不同光质夜间补光对番茄幼苗耐冷性的影响
2020-02-22刘福长杨玉凯林碧英
刘福长 杨玉凯 林碧英

摘要:【目的】探讨不同光质夜间补光对番茄幼苗耐冷性的影响,为提高番茄幼苗耐冷性提供理论和实践依据。【方法】以倍盈番茄为试验材料,采用光质分别为白光(W)、红光(R)、蓝光(B)和红蓝组合光配比(2R/8B、SR/5B、8R/2B)等6个组合进行夜间补光处理,20d后各处理番茄幼苗置于亚低温(100C/5℃)下胁迫,以未补光所培育番茄幼苗为对照。【结果】与对照相比,各补光处理所培育的番茄幼苗在亚低温处理期间叶片中的电解质渗透率和丙二醛(MDA)含量上升幅度均显著低于对照,红蓝复合光(2R/8B、SR/5B、8R/2B)在亚低溫胁迫期间叶片一直保持着较高的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)活性,在亚低温胁迫6d时以红蓝复合8R/2B活性上升幅度最高,分别达到399.49%、129.01%和135.74%,显著高于对照。【结论】夜间补光处理均可提高番茄幼苗的耐冷性,其中以红蓝复合8R/2B补光处理的幼苗的抗低温胁迫能力最强。
关键词:番茄;苗期;补光;光质;低温胁迫;耐冷性;逆境生理指标
中图分类号:S641.2
文献标志码:A
文章编号:1008-0384(2020)11-1222-06
0引言
【研究意义】设施温室在蔬菜反季节栽培和早春育苗方面起主导作用,但蔬菜生长发育时期易受低温寡照的不利影响。近年来南方冬季经常出现异常低温情况,如2016年1月福建省遇到大寒流,导致设施温室内的番茄和辣椒等喜温蔬菜受到严重的冻害,经济损失严重。提高植物自身的耐冷性是减少冻害的重要因素。【前人研究进展】光是植物生长发育最重要的环境因子之一,光照强度和光质对植物的生长、光形态建成和生理代谢均有显著影响[1],可提高植物对虫害的抗性[2]。光质对于植物对环境胁迫(低温、高温、干旱、盐分)的响应也有重要影响,通过调整光质可显著提高植物对环境胁迫的抗性[3]。研究发现,光敏色素是启动冷驯化的光受体,可通过控制相关的冷诱导基因的表达进而影响植物的耐冷性[4]。超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)均属于酶促防御系统,与白光对照相比,红光和蓝光培育的番茄幼苗在低温胁迫下依然保持较高的酶活性[5],与白光对照相比,黄光处理下玉湖莴苣在低温胁迫下SOD、POD、CAT活性提升最高,有利于提升莴苣的耐寒性[6]。【本研究切入点】前人在番茄上的光环境调控研究,主要体现在光环境对于番茄产量和品质的调控上,但对于提升番茄幼苗耐冷性的研究较少。【拟解决的关键问题】本研究以番茄为研究材料,设计6个补光处理,以不补光为对照,探讨不同光质的夜间补光处理对番茄幼苗耐冷性的影响,以期为提高番茄幼苗耐冷性提供理论和实践依据。
1材料与方法
1.1试验材料
供试番茄(Lycopersicon esculentum)品种为倍盈,原产地为以色列,种子由福州市蔬菜科学研究所提供。
1.2试验光质设计
