白花蛇舌草种子萌发的光敏感性
2020-03-03程凯龙李晓琳王瑞霞杨凤玲刘大伟刘宗瑞王妮娜郭元棣冯佳丽朱梦婷
程凯龙 李晓琳 王瑞霞 杨凤玲 刘大伟 刘宗瑞 王妮娜 郭元棣 冯佳丽 朱梦婷

摘要:探求白花蛇舌草种子的光敏感性,以期为白花蛇舌草野外资源的恢复和栽培生产提供技术支持。研究温度、光照、植物生长调节剂(GA3、6-BA、乙烯和IAA)和含氮化合物(NaNO2、硝普钠和NaNO3)等对白花蛇舌草种子萌发的影响。结果显示,在25~30℃时,光照促进白花蛇舌草种子的萌发,黑暗则抑制其种子的萌发,光周期和光质分别以24h光照/0h黑暗和白光的效果最好;GA3、IAA、6-BA、NaNO2和NaNO3会显著抑制白花蛇舌草种子的萌发,不能在黑暗条件下促进种子萌发,而0.0001~0.01mmol/L的乙烯利可以显著促进白花蛇舌草种子的萌发,0.01mmol/L硝普钠可适当提高白花蛇舌草种子的发芽率。白花蛇舌草种子萌发的最佳技术体系:在25~30℃、24h光照/0h黑暗的白光下,用0.0001~0.01mmol/L的乙烯利处理种子,可以提高发芽率;同时,0.01mmol/L硝普钠可以代替光照诱导萌发。
关键词:白花蛇舌草;光敏感性;萌发;植物生长调节剂;含氮化合物
中图分类号:S567.21+9.01文献标志码:A
文章编号:1002-1302(2020)22-0164-05
通信作者:王瑞霞,博士,副教授,主要从事种子生物学的研究。E-mail:wrx_happy@163.com。
白花蛇舌草(HedyotisdiffusaWilld.)为茜草科(Rubiaceae)耳草属,1年生无毛纤细披散草本。产于广东、香港、广西、海南、安徽、云南等地。据2010年《中华人民共和国药典》记载,白花蛇舌草以全草入药,具有清热解毒、利湿消肿、活血止痛、抗肿瘤等功效[1]。内服用于治疗肿瘤、蛇咬伤、小儿疳积等;外用用于治疗泡疮、刀伤、跌打等。是近年来研究较多的抗癌中草药之一,广泛用于治疗多种恶性肿瘤,市场需求量大增。因此,提高白花蛇舌草的栽培产量和质量,对于满足其市场供应和保护其野生资源具有重要作用。白花蛇舌草的繁殖是播种繁殖,研究白花蛇舌草种子萌发特性对白花蛇舌草资源的恢复和栽培标准化操作规程的制定具有重要意义。
