青藏高原高寒金露梅灌丛湍流热通量交换与分配特征及其环境影响机制
2020-02-26张法伟李红琴李英年曹广民周华坤
张法伟,韩 赟,李红琴,,李英年,曹广民,周华坤,3
青藏高原高寒金露梅灌丛湍流热通量交换与分配特征及其环境影响机制
张法伟1,2,3,韩 赟1,李红琴1,2,李英年2**,曹广民2,周华坤2,3
(1. 洛阳师范学院生命科学学院,洛阳 471934;2. 中国科学院西北高原生物研究所高原生物适应与进化重点实验室,西宁 810008;3. 中国科学院西北高原生物研究所青海省寒区恢复生态学重点实验室,西宁 810008)
青藏高原重要植被类型之一高寒灌丛的湍流热通量交换是局地微气候特征和植被物候事件的主要调控因素,但其时间格局与分配特征及环境影响机制尚不明确。以青藏高原东北隅的高寒金露梅()灌丛为研究对象,基于涡度相关系统连续观测的湍流热通量,研究该生态系统显热通量和潜热通量交换与分配的特征及潜在环境调控过程。结果表明(1)全年逐时显热通量和逐时潜热通量的平均日变化均表现出单峰型特征,最大值出现在13:30左右。在非生长季(11月−翌年4月)、生长初期(5月)和生长季末期(10月),热量交换以显热通量为主,而在生长季中期(6−9月)则以潜热通量居多;(2)显热通量呈现出双峰型季节特征,最大峰和次高峰分别出现在4月中旬和10月上旬。潜热通量为单峰型季节变化,最大值在7月下旬;(3)湍流热通量的逐时、逐日变异均主要受控于太阳短波辐射;(4)波文比呈现出U型季节变化,而解耦系数、蒸散比例表现为钟型季节变化,热量分配指标在非生长季和生长季分别受控于土壤表层温度和增强植被指数。高寒金露梅灌丛的热量交换主要受控于太阳辐射,热量分配则受下垫面温度和植被覆盖影响。
潜热通量;显热通量;涡度相关系统;波文比;解耦系数
青藏高原被誉为“地球第三级”,是亚洲季风气候形成的重要强迫源之一,是中国乃至全球气候变化的驱动机和放大器[1]。高寒灌丛是青藏高原的重要植被类型之一,面积约0.106×106km2[2],通常位于海拔较高的山区,如山体阴坡或半阴坡或山底的冲积层和洪积层,其碳素固持、水源涵养、气候调节等生态功能对气候变化和人类活动的响应相对于低海拔的高寒草甸更加敏感[3]。近年来,对高寒灌丛的碳素交换及其环境调控机制的研究取得了较大进展[4−8],但对其近地层能量交换与分配的时空特征的研究较为薄弱。地表湍流热通量包括显热通量和潜热通量,其交换强度和分配策略是局地微气候形成的最基本因素和植被活动的重要调控因子[9]。相对于生物地球化学功能(如温室气体吸收),高寒系统的生物物理效应(如蒸散吸热、辐射收支)对全球气候变化的贡献虽然显著但尚待进一步量化[10−11]。而高寒地区环境恶劣,加之观测技术的限制,其热量交换和分配特征及环境控制机制的研究相对较少,成为区域气候模式评估的不确定性来源之一[12−13]。
系统潜热通量和显热通量是下垫面和大气之间的水热垂直梯度及其传输阻抗的物理表达[14−15]。高寒矮嵩草()草甸年均显热通量与潜热通量几乎相等,但高寒沼泽草甸的前者仅为后者的1/2,两种植被类型的显热通量和潜热通量均分别呈现出双峰型和单峰型的季节变化[16−17]。高寒系统的热量分配策略与下垫面水热状况、植被水文多样性、大气蒸发需求密切相关[18−19]。系统热量分配在非生长季中以显热通量为主[16−17, 20]。潜热通量在植被生长季中占比较大,在高寒矮嵩草草甸和高寒沼泽草甸中表现为能量限制,即其分配主要受控于辐射有效能[17, 21],而在高寒草原中则受控于植被生长和土壤水分有效含量[22]。高寒系统的水分收支(降水与潜热蒸散之差)基本处于平衡或盈余状态[21−23]。高寒灌丛的植被以灌木为主、草本嵌套,呈现出较高的景观异质性,而灌木植物相对草本植物,具有更保守型的水分利用策略[24],同时灌丛中苔藓、地衣等隐花植物的盖度较高且其短波反射率和水分过程的时间变异更大[5],更增加了高寒灌丛热量交换与分配特征及其环境控制机制的复杂度,有待进一步深入研究[3]。