试验LED光源均采购于惠州可道科技股份有限公司。红光R(峰值波长660nm)、蓝光B(峰值波长450nm),白光(W)的波长范围410-760 nm,3种颜色光质的光照强度可调节范围为0~255μmol·m-2·s-1。光质处理在智能光照培养箱中进行,培养箱内部构造为4层×2列,每个独立隔间规格为60cm×60cm×60cm(长×宽×高),内部采用银白色隔板,可有效促进内部光照的均匀度。每个隔间配有单独的风扇和温度传感器,当温度高于所设定温度可自行降温调节。LED灯条可通过磁铁吸附在可升降的灯板上,通过调节灯板的高度使各处理光强均保持在100μmol·m·s-1,光强用HiPoint HR 350 LED光谱计测定。LED光源设置6个处理,分别是白光W、红光R、蓝光B和红蓝组合光配比(2R18B、5R/5B、8R/2B),以不补光为对照(CK)(表1)。
1.3试验过程
试验于2017年9月至2018年5月在福建农林大学现代农业设施温室和人工气候室内进行。前期对种子进行常规温汤浸种,然后将种子置于28℃恒温箱中催芽,待种子“露白”后,即可进行播种,播于72孔穴苗盘中育苗,育苗基质为m草炭:m珍珠岩:m蛭石=3:1:1。待幼苗子叶展开新叶显露后,将番茄幼苗移到10cm×8cm的营养袋,缓苗2d后开始试验处理。试验期间白天番茄幼苗在智能温室中进行培养,夜间搬人智能光照培养箱进行4h补光处理,补光时段为(18:00~22:00),白天温度25℃左右,夜间温度18℃,湿度为60%~70%,每隔2d浇灌100mL的山崎番茄专用营养液。
将不同光质补光处理20d后的番茄幼苗放入低温培养箱中进行亚低温胁迫处理,温度设置为10/5℃(昼/夜),相对湿度为50%~60%,光照时长均为12h(昼/夜),光照强度为5000 Lx,于亚低温胁迫处理第0、3、6d随机选取3株叶片剪碎混合后进行番茄幼苗的相关生理生化指标的测定。
1.4测定指标与方法
丙二醛( MDA)含量的测定参考陈建勋等[7]的方法;采用电导仪法测定亚低温胁迫下叶片的电解质渗透率[8];脯氨酸含量采用磺基水杨酸法测定[8];过氧化氢酶(CAT)活性的测定采用紫外吸收法[8],过氧化物酶活性(POD)的测定采用愈创木酚法[8];超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑(NBT)法[8]。
1.5数据处理
试验数据使用SPASS20.0统计软件进行显著性分析和方差分析。
2结果与分析
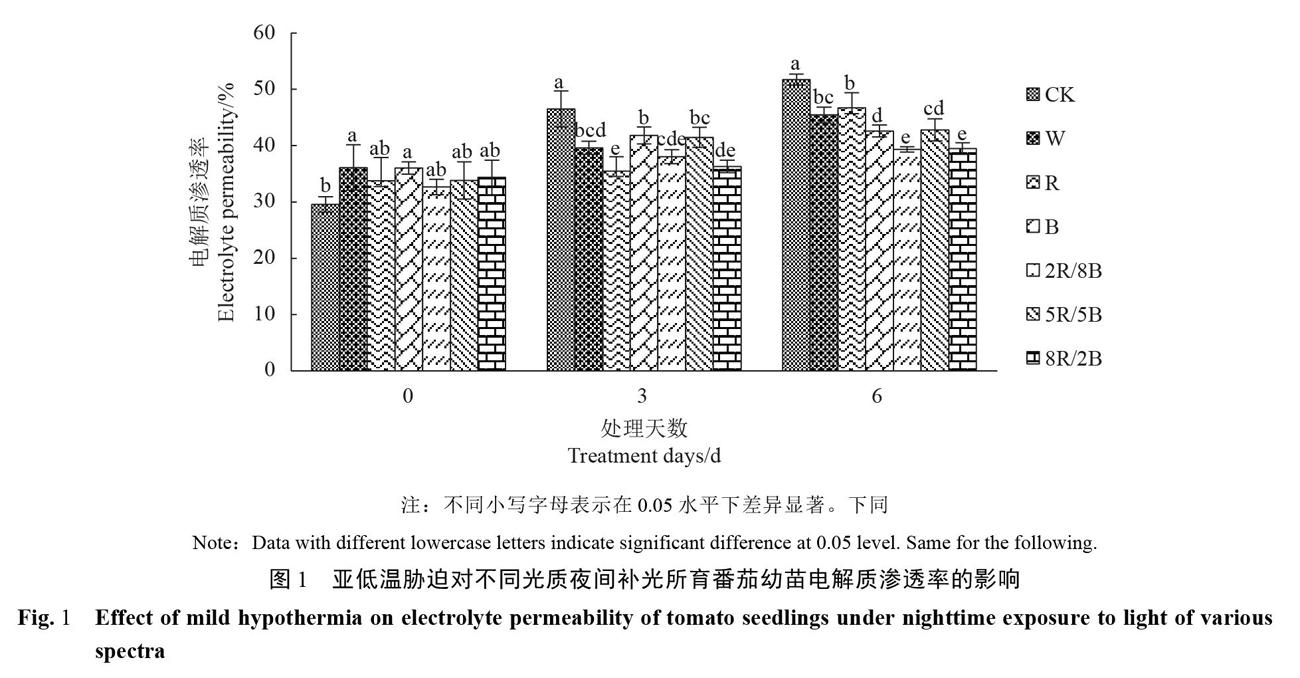
2.1亚低温胁迫对番茄幼苗电解质渗透率的影响
植物受到亚低温胁迫时,会导致细胞膜受损,质膜透性变大,进而导致电解质渗漏。图1显示,随着亚低温胁迫时间的延长,各处理的番茄幼苗电解质渗透率均呈现上升趋势。在亚低温胁迫处理第3、6d时,各补光处理的电解质渗透率均低于对照,且均达到显著性差异。在亚低温胁迫处理第6d时,其中对照电解质渗透率与亚低温胁迫开始时相比升幅达74.82%,其次为R补光处理,电解质渗透率上升幅度为38.36%,上升幅度最小的为8R/2B补光处理,仅为14.88%。
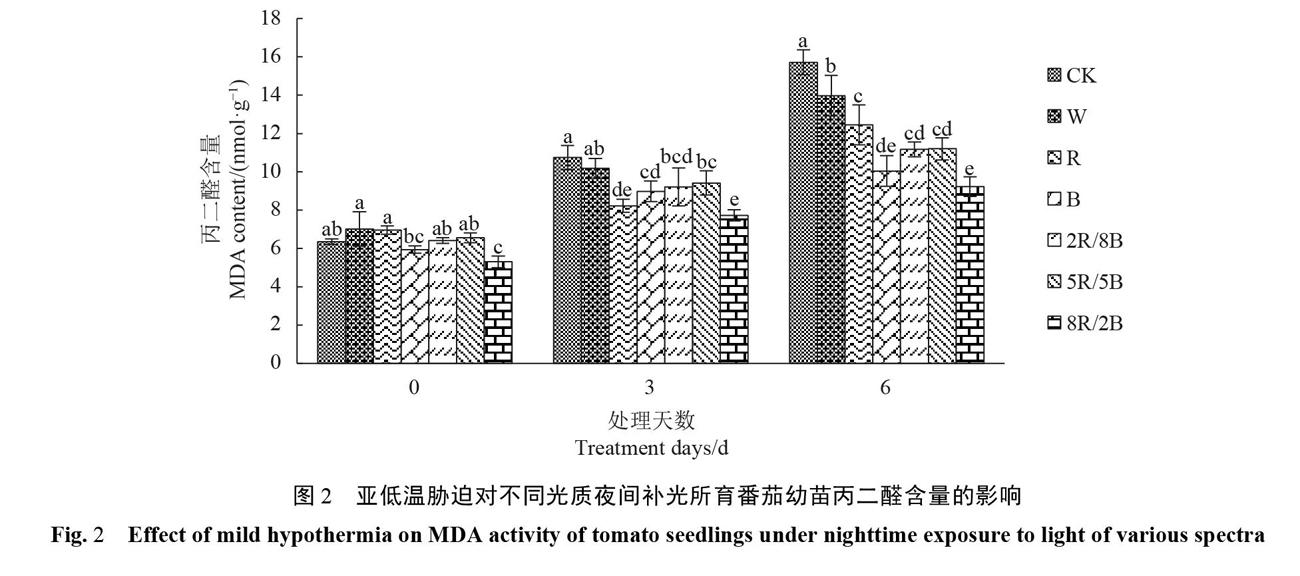
2.2亚低温胁迫对番茄幼苗丙二醛含量的影响
