着显著负相关关系。DO浓度与TN、NO3--N、TP浓度呈显著正相关关系。水温与Chl-a浓度呈显著正相关,与氮磷营养盐呈显著负相关关系。
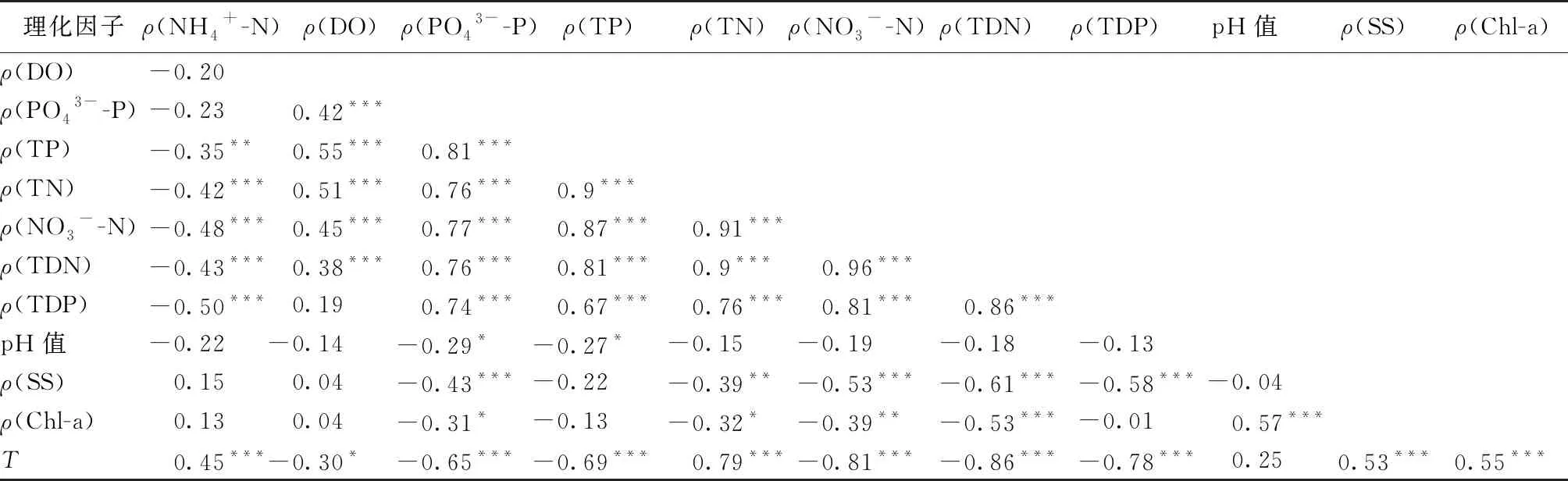
表1 实验期间理化因子的相关性分析
3 讨论
3.1 扰动对水体中氮磷浓度的影响
实验期间3种处理方式下TN和NO3--N浓度均呈下降趋势,但与不扰动相比扰动显著降低了水体中氮浓度。这可能是细菌的反硝化脱氮引起的。传统的生物脱氮包括硝化和反硝化2个独立的过程,白天蓝藻的强烈光合作用使水体产生富氧环境,硝化细菌在碱性环境下将水体中的NH4+-N氧化为NO3--N,夜间藻类的生长堆积、呼吸作用以及衰亡腐烂使水体产生缺氧甚至厌氧环境[16],反硝化细菌在ρ(DO)低于6.4 mg·L-1时转向厌氧呼吸,还原NO3--N为氮气,从而脱除水中氮[17]。水体扰动会在很大程度上增加水中的氧气,使得白天间歇扰动与持续扰动组氧化NO3--N的能力增强[18]。间歇扰动只在白天进行,扰动增加了进入水体的CO2量,为藻类的生长提供碳源,因此其藻类在夜间的呼吸作用、生长堆积和衰亡腐烂消耗的DO会比对照组多,而持续扰动组夜间水体中氧气含量偏高,导致反硝化脱氮能力减弱,最终导致间歇扰动组去除NO3--N的效率最高(图2)。综上所述,间歇扰动的TN、NO3--N浓度显著降低可能与其较低的DO浓度导致反硝化脱氮作用更强有关。然而实验过程中NH4+-N浓度呈波动增长的趋势,这可能与藻类生消过程中藻类残体中有机氮的矿化有关。实验中藻类生消周期约一周左右,NH4+-N浓度的波动周期与此相似,并略有滞后。当藻类分解时,细菌会通过矿化作用使有机氮转化为氨氮,导致NH4+-N浓度的增加[19]。
3种处理方式的TDP和TP浓度均有减少,间歇扰动对促进水体中TDP减少的效果更显著。姚亮宇等[10]发现相对于静水条件,有规律的、一定强度范围内的扰动会促进藻类对营养盐的吸收。这与该研究的结论一致,间歇扰动与持续扰动的TP浓度比对照组降幅大。另外,氮磷浓度与水温呈显著负相关关系,磷的减少可能与实验后期水温升高及藻类增殖有关。温度升高有利于提高藻类代谢速率及生长速率,并通过释放碱性磷酸酶加大对营养盐的吸收[20-22]。需要说明的是,实验过程中部分藻类会附着到桶壁上,这可能是导致水体中TP浓度显著减少的另一个原因。
3.2 扰动对水体中叶绿素含量的影响
持续扰动的Chl-a浓度显著高于其他2个实验组(图4),并且Chl-a浓度与水温呈显著正相关(表1)。这可能是因为温度的上升会加剧水体的垂直分层,不利于较深水体藻类的生长,持续扰动更好地促进了水体垂向混合,利于藻类生长。此外,持续扰动组由于部分水动力扰动的机械能转化为热能,故实验组的水温始终高于其他2个实验组,这可能是持续扰动组Chl-a浓度显著高于其他2个实验组的另一个原因。KARP-BOSS等[23]报道,扰动产生的剪切力会造成藻细胞的机械损伤,破坏其生理活性。该研究显示持续扰动的Chl-a浓度在第10天达到最高,约为开始时的3倍,说明该实验的水动力条件(相当于太湖实际平均风浪强度)对藻类细胞没有明显的机械损伤。研究表明扰动能够影响藻类对光能和营养物质的吸收,改变水体中浮游植物的组成及多样性[24]。该研究也发现持续扰动组在实验后期时微囊藻占比从起始状态的80%减小到40%左右,而绿藻的占比明显升高(数据未呈现)。另外,间歇扰动时的DO浓度低,可能是造成Chl-a浓度低的主要原因之一,这与文献[19]的结论一致。综上所述,适宜强度的持续扰动促进了水体中Chl-a浓度的增高,加大了藻类水华发生的风险。
4 结论
(1)实验期间水体中的TN和NO3--N浓度均持续减少,但间歇扰动对促进水体中氮减少的效果最显著。
(2)实验期间水体中的TP和TDP浓度均持续减少,但间歇扰动对促进水体中TDP减少的效果最显著。
(3)适宜强度的持续扰动促进了水体中Chl-a浓度的增高,有利于藻类生物量的积累。
致谢:感谢中国科学院南京地理与湖泊研究所太湖湖泊生态系统研究站薛静琛、胡洋、邵克强、孙敦平在实验过程及理化数据测试中的帮助。
