水生植物对不同形态氮素污水的生理响应及相应脱氮效率
2020-02-22陈丹艳周威段婧婧薛利红冯彦房
陈丹艳 周威 段婧婧 薛利红 冯彦房

摘要:為了探究黄花水龙(Jussiaea stipulacea Ohwi)和铜钱草(Hydrocotyle verticillata)在不同形态氮素污水中的抗逆性和适应性,开展了水箱模拟试验,模拟农田径流污水[总氮(TN)质量浓度为8 mg/L,总磷(TP)质量浓度为1 mg/L],研究这2种水生植物对污水中TN、NO-3-N、NH+4-N的净化能力及其抗性生理响应,设置2种形态氮素:以NO-3-N为主(NO-3-N∶NH+4-N=4∶1)和以NH+4-N为主(NH+4-N∶NO-3-N=4∶1)。结果表明:(1)在2种形态氮素下2种水生植物处理对氮素去除率明显高于无水生植物的对照,其中黄花水龙对氮素去除率较高,在高NO-3-N和高NH+4-N形态下对TN的去除率分别为48%~69%和69%~91%;(2)对不同形态氮素污水,2种水生植物处理对NO-3-N的去除效果均高于NH+4-N,其中黄花水龙在高NO-3-N和高NH+4-N形态下对NO-3-N的去除率分别高达58%~66%和79%~91%,而对NH+4-N的去除率仅为-65%~25%和21%~62%;(3)夏季试验期间,高质量浓度氮素增加了黄花水龙和铜钱草的氧化胁迫作用,2种水生植物通过调节体内抗氧化酶机制来响应。铜钱草体内的超氧化物歧化酶(SOD)和过氧化物酶(POD)活性在不同形态氮素下除第1周外均高于黄花水龙,表明在外界高质量浓度氮条件下,铜钱草受到了较大的胁迫;高NO-3-N形态下黄花水龙的蛋白质和叶绿素含量在试验过程中均高于铜钱草,表明在高硝氮污水胁迫下黄花水龙耐受性更好。
关键词:黄花水龙;铜钱草;氮素形态;生理响应;抗氧化酶
中图分类号:X592文献标识码:A文章编号:1000-4440(2020)06-1468-07
Abstract:In order to explore the stress resistance and adaptability of Jussiaea stipulacea Ohwi and Hydrocotyle verticillata in sewage with different forms of nitrogen, a water tank experiment was carried out. Using simulated farmland runoff wastewater(the mass concentration of total nitrogen was 8 mg/L, the mass concentration of total phosphorus was 1 mg/L),two forms of nitrogen were set:mainly NO-3-N(NO-3-N∶NH+4-N=4∶1) and NH+4-N(NH+4-N∶NO-3-N=4∶1),to study the purification ability of these two aquatic plants to TN, NO-3-N and NH+4-N in sewage and its resistance physiological response. The results showed that under the two forms of nitrogen, the nitrogen removal rates in the two aquatic plants treatments were significantly higher than those in the control, and the nitrogen removal rate of Jussiaea stipulacea Ohwi was the highest, and the removal rates of TN were 48%-69% and 69%-91% under high NO-3-N and high NH+4-N conditions, respectively. In wastewater with different forms of nitrogen, the removal efficiency of NO-3-N in two aquatic plants treatments was higher than that of NH+4-N. The NO-3-N removal rates of Jussiaea stipulacea Ohwi were 58%-66% and 79%-91% under the conditions of high NO-3-N and high NH+4-N, respectively, while the removal rates of NH+4-N were -65%-25% and 21%-62%, respectively. The high concentration of nitrogen increased the oxidative stress of Jussiaea stipulacea Ohwi and Hydrocotyle verticillata. The two aquatic plants responded to the stress by regulating the antioxidant enzyme mechanism. The activities of superoxide dismutase(SOD)and peroxidase(POD)in Hydrocotyle verticillata were higher than those in Jussiaea stipulacea Ohwi in each cycle except the first week, indicating that under high nitrogen conditions, Hydrocotyle verticillata was under greater stress. Under the conditions of high NO-3-N, the contents of protein and chlorophyll in Jussiaea stipulacea Ohwi were higher than those in Hydrocotyle verticillata during the experiment, indicating that the Jussiaea stipulacea Ohwi was more tolerant under high nitrate wastewater stress.