图2显示,随着亚低温胁迫时间的延长,各处理番茄幼苗叶片的MDA含量均呈上升趋势,且变化趋势与电解质渗透率变化相似。在亚低温胁迫处理第3d时,各补光处理的电解质渗透率均低于对照,除W补光处理外,其他补光处理均达到显著性差异。在亚低温胁迫处理第6d,与亚低温胁迫开始时相比,各补光处理的幼苗叶片中MDA含量上升幅度均低于对照,上升幅度大小具体表现为:CK(147.64%)>W(98.93%)>R(79.37%)>5R/5B(78.60%)>8R/2B(73.58%)>2R/8B(72.32%)>B(69.27%)。
2.3亞低温胁迫对番茄幼苗SOD活性的影响
图3显示,在亚低温胁迫处理下,各补光处理与对照所培育幼苗叶片中SOD活性变化一致,均是随着亚低温胁迫时间的延长SOD活性均大幅增加。在亚低温胁迫处理第3d时,各补光处理的SOD活性均高于对照,且均达到显著性差异,其中W补光处理的SOD活性最高。在亚低温胁迫处理第6d时,R、B补光处理和红蓝复合光(2R/8B、5R/5B、8R/2B)补光处理的番茄幼苗SOD活性与Od相比上升幅度均高于对照和W补光处理,其中8R/2B补光处理的幼苗叶片中SOD的活性相较于亚低温胁迫开始时上升幅度最大(399.4g%),其次为B补光处理(388.19%),对照下培育的幼苗SOD活性上升幅度最小(仅为271.04%)。
2.4亚低温胁迫对番茄幼苗POD活性的影响
图4显示,在亚低温胁迫处理下,各处理的番茄幼苗叶片中POD活性变化趋势与SOD活性变化一致,均是随着亚低温胁迫时间的延长而增加。在亚低温胁迫处理第3d时,各处理的POD活性较Od上升幅度较小,但各补光处理的POD活性均高于对照,除R补光处理外,其他补光处理均达到显著性差异。亚低温胁迫处理第6d时,8R/2B、5R/5B、B和W补光处理下培育的番茄幼苗叶片中的POD活性相比亚低温胁迫开始时分别上升了129.01%、106.98%、113.69%和90.80%,且均高于对照(85.80%),其中以8R/2B补光处理下POD活性最高。但R和2R/8B补光处理下幼苗POD活性仅上升了80.28%和56.33%,上升幅度均低于对照。
2.5亚低温胁迫对番茄幼苗CAT活性的影响
图5显示,在亚低温胁迫处理期间,随着亚低温胁迫时间的延长,各处理的番茄幼苗叶片中CAT活性均呈上升趋势。在亚低温胁迫处理第3d时,各补光处理的CAT活性均高于对照,除R补光处理外,其他处理均达到显著性差异。而在亚低温胁迫处理第6d时,各处理培育的番茄幼苗叶片中CAT活性相较于亚低温胁迫开始时均有较大幅度的上升,其上升幅度具体表现为:8R/2B(135.74%)>5R/5B(133.13%)>B(125.74%)>2R/8B(117.50%)>R(115.8g%)>W(94.77%)>CK(87.41%)。
3讨论与结论
植物细胞的膜系统是其生命有机体的基础结构,往往是植物易受伤害的敏感部位,冻害或低温造成植物死亡本质的原因也是其生物膜被破坏[9]。在受到低温胁迫时质膜会遭到破坏,细胞膜选择透性能力减弱,导致胞内电解质外渗[10]。由此,可根据电解质渗透率的高低,来判断植株所受冻害程度。本研究结果表明,在亚低温胁迫处理期间,各补光处理所培育的番茄幼苗电解质渗透率均呈现上升趋势,其结果与王芳等[11]的研究结果相一致。其中未补光处理(CK)培育番茄幼苗叶片中电解质上升幅度最大,而以8R/2B补光处理培育的番茄幼苗叶片中电解质上升幅度最小。