种子萌发是通过播种繁殖的第一步,而植物種子萌发是一个复杂的过程,受多种内外因素的影响,如水分、温度、光、激素及化学试剂等。其中,温度和光照是植物生长和发育重要的环境因子,影响种子的萌发。王鸿彬等通过研究不同恒温和变温条件对江南油杉种子萌发的影响,确定了江南油杉的最佳萌发温度[2]。黄振英等研究发现,梭梭种子萌发的最适温度是10℃,当温度超过20℃时,温度越高,萌发率越低[3]。光对不同植物种子萌发的影响不同。光对有些植物种子的萌发起着重要的调节作用,如龙胆草[4]、红掌[5]、鹅不食[6]、无芒隐子草及条叶车前[7]等的种子在黑暗条件下萌发受到抑制,而矮沙冬青和梭梭的种子萌发在光照和黑暗条件下无显著差异[3,8]。此外,不同光质和光周期对种子萌发的作用效果不同。红光可以促进莴苣种子萌发,蓝光提高了种子发芽指数[9];光暗周期处理可以促进鹅不食种子的萌发[6]。
植物激素也影响着种子萌发。一些植物激素可以部分或者全部代替光照,在黑暗条件下促进种子萌发。赤霉素(GA3)、细胞分裂素和外源乙烯可以促进一些植物的种子萌发,如羊草[10]、油松[11]、拟南芥[12]、番茄[13]和黄独脚金[14]等。
含氮化合物能够与光、植物激素相互作用,促进种子的萌发[15]。一氧化氮(NO)是结构简单、具有水溶性和脂溶性的气体活性分子,参与植物光调节过程,能够促进许多植物种子的萌发,特别是光敏感种子的萌发[16]。外源NO供体为硝普钠(亚硝基铁氢化钠,SNP),0.50mmol/LSNP约能产生2μmol/LNO气体[17]。崔兴国等研究发现,低浓度的SNP处理可以促进黄芪种子萌发,过高浓度的SNP处理则抑制种子的萌发和幼苗生长[18]。土壤中脱氮作用和硝化菌是NO的主要来源,土壤中的硝酸盐和亚硝酸盐浓聚物可以影响种子的萌发[19]。不同种类的硝酸盐可以影响茄子种子萌发[20]。
光周期[21]和光照度[22]会影响白花蛇舌草种子的萌发。郭巧生等研究发现,变温和GA3处理可以提高白花蛇舌草种子的萌发率,并提出白花蛇舌草种子具有光敏特性[23]。但是,尚不明确光质、细胞分裂素和含氮化合物对白花蛇舌草种子萌发的影响。对于白花蛇舌草种子的光敏感性还缺乏系统的研究。
白花蛇舌草种子具有光敏感性,而在黑暗条件下,植物激素和含氮化合物能部分或全部代替光照促进种子萌发吗?不同光质是否影响种子萌发?本项目以白花蛇舌草种子为材料,研究温度、光照、植物激素[GA3、6-苄氨基腺嘌呤(6-BA)、乙烯和吲哚乙酸(IAA)]和含氮化合物(NaNO2、硝普钠和NaNO3)等对种子萌发的影响,以期了解影响其萌发的主要因子,提高发芽率,缩短发芽时间,建立最佳的白花蛇舌草种子萌发技术体系,为野外资源恢复和栽培生产提供科学依据。