以青藏高原东北隅的高寒金露梅()灌丛为研究对象,利用被公认为是地表物质能量交换最准确的观测方法即涡度相关技术[25−26]进行系统湍流热通量交换与分配的研究。由于气候严酷导致仪器运行不稳,经筛选分析发现2004年的全年观测数据有效率较高,约为71%,远高于其余年份的50%左右,因此,选择该年观测数据研究高寒灌丛系统湍流热通量交换和分配的时间特征,探讨环境因子的潜在控制过程,以期深入认知系统的生物物理效应,为准确理解高寒系统能量收支及其分配格局提供数据支撑,也为高寒地区陆面过程模型的发展提供理论依据。
1 材料与方法
1.1 研究区概况
研究点位于青海海北高寒草地生态系统国家野外科学观测研究站(海北站,37°37′N, 101°19′E,3200m)北部约8km的高寒金露梅灌丛(3400m)。研究区为典型的高原大陆性气候,无明显四季之分,暖季多雨而短暂,冷季干燥而漫长。2003−2012年灌丛区域的年均气温和降水分别约为–1.3℃和461.0mm,其中约75%的降水集中在6−9月。研究区的植被群落由灌木和草本镶嵌组成。上层金露梅灌丛高度和盖度约为50cm和60%,下层草本植物高约10㎝,相对盖度约为80%,优势种包括异针茅()、藏异燕麦()、垂穗披碱草()、柔软紫菀()、珠芽蓼()、矮火绒草()等。土壤为沙壤土,归为暗沃寒冻雏形土(Mollic Gryic Cambisols)。研究区为冬季牧场,放牧强度约为3.7羊单位·hm−2,放牧时间约为10月上旬−翌年5月下旬。根据物候观测和相关研究结果[3, 5, 22],11月−翌年4月为非生长季,5−10月为植被生长季,同时将6−9月细分为生长季中期(简称生长季),5月和10月划分为生长季初期和生长季末期(简称过渡期),分别研究非生长季(11月−翌年4月)、生长季(6−9月)和生长过渡期(5月和10月)3个阶段的湍流热通量交换特征。
1.2 数据来源
涡度相关系统安装在地势相对平坦、地形开阔的高寒金露梅灌丛试验场,用于观测该生态系统的潜热通量(LET)、显热通量(H)及微气象环境因子。该系统主要包括开路CO2/H2O快速红外气体分析仪(LI-7500,Li-Cor,美国)和三维超声风速仪(CSAT-3,Campbell,美国),传感器中心距离地面2.2m,仪器采样频率为10Hz,计算并存储30min平均值。系统同步观测相关常规微气象指标,具体包括空气温湿度(HMP45C,Vaisala,芬兰)、风速风向(RM Young,美国)、辐射四分量(向上长、短波辐射和向下长、短波辐射,CM11,Kipp&Zonen,荷兰)和光合光量子通量密度(LI-190SB,LI-Cor,美国)、0.5m高度处的降水量(52203,RM Young,美国)、5cm土壤温度(105T,Campbell,美国)和10cm土壤体积含水量(CS616,Campbell,美国)等环境因子。数据输出均为30min的平均值。详细的观测仪器和方法可参见相关文献[4−5,7]。选取2004年1月1日−12月31日的数据进行分析。由于停电、仪器异常等造成的潜热通量和显热通量数据缺失进行线性插补。观测数据的有效率为71%,能量闭合率为76%,表明湍流热通量的观测数据可信[27]。
采用增强植被指数(EVI)研究植被覆盖对湍流热通量交换的影响。EVI产品是以通量塔为中心周围1km2区域的遥感植被数据。数据来源于美国橡树岭国家实验室分布式活动档案中心(ORNLDAAC,http://daac.ornl.gov/MODIS/modis.html)。EVI的空间和时间分辨率分别为0.25km2和8d。
高寒灌丛湍流热通量传输的空气阻抗(Ra,s·m−1)和冠层阻抗(Rs,s·m−1)的计算式分别为

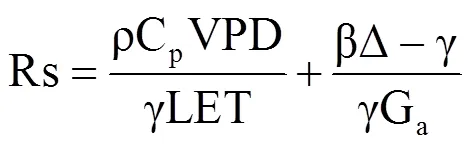
式中,Ws为2.2m处风速(m·s−1),u*为摩擦风速(m·s−1),ρ为平均空气密度(kg·m−3),Cp为空气定压比热(kPa·℃−1),VPD为饱和水汽压差(kPa),γ为空气干湿表常数(kPa·℃−1),LET为潜热通量(W·m−2,L为水的汽化潜热);β为波文比(H/LET,H为显热通量),Δ为水汽饱和曲线斜率(kPa·℃−1)。