丙二醛(MDA)作为膜质发生过氧化作用的最终产物之一,当MDA在植株体内积累到一定程度时,会导致电解质渗透率上升[12-15]。因此,不良环境会影响植物细胞膜系统发生膜质过氧化反应,MDA是植物在逆境中膜脂过氧化的产物,其含量的多少可作为细胞膜质损伤程度的参数,因此植物中丙二醛含量越低说明植物受伤害程度越小,生长越好,抗逆性越强。本试验结果表明,在亚低温胁迫处理下,各处理的MDA含量均增加,其结果与王萍等[16]的研究结果相一致。各补光处理培育的番茄幼苗在遇到亚低温胁迫时,幼苗叶片的MDA积累量均低于对照,其中以红蓝光8R/2B和蓝光B补光处理番茄幼苗在遇到亚低温胁迫时叶片中MDA积累量较低。
低温会导致植物细胞内活性氧代谢活动失衡,细胞内活性氧含量大量积累,使植物产生活性氧毒害。而抗氧化酶系统在清除自由基,防止活性氧伤害方面发挥着重要的作用[17-18]。其中SOD经常被认为是避免植物被白由基伤害的第一道防线,在保护酶系统中具有重要地位。POD是普遍存在所有植物组织中的一种氧化还原酶,在清除过氧化氢过程中具有一定作用,其活性高低与植物的耐冷性密切相关[19]。CAT是调节细胞内ROS的辅助酶,较高的CAT活性有利于ROS的清除[20]。本研究结果表明,在亚低温胁迫处理第6d时,各处理的抗氧化酶活性迅速升高,其结果与温日宇等[21]的研究结果相一致。与无补光CK相比,各补光处理下的番茄幼苗遇到亚低温胁迫时能够快速响应,在亚低温胁迫期间能一直维持着较高的抗氧化酶的活性。其中以红蓝光(8R12B)补光培育的番茄幼苗的抗氧化酶活性在遇到低温胁迫期间上升幅度最大。
综上所述,可以看出各种补光处理均可以提高番茄幼苗的耐冷性,其中以补充红蓝配比光(8R12B)所培育的番茄幼苗耐冷性最佳。
参考文献:
[1]KANGASJARVI S,NEUKERMANS J,LI S. et al Photosynthesis,photorespirationand light signalling in defence responses [J].Journalof Experimental Botany,2012,63(4):1619-1636.
[2]IZAGUIRRE M M,MAZZA C A,BIONDINI M. et al Remotesensing of future competitors: Impacts on plant defenses [J].Proceedings of the National Academy of Sciences of United States ofAmerica.2006,103(18): 7170-7174.
[3]杨有新,王峰,蔡加星,等.光质和光敏色素在植物逆境响应中的作用研究进展[J].园艺学报,2014,41(9):1861-1872.YANG Y X, WANG F.CAI J X,et al.ReCent Advances in the ROle ofLight Quality and Phytochrome in Plant Defense Resistance AgainstEnvironmental Stresses[J].Acta Horticulturae Sinica, 2014,41(9):1861-l872(in Chinese)