1材料与方法
1.1试验材料
白花蛇舌草种子由中国中医科学院中药研究所提供。
1.2白花蛇舌草种子形态学观察
选取饱满的种子,在体式显微镜和扫描电子显微镜下,观测白花蛇舌草种子大小、形状、颜色和种皮表面纹饰等外部形态特征。
1.3温度对白花蛇舌草种子萌发的影响
选取饱满的种子,用75%乙醇消毒30s,再用1%次氯酸钠消毒20min,用蒸馏水冲洗3次,滤纸吸干表面水分,即为供试种子。
将供试种子分别置于15、20、25、30、35℃,光照设定周期性光照[光周期为光照14h/黑暗10h,光照度为12μmol/(m2·s)]和全天黑暗2个处理,每个处理50粒种子,重复3次。以双层滤纸为发芽床,每天同一时间观察并记录种子萌发情况,并保持滤纸湿润(下同)。
1.4光照对种子萌发的影响
光周期对白花蛇舌草种子萌发的影响。将种子分别置于24h/0h、16h/8h、12h/12h、10h/14h、14h/10h、0h/24h的光照/黑暗光周期下,且光照度为12μmol/(m2·s)。
光质对白花蛇舌草种子萌发的影响。将种子分别置于红光、蓝光、白光下,连续照射,光照度为12μmol/(m2·s),温度为25℃。
1.5植物激素对种子萌发的影响
用0、0.0001、0.001、0.01、0.1、1.0mmol/L的GA3、6-BA、乙烯利和IAA处理种子,将种子置于25℃、全天黑暗条件下进行萌发。
1.6含氮化合物对种子萌发的影响
用0、0.005、0.01、0.05、0.1、0.5mmol/L的NaNO2、硝普钠和NaNO3处理种子,将种子置于25℃、全天黑暗条件下进行萌发。
1.7数据分析
处理后,每天同一时间检查种子发芽情况,以胚根突破种皮约1mm时记为萌发。3d后测定发芽势,7d后测发芽率,计算发芽势、发芽率、相对萌发率等各项指标,公式如下:
发芽势=3d后发芽的种子数/处理的种子总数×100%;
发芽率=7d后发芽的种子数/处理的种子总数×100%;
相对发芽率=激素或含氮化合物处理种子的发芽率/对照种子的发芽率。
用SPSS13.0统计分析软件对数据进行单因素方差分析(one-wayANOVA)、多重比较(Student-Newman-Keuls)和一般线性模型分析(generallinearmodelprocedures)(P≤0.05)。
2结果与分析
2.1白花蛇舌草种子形态学观察