解耦系数(Ω)量化了冠层阻抗(Rs)对系统蒸散的控制程度[23, 28],其计算式为

Ω的变化范围在0~1,当Ω接近1时,Rs对系统蒸散影响很小,系统蒸散主要取决于太阳辐射;相反,当Ω趋向于0时,系统蒸散则主要由Rs控制,即受控于下垫面水分的有效供给[22, 29]。Rs和Ω采用8:00−17:00的30min计算值,同时舍去了由于异常天气现象等导致的奇异值[18]。
1.3 统计分析
为了降低天气事件对湍流热通量交换的影响,按照生长季、过渡期和非生长季3个阶段分别将相同时间的显热通量和潜热通量进行平均,以探讨其逐时变化规律。由于环境因子之间较多存在共线性和非独立性,加之数据样本较多(全年17568条),故采用增强回归树(Boosted regression trees,BRT)的方法研究主要环境因子对显热通量和潜热通量逐时变异的相对影响。增强回归树是基于分类回归树算法的一种机器学习方法,通过识别环境因子对全部回归树的节点贡献来量化因子对因变量的相对驱动[30],在R3.61平台[31]上利用Dismo软件包实现。设置学习速率为0.05,树的复杂度为3,每次抽取50%的数据进行分析,并进行10次交叉验证[30]。由于日均样本容量相对较小,采用方差分析、逐步回归、相关分析等方法研究显热通量和潜热通量的季节变化及其与环境因子的关系。
2 结果与分析
2.1 主要环境因子的变化特征
由图1可见,研究区2004年的年均气温和年降水量分别为–1.9℃和493.5mm,为相对冷干的年份,二者基本表现为单峰型季节特征,即生长季雨热同季。土壤体积含水量在植被生长季平均为0.25± 0.04cm3·cm−3,与降水事件密切相关。由于土壤的冻融物理效应,土壤体积含水量在4月和11月表现为跃迁式变化。太阳短波辐射年累计值为6296.8MJ·m−2,最大月均值出现在5月。由于雨热同季,增强植被指数(EVI)最大值为7月底的0.46。但偏相关分析表明,控制降水因子后,EVI与气温呈显著正相关(N=21,P<0.001),反之与降水则无显著关系(P=0.94),暗示气温而非降水的直接作用显著影响高寒灌丛的植被生长。
2.2 湍流热通量交换特征及环境控制机制
2.2.1 日变化特征及环境控制
由图2可见,显热通量(H)和潜热通量(LET)在非生长季、过渡期和生长季均呈现出单峰型日变化特征,夜间基本维持在0值附近;H和LET在3个阶段的平均瞬时最大值依次为51.8和229.2W·m−2、111.3和224.7W·m−2、212.5和157.6W·m−2,峰值均出现在13:30左右。H在非生长季、过渡期和生长季中占日均太阳短波辐射的26.4%、43.7%和19.1%,而LET占太阳短波辐射在3个阶段依次为8.8%、16.8%和29.5%,即非生长季中H约为LET的3倍,而生长季中LET约为H的1.5倍。
以3个阶段的30min显热通量和潜热通量的观测值为因变量,采用增强回归树分析太阳短波辐射、地表长波辐射、气温、空气相对湿度、饱和水汽压差、风速、5cm土壤温度及10cm土壤体积含水量等主要环境因子的相对贡献率。结果表明(图3),3个阶段中,显热通量和潜热通量逐时变异的主要影响因子均为太阳短波辐射,其相对贡献率分别为70%和25%以上;其次,地表长波辐射对生长季潜热通量逐时变异的相对贡献率为23.8%。因此,显热通量的逐时变化主要受太阳短波辐射控制,而潜热通量则主要由太阳短波辐射和地表长波辐射共同影响。

图1 2004年高寒金露梅灌丛逐日平均气温(a)、降水量和10cm土壤体积含水量(b)、太阳短波辐射(c)、增强植被指数(d)变化过程
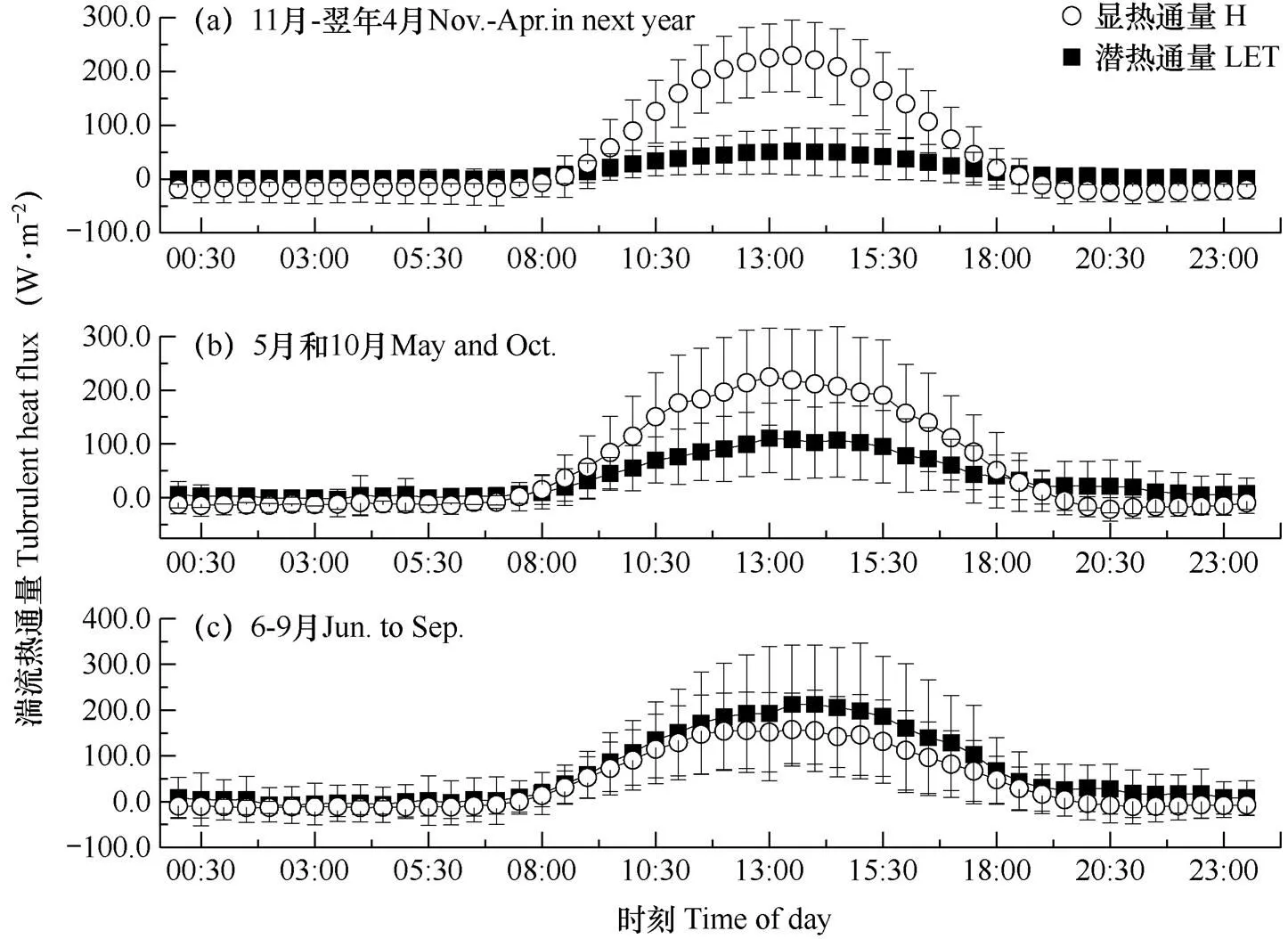
图2 2004年高寒金露梅灌丛非生长季(a)、过渡期(b)和生长季(c)30min湍流热通量的平均日变化过程
注:短线代表标准差。
Note: The error bars were standard deviations.