Key words:Jussiaea stipulacea Ohwi;Hydrocotyle verticillata;nitrogen form;physiological response;antioxidant enzyme
面源污染,亦即非点源污染(Non-point source pollution),是相对于排污点集中、排污途径明确的点源污染而言的。农业面源污染主要包括农用化学品污染(化肥、农药等)、集约化养殖场污染、农村生活污水污染等方面[1]。氮、磷作为植物生长的必需养分,在农业生产中被广泛应用,未被植物吸收的氮、磷随降雨和灌溉渗入地表水和地下水,造成水体污染和富营养化[2-3],对自然环境和人类社会产生极大的危害。水生植物修复技术适合于城市生活污水的第二级处理、风景区景观水体处理和湖泊河流修复[4]。利用生态湿地塘中水生植物对农田流失的氮、磷进行净化和去除,是当前削减农田污染、缓解水体富营养化的重要途径。而目前国内对于水环境治理和修复的研究起步较晚,对水生植物的研究多集中于对氮、磷等污染物的去除能力以及植物自身生物量的增长等方面,对不同形态氮素污水引起的植物生理响应方面的研究较少。
铜钱草和黄花水龙是江浙一带常见的水生植物,因其具有良好的适应性,很多研究者将其用于富营养化水体修复。陈友媛等[5]研究结果显示,铜钱草对富营养化水体具备较好的净化效果,其对总氮、铵态氮和总磷的去除率分别达到65.1%、95.0%和90.0%以上。常会庆等[6]研究结果表明黄花水龙对总氮、铵态氮、硝态氮和总磷的去除效率分别为62.9%、90.83%、53.90%和71.21%,且与对照相比去除率达到显著水平(P<0.05)。邵凯迪等[7]研究结果表明在低NO-3-N质量浓度下(4.76~5.09 mg/L),黄花水龙和铜钱草在试验初期对TN和NO-3-N去除率分别超过80%和90%,在试验后期黄花水龙仍保持较高的NO-3-N去除率(90%以上);在高NO-3-N质量浓度下(13. 19~13. 96 mg/L),黄花水龙对氮去除能力显著高于铜钱草。
目前,用于农业面源污染治理的水生植物接纳不同形态氮素污水的生理响应方面的研究相对较少。如何科学地选择具有一定抗氮、磷胁迫能力的水生植物对农业面源污染治理非常重要。在受到高氮等外界不利环境影响下,水生植物因其机理的差异,产生了不同的生理代谢和生化反应来减少对自身的伤害,如调节渗透压、光合作用的变化、抗氧化调节机制和内源激素调节等[8]。熊汉锋等[9]研究发现苦草叶组织的过氧化物酶(POD)活性、超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量均随氮、磷质量浓度的增加而上升。研究水生植物对不同形态氮素的生理响应有助于了解植物的生理特性,从而筛选到用于面源污染治理的适宜的水生植物。
因此,本试验在不同形态氮素污水处理下研究2种水生植物对氮素的去除效率及其蛋白质、叶绿素、丙二醛含量和过氧化物酶、超氧化物歧化酶活性等生理指标变化,以期为农业面源污染治理中植物选择提供参考。
1材料与方法
1.1试验材料