白花蛇舌草种皮呈红棕色或棕黄色(图1-A),种子小、微尘样,长250~400μm,宽150~250μm。种子外观上具棱,为不规则多面体。种皮表皮为长方形薄壁细胞,内层细胞网状(图1-B)。
2.2温度对白花蛇舌草种子萌发的影响
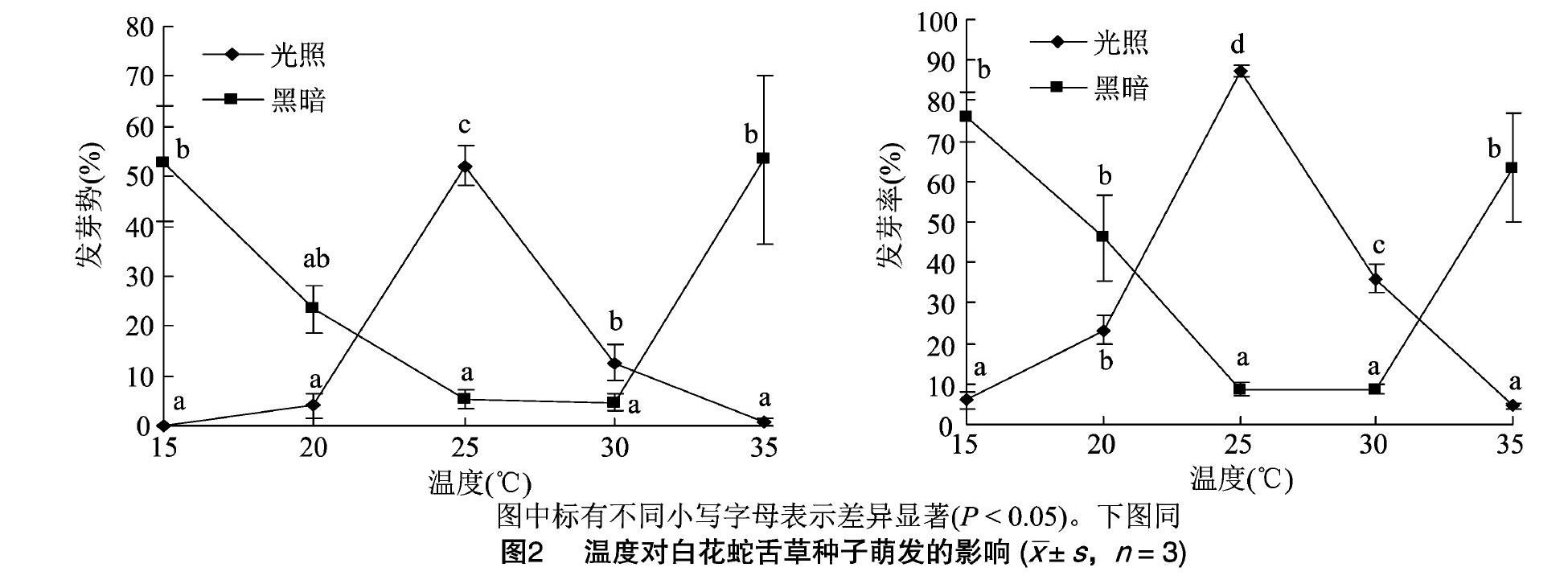
由图2可知,温度会显著影响白花蛇舌草种子的萌發(P<0.05)。在光照条件下,随着温度的升高,发芽势和发芽率先升高再降低,25℃时达到最高值;而在黑暗条件下,则相反,随着温度的升高,发芽势和发芽率均先降低再升高,25~30℃时最低。在25~30℃时,光照会促进白花蛇舌草种子的萌发,而在温度低于20℃或高于30℃时,光照则抑制其种子萌发。
2.3光照对白花蛇舌草种子萌发的影响
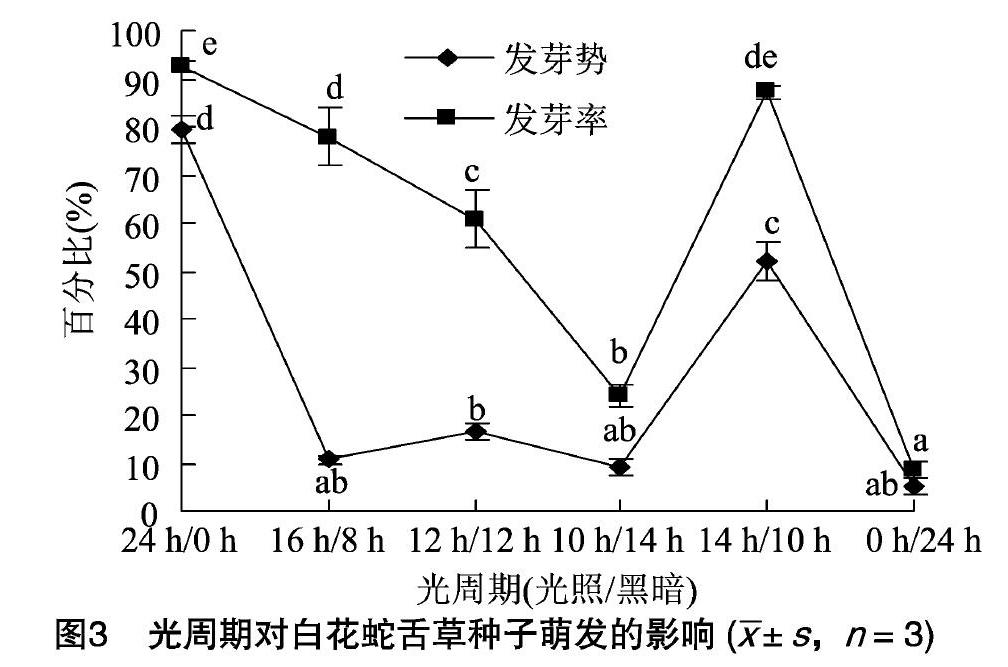
由图3可知,光周期可显著影响白花蛇舌草种子的萌发。发芽势和发芽率以24h/0h光照/黑暗条件下的值最高,14h/10h的次之,而在16h/8h、12h/12h、10h/14h、0h/24h光照/黑暗条件下萌发的种子,发芽势和发芽率均较低。
由图4可知,光质(红光、蓝光、白光)可显著影响白花蛇舌草种子的萌发,发芽势和发芽率均以白光时最高,红光时次之,蓝光时最低。
2.4植物激素对白花蛇舌草种子萌发的影响
由表1可知,0.0001~0.01mmol/L的乙烯利可显著提高白花蛇舌草种子的发芽率,而施用0.0001~1.0mmol/LGA3、IAA和6-BA的白花蛇舌草种子,不仅发芽势、发芽率多数显著低于对照(0.1mol/LIAA处理和1.0mol/L6-BA处理的发芽势除外),而且相对发芽率也低。由此可知,GA3、IAA和6-BA会抑制白花蛇舌草种子的萌发,不能代替光照促进种子萌发。
2.5含氮化合物对白花蛇舌草种子萌发的影响
由表2可知,0.005~0.5mmol/L的NaNO2和NaNO3会显著抑制白花蛇舌草种子的萌发,不能在黑暗条件下促进种子萌发,而0.01mmol/L硝普钠可适当提高白花蛇舌草种子的发芽率,说明0.01mmol/L硝普钠可代替光照诱导白花蛇舌草种子萌发。
3结论与讨论