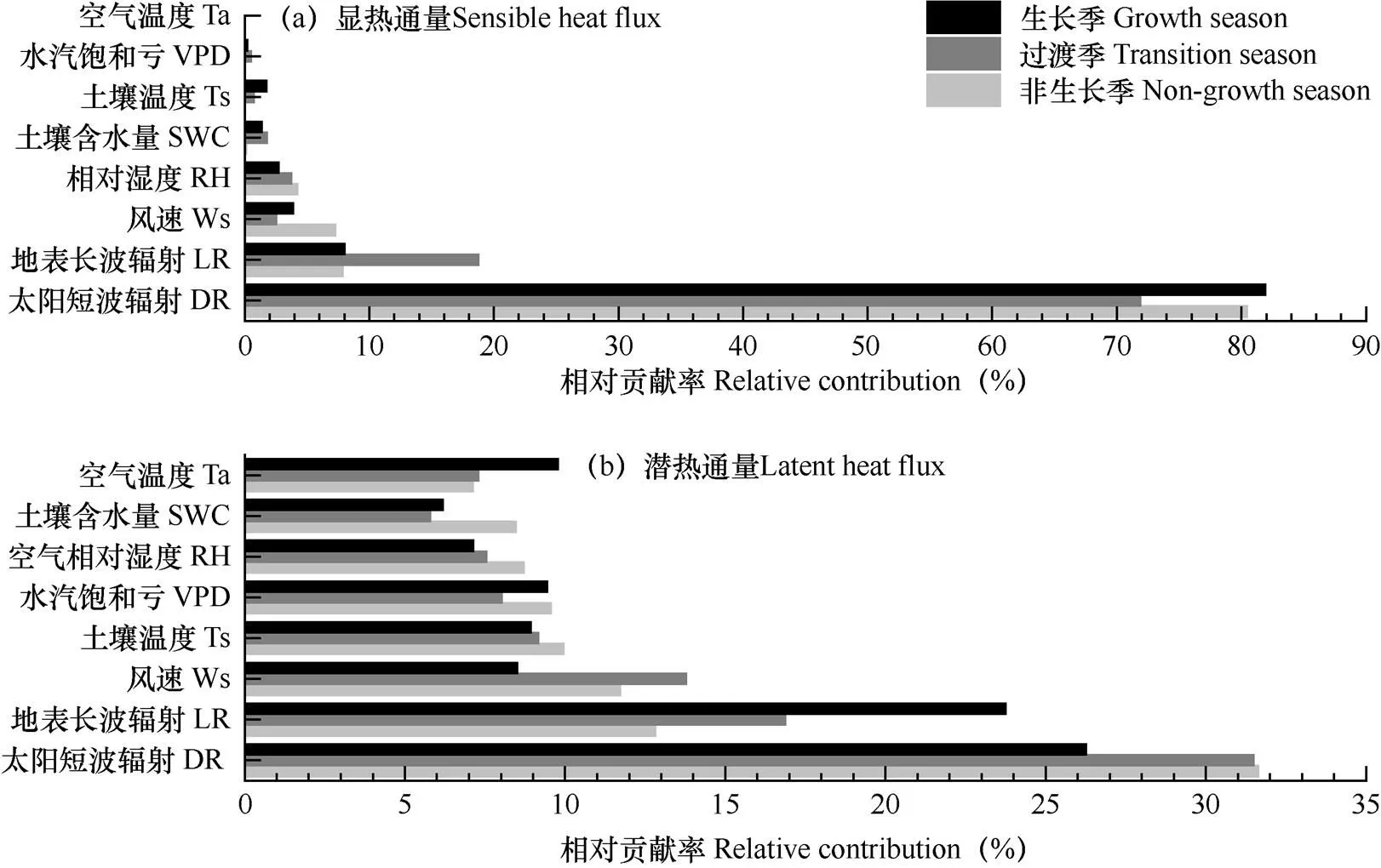
图3 主要环境因子对非生长季、过渡期和生长季3个阶段显热通量(a)和潜热通量(b)逐时变异的相对贡献率
2.2.2 季节变化特征及环境控制
为匹配EVI的时间分辨率,计算逐日湍流热通量和环境因子至8d平均值以进行后续分析。由图4可知,高寒灌丛的显热通量(H)呈现出双峰季节变化特征,日均最大峰和次高峰分别为78.0W·m−2和53.1W·m−2,依次出现在4月中旬和10月下旬。而潜热通量(LET)则表现出单峰型季节变化趋势,最大值(95.0W·m−2)出现在7月下旬。非生长季、过渡期和生长季的日均H和LET分别为46.1和15.2W·m−2、57.2和36.4W·m−2、43.1和67.1W·m−2。H和LET全年累计值分别为1495.1MJ·m−2和1137.7MJ·m−2,年均蒸散发量为503.4mm。

图4 2004年高寒金露梅灌丛湍流热通量的季节变化
逐步回归分析表明,太阳短波辐射(DR)是湍流热通量季节变化的主要环境因子(图5)。图5显示,全年H与DR回归方程的斜率(0.20)小于LET与DR的斜率(0.39),表明LET具有较强的短波辐射敏感度。在非生长季中,H与DR回归方程的斜率(0.29)大于LET与DR的(0.22);在生长季中则相反,分别为0.13和0.38;但过渡期二者相差不大,分别为0.26和0.28。同时,H在非生长季、过渡期和生长季3个生长阶段与DR的回归方程的斜率依次降低,而LET则呈现出逐渐升高的趋势,说明高寒灌丛的植被生长改变了太阳短波辐射对系统热量交换的影响强度和方式。
2.3 湍流热通量分配特征及环境影响
阻抗分析是探讨湍流热通量分配的重要前提。由图6a可知,研究区高寒灌丛日均空气阻抗(Ra)和冠层阻抗(Rs)均呈现出先降低后升高的U型季节动态。年均Ra为50.1s·m−1,其最小值出现在5月初(36.8s·m−1)。而年均Rs为387.9s·m−1,最小值出现在8月底(114.8s·m−1),其中生长季平均为168.3s·m−1。Ra(R2=0.52,P<0.001,N=46)和生长季Rs(R2=0.63,P<0.001,N=21)与风速呈负指数相关。由图6b可知,蒸散比例[LET/(H+LET)、ER]与解耦系数(Ω)则呈现一致的季节趋势,且均与波文比(β)的变化特征相反。在非生长季、过渡期和生长季的日均β分别为5.7、2.3和1.0,3个阶段的日均ER和Ω依次为0.2和0.2、0.4和0.3、0.6和0.6。因此,湍流热通量在生长季主要分配至潜热通量,而另外2个阶段则主要被显热通量消耗。年均β、ER和Ω分别为3.6、0.4和0.4。