[4]FRANKLIN K A,wHITELAM G C.Light.quality regulation offreezing t01erance in Arabidopsis thaliana[J].Nature Genetics ,2007,39(11):1410-1413.
[5]蒲高斌.光质对番茄生育特性及其果实品质的影响[D].泰安:山东农业大学,2005.
PU G B Effects of Light Qualities on the Growth Characteristics andQualities of Tomato[D].Taian:Shandong Agriculture University,2005(in Chinese)
[6]许莉.光质对叶用莴苣生理特性及品质的影响[D].泰安:山东农业大学,2007.
XU L Effect of Light Quality on Physiological Characteristics andQuality of Leaf Lettuce[D].Taian:Shandong Agriculture University,2007.(in Chinese)
[7]陈建勋,王晓峰,植物生理学实验指导[M].广州:华南理工大学出版社,2015.
[8]王学奎,黄见良.植物生理生化实验原理与技术[M].北京:高等教育出版社,2015.
[9]鞏振辉.作物育种学[M].北京:农业出版社,2007.
[10]陈禹兴,付连双,王晓楠,等.低温胁迫对冬小麦恢复生长后植株细胞膜透性和丙二醛含量的影响[J].东北农业大学学报,2010,41(10):10-16.
CHEN Y X,FU L S,WANG X N,et al Effect of freezing stress onmembrane pemeability and MDA content in the re-growth plant Ofwinter Wheat culcivars[J].Journal of Northeast AgriculturalUniversity,2010,41(10):10-16.(in Chinese)
[11]王芳,李永生,彭云玲,等,外源一氧化氮对玉米幼苗抗低温胁迫的影响[J].干旱地区农业研究,2017,35(4):270-275.
WANG F,LI Y S,PENG Y L,et al.Effects Of exogenoUs nitric oxideon low temperature stress of maize seedlings[J]. AgriculturalResearch in the Arid Areas 2017,35(4):270-275.(in Chinese)
[12]DOBRETS0v G E,BORSCHEvSKAYA T A,PETROv v A,et al.The increaSeOf phOSphOlipid bilayer rigidity aRer lipidperoxidation[J].FEBS Letters,1977,84:195.