种子发芽与光的关系被称为光敏感性,这与光敏色素有关[24]。光敏色素和温度间存在互作,不同温度下种子的发芽受到不同光敏色素家族的调控[25]。在25~30℃时,光照促进白花蛇舌草的种子萌发,24h/0h的光照/黑暗光周期效果最好,14h/10h的次之,在此温度范围内,黑暗则抑制其种子的萌发;而当温度低于20℃或[HJ2mm]高于30℃时,情况则恰恰相反,黑暗促进其种子萌发,而光照则抑制种子萌发。本研究结果表明,温度和光周期同时影响白花蛇舌草种子的萌发,这与Lim等的研究结果[21]一致。此外,光质也影响种子萌发[26]。对于白花蛇舌草的种子,在红光和蓝光条件下的发芽势和发芽率均低于白光条件下的。可见,温度、光周期和光质都会影响白花蛇舌草种子的萌发。
GA3、IAA和6-BA抑制白花蛇舌草种子的萌发。这与郭巧生等的研究结果[23]相反。而0.0001~0.01mmol/L的乙烯利可以显著促进白花蛇舌草种子的萌发,这与莫云容等的研究结果[27]相似。
NaNO2和NaNO3显著抑制白花蛇舌草种子的萌发,不能在黑暗条件下促进种子萌发,而0.01mmol/L硝普钠可以适当提高白花蛇舌草种子的发芽率,可代替光照诱导白花蛇舌草种子萌发。这与崔兴国等的研究结果[18]一致。NO在植物体内具有双重作用,低浓度时参与植物生长发育的信号转导过程,而高浓度时则产生过氧亚硝酸,进而破坏生物大分子的结构与功能[28]。
温度、光周期、光质、乙烯利和硝普钠都会影响白花蛇舌草种子的萌发,白花蛇舌草种子萌发的最佳技术体系:在25~30℃、24h光照/0h黑暗光周期的白光下,用0.0001~0.01mmol/L的乙烯利处理种子,可提高发芽率;同时,0.01mmol/L硝普钠可代替光照诱导萌发。该研究结果可为野外资源恢复和栽培生产提供技术支持。
参考文献:
[1]国家药典委员会.中华人民共和国药典[S].一部.北京:中国医药科技出版社,2010.
[2]王鸿彬,刘雄盛,李志辉,等.温度对江南油杉种子萌发特性的影响[J].种子,2018,3(1):46-51.
[3]黄振英,张新时,GuttermanY,等.光照、温度和盐分对梭梭种子萌发的影响[J].植物生理学报,2001,27(3):275-280.
[4]程云清,周革,贾飞飞.光照处理对龙胆草种子萌发的影响[J].吉林师范大学学报(自然科学版),2005,26(3):78-79.
[5]姜蕾,易懋生,兰天维,等.红掌种子萌发特性的研究[J].种子,2006,25(1):19-22.
[6]张舒娜,潘晓曦,马琳,等.光对鹅不食种子萌发及幼苗的影响[J].种子,2016,35(11):91-93.
[7]鱼小军,王彦荣,龙瑞军.光照、盐分和埋深对无芒隐子草和条叶车前种子萌发的影响[J].生态学杂志,2006,25(4):395-398.