黄花水龙(Jussiaea stipulacea Ohwi),柳叶菜科丁香蓼属,多年生浮叶植物;铜钱草(Hydrocotyle verticillata),天胡荽亚科天胡荽属,多年生匍匐草本植物。2种植物均采集于江苏省农业科学院水生植物培育池,选取生长状况良好的植株。试验所用的模拟污水用硝酸钠、氯化铵和磷酸二氢钾人工配制。试验容器采用周转箱,其内部长、宽、高分别为755 mm、520 mm、500 mm。
1.2试验方法
试验场地选取江苏省农业科学院大棚科研试验基地,依次排放18个周转箱。试验中氮、磷模拟质量浓度:总氮(TN)质量浓度为8 mg/L,总磷(TP)质量浓度为1 mg/L。设置3个处理,包括2个水生植物处理和1个无植物的对照,每个处理分别设3个重复,分别培养在高NO-3-N(NO-3-N∶NH+4-N=4∶1)和高NH+4-N(NH+4-N∶NO-3-N=4∶1)污水中,每箱污水109 L。铜钱草每箱栽种35 g左右,黄花水龙每箱栽种6株,约为60 g左右。从2019年7月25日开始,到2019年8月28日结束。正式试验从8月1日开始,每14 d为一个阶段,中间换1次水。水样每1 d取1次,每次取样时使用五点取样法取100 ml水样,保存于4 ℃冰箱中。同时在每次取样时原位测定溶解氧(DO)浓度、酸碱度(pH)和温度。每7 d每箱取出相同质量的植物样品,用自来水洗净沥干后,用锡纸包裹后保存于-30 ℃冰箱中。
1.3指标测定
水温、溶解氧(DO)浓度和pH使用意大利哈纳HI9829便携式水质测定仪进行现场测定。水体中NO-3-N、NH+4-N和TN浓度使用荷兰SKALAR SAN+SYSTEM流动分析仪测定。植物生理指标(蛋白质含量、叶绿素含量、过氧化物酶活性、丙二醛含量和超氧化物歧化酶活性)使用南京建成生物工程研究所生产的试剂盒测定。
每一阶段水体中TN、NO-3-N和NH+4-N的去除率使用以下公式计算:
1.4数据分析
采用WPS及SPSS19.0进行数据处理,采用Duncans法进行差异显著性分析。
2结果与分析
2.12种水生植物在不同形态氮素下的脱氮效率
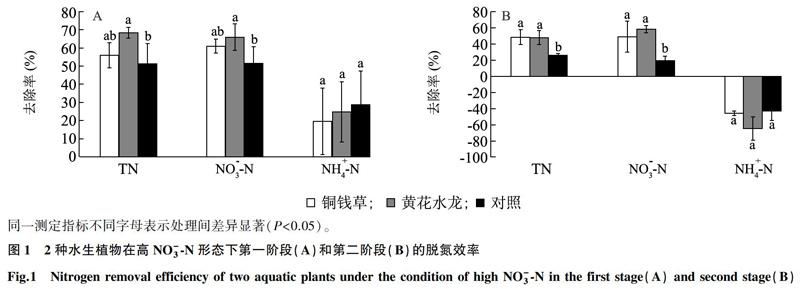
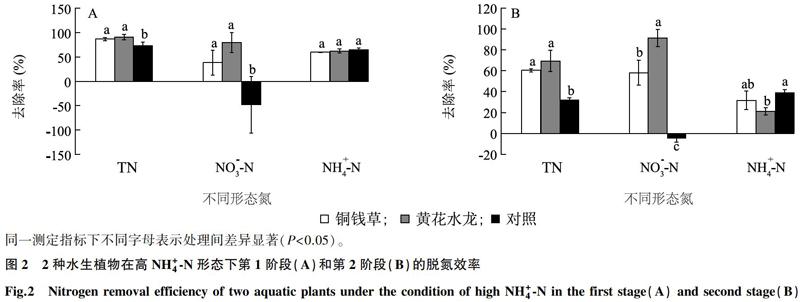
黄花水龙和铜钱草2种水生植物在高NO-3-N形态下的脱氮效率如图1所示。试验第1阶段黄花水龙处理对TN和NO-3-N的去除率与铜钱草处理无显著差异,但均显著高于对照,第2阶段2种水生植物处理对TN和NO-3-N的去除率均显著高于对照。2个试验阶段2种水生植物处理间对NH+4-N的去除率均没有显著差异。2种水生植物处理对TN、NO-3-N和NH+4-N的去除率第1阶段均高于第2阶段。其中铜钱草对TN、NO-3-N和NH+4-N的去除率分別为49%~56%、49%~61%和-46%~20%,黄花水龙分别为48%~69%、58%~66%和-65%~25%。
2种植物在高NH+4-N形态下的脱氮效率如图2所示。2种水生植物处理在2个阶段对TN和NO-3-N的去除率均高于对照,其中第2阶段对NO-3-N的去除率黄花水龙处理显著高于铜钱草处理。2种水生植物处理间在2个阶段对NH+4-N的去除率无显示差异。铜钱草对TN、NO-3-N和NH+4-N的去除率分别为60%~87%、38%~58%和32%~60%,黄花水龙分别为69%~91%、79%~91%和21%~62%。