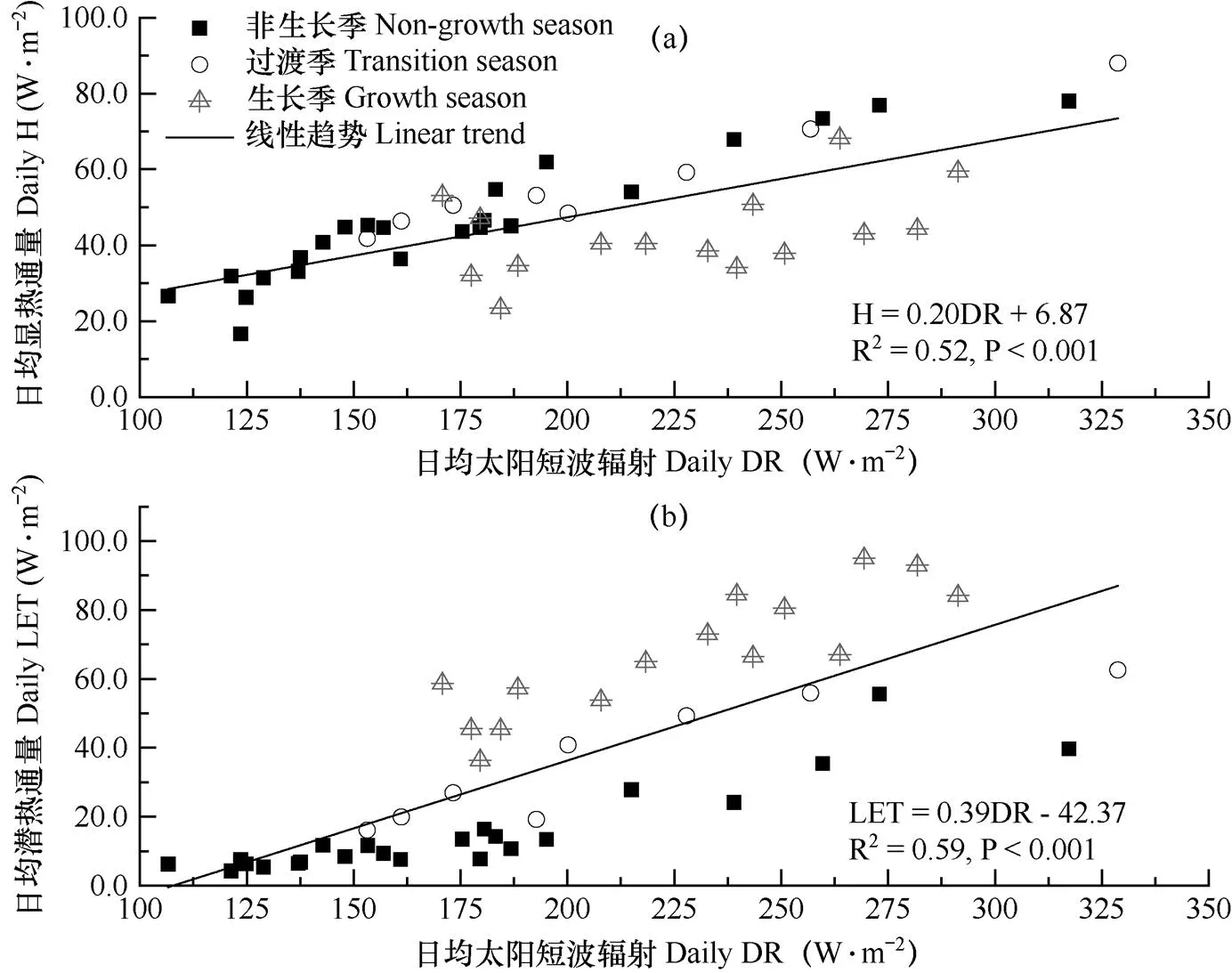
图5 3个阶段日均显热通量(a)、潜热通量(b)与太阳短波辐射(DR)的关系

图6 日均空气阻抗、冠层阻抗(a)和波文比、解耦系数、蒸散比例(b)的变化特征
逐步回归分析表明,土壤温度(Ts)是湍流热通量分配指标(β、Ω和ER)季节变异的主要影响因素。由图7可见,日均β(β=5.9exp(–Ts/15.2)–2.0,R2=0.83)、Ω(Ω=0.3exp(Ts/17.3)+0.05,R2=0.86)和ER(ER=0.5exp(–Ts/22.6)+0.2,R2=0.83)均与日均Ts呈现出显著指数关系(图7a)。生长季β、Ω和ER与增强植被指数(EVI)线性相关(图7b)。但控制Ts后,EVI与β的偏相关分析表明EVI(N=21,P=0.043),而并非Ts(N=21,P=0.77),是生长季日均β变异的显著控制因子。同理,生长季Ω也主要受EVI(P=0.05)的调控。但ER与Ts和EVI的偏相关分析表明,Ts(P=0.98)和EVI(P=0.10)对ER的单独效应均不显著。
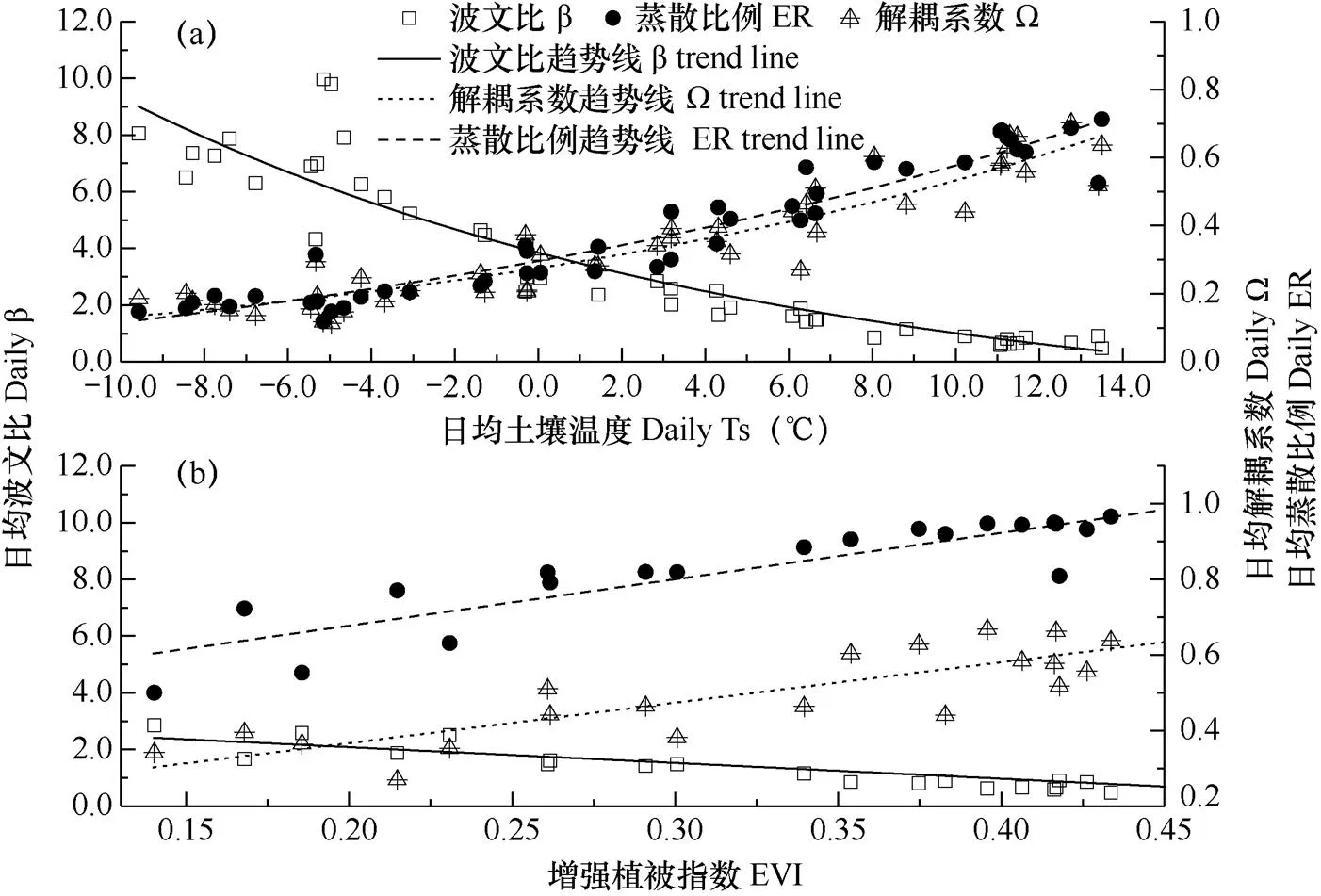
图7 日均波文比、蒸散比例、解耦系数与全年日均土壤温度(Ts,a)及生长季增强植被指数(EVI,b)的关系
3 结论与讨论
3.1 讨论
3.1.1 太阳辐射对高寒系统湍流热通量交换的影响
太阳短波辐射是系统湍流热通量在短时间尺度的主要控制因子,表明高寒灌丛的湍流热通量交换受辐射能量限制,这与邻近较低海拔(3200m)高寒草甸的研究结果一致[3, 17, 21]。这可能与系统辐射收支及下垫面状况两个因素有关。(1)尽管高寒灌丛的太阳短波辐射较高[7],其瞬时极值达1213W·m−2,但由于长波净辐射损失较高[16],其年均长波净辐射为–79.4W·m−2,长波损失率(长波净辐射占太阳短波辐射的百分率)为38.7%,系统有效辐射能量相对不高,致使区域湍流热通量与太阳短波辐射呈现显著正线性相关而并非渐进饱和特征[21];(2)下垫面土壤水分供应相对充足,生长季约为0.25cm3·cm−3,远超过高寒灌丛的土壤凋萎含水量0.095cm3·cm−3[21],再加上相对充沛的生长季降水补给(444.3mm),虽然潜热通量消耗了土壤水分,但其消耗强度还远未达到土壤水分反过来限制潜热通量的程度[10−11],表明高寒灌丛潜热通量不会受到下垫面水分供给的限制。另外,生长季潜热通量与土壤体积含水量的负相关关系(R2=0.24,P=0.02,N=21)也暗示土壤水分供给不会成为高寒灌丛潜热通量的限制因素。