[8]杨期和,葛学军,叶万辉,等.矮沙冬青种子特性和萌发影响因素的研究[J].植物生态学报,2004,28(5):651-66.
[9]李雯琳,郁继华,杨其长.不同光质对叶用莴苣种子萌发和幼苗酶活性的影响[J].甘肃农业大学学报,2013,48(3):44-49.
[10]马红媛,梁正伟,黄立华,等.4种外源激素处理对羊草种子萌发和幼苗生长的影响[J].干旱地区农业研究,2008,26(2):69-73.
[11]周建荣.不同激素处理对油松种子萌发特性的影响[J].种子,2016,35(1):85-87.
[12]李凤玲,陈季楚,赵毓橘.赤霉素和光对拟南芥种子萌发和幼苗生长的影响[J].植物生理与分子生物学学报,2000,26(2):101-104.
[13]GrootSPC,KarssenCM.Gibberellinsregulateseedgerminationintomatobyendospermweakening:astudywithgibberellin-deficientmutants[J].Planta,1987,171(4):525-531.
[14]SugimotoaY,AliaAM,YabutaaS,etal.GerminationstrategyofStrigahermonthicainvolvesregulationofethylenebiosynthesis[J].PhysiologyPlant,2003,119(1):137-145.
[15]廖卓毅,周健,钱存梦,等.一氧化氮调控种子休眠与萌发的研究进展[J].安徽农业科学,2015,43(3):7-11.
[16]GibaZ,GrubiicD,KonjevicR.Nitrogenoxidesasenviron-mentalsensorsforseeds[J].SeedScienceResearch,2003,13(3):187-196.
[17]DelledonneM,XiaYJ,DixonRA.Nitricoxidefunctionasasignalinplantdiseaseresistance[J].Nature,1998,394(6693):585-588.
[18]崔兴国,时丽冉.NO供体SNP对黄芪种子萌发和幼苗生长的影响[J].河北农业科学,2011,15(2):107-108.
[19]JacobsenJV,BarreroJM,HughesT,etal.Rolesforbluelight,jasmonateandnitricoxideintheregulationofdormancyandgerminationinwheatgrain(TriticumaestivumL.)[J].Planta,2013,238:121-138.
[20]贡金梅,吴慧,高杰.4种硝酸盐对茄子种子萌发及根系的影响[J].新疆农业科学,2014,51(7):1292-1299.
[21]LimUK,KimSH,LeeHJ.Thegrowth,seedgerminationandanticancereffectofHedyotisdiffusa[J].KoreanJournalofEcology,1994,17(4):523-531.
[22]ZhangM,TanXH,ZhangY,etal.HedayotisdiffusaWilld.seedgerminationandpartsofallelopathy[J].ChineseWildPlantResources,2012,31(1):33-37.
[23]郭巧生,吳传万,刘俊,等.白花蛇舌草种子萌发特性[J].中药材,2001,24(8):548-550.
[24]姜勇,王文杰,李艳红,等.光质、光强对入侵植物紫荆泽兰种子萌发及幼苗状态的影响[J].植物研究,2012,32(4):415-419.
[25]DonohueK,HeschelMS,ButlerCM,etal.Diversificationofphytohromecontributionstogerminationasafunctionofseed-maturationenvironment[J].NewPhytologist,2008,177(2):367-379.
[26]王燕,张亚见,何茂盛,等.光质对植物形态结构和生长的影响[J].安徽农业科学,2018,46(19):22-25.
[27]莫云容,张培欣,朱海山,等.外源乙烯对不同品种辣椒种子萌发的影响[J].云南农业大学学报(自然科学版),2016,30(2):268-273.
[28]ZhangYY,LiuYL.Sourceandfunctionofnitricoxideinplants[J].ActaBotanicaBoreali-OccidentaliaSinica,2004,24(5):921-929.