2.22种水生植物在不同形态氮素下蛋白质和叶绿素含量变化
在不同形态氮素下2种水生植物蛋白质含量如图3所示,第2阶段的蛋白质含量低于第1阶段,与第1阶段相比下降了77%~89%,表明换水后2种水生植物生长受到严重胁迫。高NO-3-N形态下第1周2种水生植物蛋白质含量差异显著,高NH+4-N形态下2种水生植物蛋白质含量没有显著差异。
如图4所示,叶绿素含量与蛋白质含量的变化类似,2种水生植物第2阶段的叶绿素含量均低于第1阶段,表明在第2阶段植物生长受到严重胁迫。高NO-3-N形态下黄花水龙叶绿素含量均高于铜钱草,高NH+4-N形态下2种水生植物叶绿素含量无显著差异。高NO-3-N形态下第1阶段黄花水龙和铜钱草的叶绿素含量分别下降了71%和79%;第2阶段铜钱草叶绿素含量下降了37%,黄花水龙叶绿素含量没有变化。高NH+4-N形态下第1阶段铜钱草叶绿素含量下降了14%,而黄花水龙叶绿素含量上升了9%;第2阶段铜钱草叶绿素含量上升了49%,黄花水龙叶绿素含量下降了31%。第1周不同氮素形态下黄花水龙的叶绿素含量表现出显著差异,第4周高NO-3-N形态下2种水生植物叶绿素含量呈现显著差异。
2.32种水生植物在不同形态氮素下抗氧化酶活性变化
超氧化物歧化酶(SOD)是活性氧清除系统中第1个发挥作用的抗氧化酶,是细胞防御活性氧毒害作用的第1道防线。如图5所示,第1阶段2种水生植物在不同形态氮素下随试验时间增加体内SOD活性均有所下降,第2阶段2种水生植物在不同形态氮素下随试验时间增加体内SOD活性变化不大。第1阶段在相同形态氮素下2种水生植物表现出显著差异,第2阶段在相同形态氮素下2种水生植物只在第3周表现出显著差异。
2种水生植物过氧化物酶(POD)活性的变化如图6所示。从图6中可以看出,换水后POD活性显著增加。第1阶段铜钱草在2种形态氮素下的POD活性增加率高于黄花水龙;第2阶段铜钱草在2种形态氮素下的POD活性下降高于黄花水龙。第1阶段铜钱草POD活性在不同形态氮素下无显著差异,而黄花水龙只在第1周差异显著。高NO-3-N形态下2种水生植物POD活性在第2周和第3周差异显著,高NH+4-N形态下2种水生植物POD活性除第4周外均有显著差异。
2.42种水生植物在不同形态氮素下丙二醛活性的变化
丙二醛(MDA)是膜脂过氧化的重要产物,作为膜脂过氧化指标,其含量的变化可反映逆境条件下膜系统受伤害的程度。2种水生植物MDA含量变化如图7所示,相同浓度下2种水生植物的MDA含量比较一致,只有在第1周高NO-3-N形态下和第4周高NH+4-N形态下2种水生植物表现出显著差异。第1阶段高NO-3-N形态下黄花水龙和铜钱草的MDA含量均下降,分别下降80%和81%;高NH+4-N形态下2种水生植物则均上升了41%。第2阶段高NO-3-N形态下黄花水龙和铜钱草的MDA含量分别下降了38%和43%,高NH+4-N形态下2种水生植物则分别下降了5%和94%。
3讨论
3.1沟渠植物水质净化效果及影响因素