近期有关研究发现,青藏高原腹地的高寒紫花针茅()草原的潜热通量主要受土壤水分限制[22]。这可能是由于高寒草原一般分布在高原中部和西部,其气候相对干燥和寒冷,导致优势种多以耐寒、耐旱植被为主。而高寒灌丛、草甸则主要分布在青藏高原东北部和东南部,区域气候相对湿润和温暖,优势种多以耐寒中生型植被为主[2]。因此,在评估青藏高原气候演变[1]和发展区域气候模式[10]时,应充分考虑不同植被生活型导致的地气潜热通量交换的差异所引起的大气边界层变化[11, 24]。
3.1.2 植被状况对湍流热通量的影响
植被状况对湍流热通量的影响主要通过改变热量通量(强度和分配)的直接方式和影响系统辐射收支(短波反射率和长波收支)等间接途径[9, 11]。植被生长降低了地表反射率(非生长季和生长季分别为0.19和0.13)和长波辐射损失率(非生长季和生长季分别为0.51和0.32),表明植被生长有利于高寒灌丛系统辐射能的吸收[9, 11]。生长季中潜热通量与增强植被指数和太阳短波辐射的一般线性模型(R2=0.96,P<0.001)表明,太阳短波辐射(P=0.004)而并非增强植被指数(P=0.07)是潜热通量的主要影响因素。另外生长季的解耦系数高达0.6,也表明该系统的潜热通量主要受辐射能控制[16, 21]。即植被生长对湍流热通量交换强度的影响较为有限。同时,本研究也表明热量分配(波文比、解耦系数和蒸散比例)主要受增强植被指数调控,表明植被生长是影响系统热量分配的重要因素[13, 16]。所以,本研究推测植被生长主要调控高寒灌丛湍流热通量的分配特征而非交换强度,这与邻近区域高寒草甸[21]和祁连云杉林[20]的研究结果较为一致。
3.2 结论
高寒金露梅灌丛湍流热通量交换平均日变化表现为单峰型特征,在非生长季以显热通量为主,约为潜热通量的3倍,而在生长季潜热通量略大,约为显热通量的1.5倍。逐日显热通量和潜热通量分别呈现出双峰型和单峰型季节变化。太阳短波辐射是湍流热通量逐时、逐日交换的主要调控因子。波文比、解耦系数和潜热比例等热量分配指标在生长季受控于增强植被指数,在非生长季受土壤温度影响。研究结果表明高寒金露梅灌丛湍流热通量交换强度主要受太阳辐射能量限制,而分配方式则受表层土壤温度和植被生长状况影响。
[1] 冯松,汤懋苍,王冬梅.青藏高原是我国气候变化启动区的新证据[J].科学通报,1998,43(6):633-636.