[13]周龙,廖康,王磊,等,低温胁迫对新疆野生樱桃李电解质渗出率和丙二醛含量的影响[J].新疆农业大学学报,2006,29(1):47-50.
ZHOU L,LIAO K,WANG L,et al. StUdy On Resistance to ColdneSSof Pmnus divaricate[J]. Journal of Xinjiang Agricultural University,2006,29(1):47-50.(inChinese)
[14]陈磊,朱月林,杨立飞,等.氮素不同形态配比对菜用大豆生长、种子抗氧化酶活性及活性氧代谢的影响[J].植物营养与肥料学报,2010,16(3):768-772.
CHEN L, ZHU Y L,YANG L F,et al.Effects of nitrogen forms andratios on plant growth, seed antioxidant enzyme activities and reactiveoxygen metabolism of vegetable soybean [J]. Journal of PlantNutrition and Fertilizer,2010,16(3):768-772.(in Chinese)
[15]张淑英,褚贵新,梁永超.不同铵硝配比对低温胁迫棉花幼苗生长及抗氧化酶活性的影响[J].植物营养与肥料学报,2017,23(3):721-729.
ZHANG S Y, CHU G X, LIANG Y C.Impacts of differentammonium/nitrate ratios on the growth and antioxidant defenseenzyme activities in cotton seedlings exposed to low-temperaturestress [J]. Journal of Plant Nutrition and Fertilizer,2017,23(3):721-729.(in Chinese)
[16]王萍,吕星光,李敏.低温胁迫对西瓜砧木幼苗生长和生理指标的影响[J].北方园艺,2017(10):35-38.
WANG P,LYU X G,LI M. Effects of Low Temperature Stress onGrowth and Physiological Indexes of Watermelon RootstockSeedlings [J].NorthernHorticulture,2017(10):35-38.(inChinese)
[17]IMAHORI Y. TAKEMURA M, BAI J.Chilling-induced oxidativestress and antioxidant responses in mume (Prumus mume) fruit duringlow temperature storage [J]. Postharvest Biology and Technology,2008(49):54-60.
[18] ASHRAF M. Biotechnological approach of improving plant salttolerance using antantioxidants as markers [J]. BiotechnologyAdvances,2009(27):84-93.
[19]馬骊,孙万仓,刘自刚,等.低温胁迫下白菜型与甘蓝型冬油菜抗寒基因表达差异[J].中国油料作物学报,2016, 32(3):135-142.
MA L, SUN W C,LIU Z G,et al. Expression of cold resistance genesfrom winter rapeseed of Brassica rapa and B napus under chillingstress [J]. Chinese Journal of Oil Crop Sciences,2016,32(3):135-142.(in Chinese)
[20]秦文斌,山溪,张振超,等.低温胁迫对甘蓝幼苗抗逆生理指标的影响[J].核农学报,2018,32(3):576-581.
QIN W B,SHAN X,ZHAN Z C,et al.Effect of Low TemperatureStress on Anti-stress Physiological Indexes of Cabbage Seedlings [J].Journal of Nuclear Agricultural Sciences,, 2018,32(3):576-581. (in Chinese)
[21]温日宇,刘建霞,李顺,等.低温胁迫对不同藜麦幼苗生理生化特性的影响[J].种子,2019,38(5):53-56.
WEN R Y, LIU J X. LI S, et al.Effect of Low Temperature Stress onPhysiological and Biochemical Characteristics of Different QuinoaSeedlings[J]. Seed,,2019, 38(5):53-56.(in Chinese)
(责任编辑:黄爱萍)
刘福长,杨玉凯,林碧英.不同光质夜间补光对番茄幼苗耐冷性的影响[J].福建农业学报,2020,35(11):1222-1227.
LIU F C,YANG Y K,LIN B Y. Effect of Nighttime Exposure to Light from Color Lamps on Cold Tolerance of Tomato Seedlings [J].Fujian Journalof Agricultural Sciences,2020,35(11):1222-1227.
收稿日期:2020-01-13初稿:2020-03-30修改稿
作者简介:刘福长(1962-),男,农艺师,研究方向:设施园艺(E-mail: 490835069@qq.com)
*通信作者:林碧英(1963-),女,教授,研究方向:蔬菜学与设施园艺(E-mail:lby3675878@163.com)
基金项目:福建省科技计划重大专项(208IN20002-2)