沟渠植物不但可以通过吸收作用去除氮、磷等污染物,还可以通过根系释放氧气和分泌物加速氮的转化。本试验于8月份进行,为沟渠植物的生长旺盛期,对模拟污水有较好的净化效果。总体而言,2种水生植物对TN、NO-3-N和NH+4-N的最高去除效率分别为91%、91%和62%,且均在高NH+4-N氮素形态下去除率达到最大值。不同植物种类在营养吸收能力、根系分布、氧气释放量、生物量和抗逆性方面存在差异,从而对污水的净化效率不同[10]。在本试验高氮浓度下铜钱草出现叶片发黄、生物量减少的现象,其对TN、NO-3-N和NH+4-N的去除率相对较低;相比而言,黄花水龙在试验过程中生长状况较好,收获时生物量增加了2倍以上,在高NO-3-N和高NH+4-N下对氮素的去除能力较高(对TN的去除率分别为48%~69%和69%~91%,对NO-3-N的去除率分别为58%~66%和79%~91%),表明在这2种形态氮素下黄花水龙与铜钱草相比有着更好的适应性。在本试验2种形态氮素下,黄花水龙对TN和NO-3-N的去除率均高于对照,这表明黄花水龙对以NO-3-N为主要形态氮素的污水具有较大的消纳潜力,这与王超等[11]的研究结果相一致。植物残体分解后给藻类提供了基质,附植藻类开始产生,进一步威胁到植物的生存,进而影响植物对氮素的去除。本研究中铜钱草和黄花水龙在第2周最后就有附植藻类产生。已有研究结果表明,附植藻类会在沉水植物表面形成一层膜,阻碍植物对养分的吸收并且减弱光照度,進而降低水生植物的光合作用,对沉水植物的生长产生不利影响[12-13]。
沟渠植物可以通过光合作用增加水体中溶解氧的侵蚀深度,创造出好氧-厌氧的微环境,从而促进微生物的硝化/反硝化作用[14]。黄花水龙在试验期间茎部粗壮,许多沉没于水中的茎部也生长出发达的不定根系,这为微生物提供了良好的附着场所。其叶片和茎秆大量浮出水面,从一定程度上削减了水面以下光照度,从而使沉没于水中部分的光合作用强度减弱,促进了水面以下水生植物根际区域厌氧界面的生成,为微生物的反硝化作用提供更加适宜的环境。无植物的对照处理也具有一定的氮素去除能力,这主要是其中生长的藻类及反硝化微生物共同作用的结果[15]。陈友媛等[5]研究发现,在TN质量浓度为6.70 mg/L,以NH+4-N为主要氮素,铜钱草对水中TN、NO-3-N和NH+4-N的去除率分别为65.1%、47.1%和95.0%。而在本试验中,在2个主要形态氮素下2种水生植物组对NO-3-N的去除率均较高,特别是黄花水龙,在高NO-3-N形态和高NH+4-N形态下对NO-3-N去除率分别在58%和79%以上,显著高于对照,反硝化作用在其中的影响不可忽视。
除了植物种类外,影响氮素去除率的因素还有很多,如温度、pH值、溶解氧含量、污水浓度、氮素形态等[16-18]。前人对于污染物浓度和去除效率之间的关系有过许多研究。袁东海等[16]认为人工湿地净化对污水中污染物的初始浓度有一定的要求,污染物较低情况下人工湿地净化效果较好,污染物浓度较高情况下其净化效果下降。净水效果存在铵态氮、硝铵态氮浓度阈值,阈值之内净水效果与浓度呈正相关关系,阈值因植物不同而异;水体环境氮素浓度达到一定阈值时,水生植物会受到逆境胁迫,其正常的生理活动会受到影响,去除能力亦受到限制[17]。