Feng S,Tang M C,Wang D M.New evidence of Tibetan Plateau is the start area of climate change in China[J].Chinese Science Bulletin,1998,43(6):633-636.(in Chinese)
[2] Zheng D,Zhang Q S,Wu S H.Mountain geoecology and sustainable development of the Tibetan Plateau[M].Dordercht, the Netherlands: Kluwer Academic,2000.
[3] Li H Q,Zhu J B,Zhang F W,et al.Growth stage-dependant variability in water vapor and CO2exchanges over a humid alpine shrubland on the northeastern Qinghai-Tibetan Plateau [J].Agricultural and Forest Meteorology,2019,268:55-62.
[4] Li H Q,Zhang F W,Li Y N,et al.Seasonal and interannual variations of ecosystem photosynthetic features in an alpine dwarf shrubland on the Qinghai-Tibetan Plateau,China[J]. Photosynthetica,2014,52(3):321-331.
[5] Li H Q,Zhang F W,Li Y N,et al.Seasonal and inter-annual variations in CO2fluxes over 10 years in an alpine shrubland on the Qinghai-Tibetan Plateau,China[J].Agricultural and Forest Meteorology,2016,228-229:95-103.
[6] Li H Q,Zhang F W,Wang W Y,et al.The strongest EI Niño event stimulated ecosystem respiration,not evapotranspiration, over a humid alpine meadow on the Qinghai-Tibetan Plateau [J].Ecological Indicators,2018, 91:562-569.
[7] Li Y N,Sun X M,Zhao X Q,et al.Seasonal variations and mechanism for environmental control of NEE of CO2concerning thein alpine shrub meadow of Qinghai-Tibet Plateau[J].Science in China Series D-Earth Sciences,2006,49(Supply 2):163-173.
[8] 葛怡情,闫玉龙,梁艳,等.模拟降水氮沉降对藏北高寒草甸土壤呼吸的影响[J].中国农业气象,2019,40(4):214-221.
Ge Y Q,Yan Y L,Liang Y,et al.The effects of nitrogen deposition on soil respiration in an alpine meadow in northern Tibet[J].Chinese Journal of Agrometeorology,2019,40(4):214- 221.(in Chinese)
[9] Chapin F S,Matson P A,Mooney H A.Principles of terrestrial ecosystem ecology[M].Seconded,New York,USA:Springer- Verlag,2011:142-145.
[10] Zeng Z Z,Peng L Q,Piao S L.Response of terrestrial evapotranspiration to Earth's greening[J].Current Opinion in Environmental Sustainability,2018,33:9-25.
[11] Zeng Z Z,Piao S L,Li L Z X,et al.Climate mitigation from vegetation biophysical feedbacks during the past three decades[J].Nature Climate Change,2017,7:432-436.
[12] Shen M G,Piao S L,Jeong S,et al.Evaporative cooling over the Tibetan Plateau induced by vegetation growth[J]. Proceedings of the National Academy of Sciences of the United States of America,2015,112(30):9299-9304.
[13] 秦孟晟,郝璐,郑箐舟,等.秦淮河流域土地利用/覆被变化对蒸散量变化的贡献[J].中国农业气象,2019,40(5):269-283.
Qin M S,Hao L,Zheng Q Z,et al.Contributions of land use/cover change to the change of evapotranspiration in Qinhuai River Basin[J].Chinese Journal of Agrometeorology,2019,40(5):269-283.(in Chinese)
[14] Hu Z M,Yu G R,Fu Y L,et al.Effects of vegetation control on ecosystem water use efficiency within and among four grassland ecosystems in China[J].Global Change Biology, 2008,14(7):1609-1619.
[15] 张法伟,王军邦,林丽,等.高寒草甸非生长季土壤表层水汽传输阻抗的变化特征和水热驱动[J].中国农业气象,2017, 38(2):96-103.
Zhang F W,Wang J B,Lin L,et al.Temporal variations of soil surface resistance to vapor transfer and its quantitative relationship between soil temperature and soil moisture during non-growing season on an alpine meadow[J].Chinese Journal of Agrometeorology,2017,38(2):96-103.(in Chinese)
[16] Gu S,Tang Y H,Cui X Y,et al.Energy exchange between the atmosphere and a meadow ecosystem on the Qinghai- Tibetan Plateau[J].Agricultural and Forest Meteorology, 2005,129(3):175-185.
[17] Zhang F W,Li H Q,Li Y N,et al.Surface energy partitioning in alpine swamp meadow in the Qinghai Tibetan Plateau[J]. Pratacultural Science,2008,25(4):14-23.
[18] McFadden J P,Eugster W,Chapin F S.A regional study of the controls on water vapor and CO2exchange in arctic tundra[J].Ecology,2003,84(10):2762-2776.
[19] Wilson K B,Baldocchi D D,Aubinet M,et al.Energy partitioning between latent and sensible heat flux during the warm season at FLUXNET sites[J].Water Resources Research, 2002,38(12):1294.
[20] Zhu G F,Lu L,Su Y H,et al.Energy flux partitioning and evapotranspiration in a sub-alpine spruce forest ecosystem [J].Hydrological Processes,2014,28(19):5093-5104.
[21] Zhang F W,Li H Q,Wang W Y,et al.Net radiation rather than moisture supply governs the seasonal variations of evapotranspiration over an alpine meadow on the northeastern Qinghai-Tibetan Plateau[J].Ecohydrology, 2018, 11(2):e1925.
[22] Ma N,Zhang Y S,Guo Y H,et al.Environmental and biophysical controls on the evapotranspiration over the highest alpine steppe[J].Journal of Hydrology,2015,529:980- 992.
[23] Gu S,Tang Y H,Cui X Y,et al.Characterizing evapotranspiration over a meadow ecosystem on the Qinghai-Tibetan Plateau[J].Journal of Geophysical Research-Atmospheres, 2008,113(D8).
[24] Teuling A J,Seneviratne S I,Stockli R,et al.Contrasting response of European forest and grassland energy exchange to heatwaves[J].Nature Geosciences,2010,3(10):722-727.
[25] Baldocchi D D.Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems:past, present and future[J].Global Change Biology,2003,9(4): 479-492.
[26] 原文文,张劲松,孟平,等.开路与闭路涡度相关系统对森林生态系统甲烷通量观测的比较[J].中国农业气象,2019, 40(11):669-677.
Yuan W W,Zhang J S,Meng P,et al.Comparison of CH4flux measurement by open- and close-path eddy covariance system[J].Chinese Journal of Agrometeorology,2019,40(11): 669-677.(in Chinese)
[27] Wilson K,Goldstein A,Falge E,et al.Energy balance closure at FLUXNET sites[J].Agricultural and Forest Meteorology, 2002,113(1-4):223-243.
[28] Jarvis P G,Mcnaughton K G.Stomatal control of transpiration:scaling up from leaf to region[J].Advances in Ecological Research,1986,15:1-49.
[29] Baldocchi D D,Vogel C A,Hall B.Seasonal variation of energy and water vapor exchange rates above and below a boreal jack pine forest canopy[J].Journal of Geophysical Research-Atmospheres,1997,102(D24):28939-28951.
[30] Elith J,Leathwick J R,Hastie T.A working guide to boosted regression trees[J].Journal of Animal Ecology,2008,77: 802-813.
[31] R Development Core Team.R:a language and environment for statistical computing[DB/CD].Vienna:R Foundation for Statistical Computing,2006.
Turbulent Heat Exchange and Partitioning and Its Environmental Controls between the Atmosphere and an AlpineShrublands over the Qinghai- Tibetan Plateau
ZHANG Fa-wei1, 2, 3, HAN Yun1, LI Hong-qin1, 2, LI Ying-nian2, CAO Guang-min2, ZHOU Hua-kun2, 3
(1. College of Life Sciences, Luoyang Normal University, Luoyang 471934, China; 2. Key Laboratory of Adaptation and Evolution of Plateau Biota, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008; 3. Qinghai Provincial Key Laboratory of Restoration Ecology in Cold Region, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008 )
The turbulent heat flux plays important role in micro-climate environment and vegetation phenology but its temporal patterns and partitioning characteristics and associated environmental controls remain unclear in alpine shrublands, which is one of the most important vegetation types on the Qinghai-Tibetan Plateau. The continuous turbulent heat flux and routine environmental variables measured by the eddy covariance techniques were analyzed to quantify the exchange and partitioning of sensible heat flux and latent heat flux over an alpineshrublands on the northeastern Qinghai-Tibetan Plateau. The results showed that (1) the averaged diurnal variations of turbulent heat exchange both exhibited unimodal patterns, peaking at about 13:30 over the whole year-round period. The diurnal heat flux was dominated by sensible heat flux during non-growth season (November to next April) and the beginning and end of growth season (May and October), while by latent heat flux in mid-growth season from June to September; (2) The daily sensible heat flux exhibited a bimodal seasonal pattern, with the largest peak and the second peak appearing in mid-April and beginning October, respectively. The daily latent heat flux presented a unimodal seasonal pattern with a maximum in end July; (3) The diurnal and daily variations of turbulent heat flux were both mainly controlled by solar shortwave radiation; (4) Bowen ratio showed a U-shape seasonal change, while decoupling coefficient, evaporation ratio exhibited a bell-shape seasonal variation. These partitioning indices were controlled by soil temperature during non-growth season and enhanced vegetation index in growth season, respectively. These results revealed that the turbulent heat exchange was determined by solar radiation while the partitioning between sensible heat flux and latent heat flux was regulated by underlying surface temperature and plant coverage in the alpine shrublands.
Latent heat flux; Sensible heat flux; Eddy covariance technique; Bowen ratio; Decoupling coefficient
10.3969/j.issn.1000-6362.2020.02.002
张法伟,韩赟,李红琴,等.青藏高原高寒金露梅灌丛湍流热通量交换与分配特征及其环境影响机制[J].中国农业气象,2020,41(2):76-85
2010−09−23
李英年,E-mail:ynli@nwipb.cas.cn
国家自然科学基金(41877547;41730752);国家重点研发计划(2017YFA0604801);青海省科技基础条件平台建设专项项目(2018-ZJ-T09);青海省创新平台建设专项(2017-ZJ-Y20)
张法伟,E-mail:mywing963@126.com