3.2沟渠植物对氮素胁迫的生理响应机制
20世纪60年代末,Fridovieh提出生物自由基伤害假说,植物在逆境条件下,细胞内产生过量自由基,这些自由基能引发膜脂过氧化作用,造成膜系统的伤害[19]。自由基还能干扰植物细胞的光合、呼吸及其他代谢过程,严重时会导致植物细胞死亡[20-21]。超氧化物歧化酶(SOD)和过氧化物酶(POD)是保护系统的主要酶[22]。当植物受到高浓度氮素胁迫时,SOD和POD通过协调作用,防御细胞膜过氧化,从而减轻植物受害程度。当植物受到的胁迫越严重时,体内SOD和POD活性越高,对逆境的耐受性也就越强。铜钱草体内的SOD和POD活性在高氮素条件下显著上升,说明铜钱草受到了较大的环境胁迫,且SOD和POD在其抵抗环境胁迫中可能扮演重要角色。
本试验是在江苏省农业科学院大棚科研试验基地进行的,夏季白天温度最高可达45 ℃。研究发现,在40 ℃高温胁迫下铜钱草植株叶片变黄,叶绿素含量、Fv/Fm、qP和ETR等参数值下降,光曲线呈现不规律变化[23]。本试验中,在高温和高氮污水的双重作用下,尤其是高NO-3-N处理时黄花水龙和铜钱草体内的膜脂过氧化加剧,生长受到胁迫。
丙二醛(MDA)是高活性的脂过氧化物,能交联脂类、核酸、糖类及蛋白质,逆境条件下其在细胞中的积累常造成质膜受损[24]。在本试验中,在换水前的2周,铜钱草的MDA含量在不同形态氮素下均高于黄花水龙,表明这一阶段铜钱草受到严重胁迫。植物体内的蛋白质大多数是参与各种代谢的酶类,其含量是了解植物体总代谢的一个重要指标[25]。而在逆境条件下,不同植物体内蛋白质的合成会表现出一定的差异[26]。因此,可用植物体内的可溶性蛋白质含量判定污水对植物生理的影响情况。本试验高NO-3-N形态下黄花水龙的蛋白质含量在试验第1阶段中显著高于铜钱草,表明在这一阶段高NO-3-N污水胁迫下,黄花水龙的耐受性更好。
光合作用被认为是植物对环境变化反应最为敏感的生理过程之一[27-29],而叶绿素是光合作用中完成光能吸收的主要色素[30]。高盐胁迫抑制叶绿素合成关键酶的活性,激活叶绿素酶,从而加速叶绿素的降解[31]。但有研究结果表明水体中较高的氮、磷浓度对水生植物而言,与盐胁迫、环境污染物胁迫一样是一种逆境胁迫,影响其正常生理活动[10,22,25]。在本试验中,高NO-3-N形态下黄花水龙的叶绿素含量在试验过程中均高于铜钱草,同样表明黄花水龙受到的环境胁迫影响较小。
综上所述,黄花水龙和铜钱草在接纳2种形态氮素污水时对氮素的去除率明显高于无植物对照,且黄花水龙对氮素去除率较高。与对照相比,在高NO-3-N和高NH+4-N形态下2种水生植物对氮素,特别是NO-3-N和TN的去处效率均较高。试验期间,高浓度氮素增加了黄花水龙和铜钱草的氧化胁迫作用,2种水生植物通过调节体内抗氧化酶机制响应氧化胁迫。在外界高氮浓度条件下,铜钱草受到的胁迫较大,而黄花水龙的耐受性较好。
参考文献:
[1]杨林章,冯彦房,施卫明,等. 我国农业面源污染治理技术研究进展[J].中国生态农业学报,2013,21(1):96-101.
[2]欧阳威,蔡冠清,黄浩波,等. 小流域农业面源氮污染时空特征及与土壤呼吸硝化关系分析[J].环境科学,2014,35(6):2411-2418.
[3]宋立芳,王毅,吴金水,等. 水稻种植对中亚热带红壤丘陵区小流域氮磷养分输出的影响[J].环境学,2014,35(1):150-156.
[4]张芳,易能,张振华,等. 不同类型水生植物对富营养化水体氮转化及环境因素的影响[J].江苏农业学报,2015,31(5):1045-1052.
[5]陈友媛,崔香,董滨,等. 3种水培观赏植物净化模拟污水的试验研究[J].水土保持学报,2011,25(2):253-257.
[6]常会庆,王世华,徐晓峰,等. 两种水生植物对重富营养化水体修复效果研究[J].水土保持研究,2012,19(5):262-265.
[7]邵凯迪,段婧婧,薛利红,等. 5种水生植物对模拟菜地径流中总氮和硝氮凈化效果[J].环境工程技术学报,2020,10(3):406-413.
[8]李敏,张健,李玉娟,等. 植物耐盐生理及耐盐基因的研究进展[J]. 江苏农业科学,2012,40(10):45-49.
[9]熊汉锋,谭启玲,刘艳玲. 不同营养状态下苦草的生理响应[J].华中农业大学学报,2009,28(4):442-445.
[10]孙瑞莲,刘健. 3种挺水植物对污水的净化效果及生理响应[J].生态环境学报,2018,27(5):926-932.
[11]王超,张文明,王沛芳,等. 黄花水龙对富营养化水体中氮磷去除效果的研究[J].环境科学,2007(5):975-981.
[12]宋玉芝,黄瑾,秦伯强. 附着生物对太湖常见的两种沉水植物快速光曲线的影响[J].湖泊科学,2010,22(6):935-940.
[13]张艳娜,宋玉芝,王敏,等. 附植藻类对沉水植物狐尾藻生长及生理的影[J].环境科学学报,2015,35(7): 2282-2288.
[14]刘丹丹,李正魁,叶忠香,等. 伊乐藻和氮循环菌技术对太湖氮素吸收和反硝化的影响[J].环境科学,2014,35(10):3764-3768.
[15]刘志迎,许海,詹旭,等. 蓝藻水华对太湖水柱反硝化作用的影响[J].环境科学,2019,40(3):1261-1268.
[16]袁东海,高士祥,任全进,等. 几种挺水植物净化生活污水总氮和总磷效果的研究[J].水土保持学报,2004,18(4):77-80,92.
[17]陆庆楠,贺宇欣,莊文化,等. 粉绿狐尾藻净水效果对氮磷浓度的响应机制[J].中国农村水利水电,2019(2):11-15.
[18]刘凤梅. 温度对凤眼莲浮岛净化效果影响的研究[J].环境保护科学,2013,39(3):9-11,54.
[19]FRIDOVIEH I. The biology of oxygen radical[J].Science,1975,201:875-880.
[20]李合生. 植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[21]BOWLER C. Superoxide dismutase and stress tolerance[J]. Plant Molecular Biology,1992,43:83-91.
[22]熊汉锋,谭启玲,李伟,等. 不同营养状态下3种沉水植物的生理响应[J].海洋湖沼通报,2009(4):62-66.
[23]张柳. 南美天胡荽对光温胁迫的适应性及园林应用研究[D].长沙:中南林业科技大学,2014.
[24]HARE P D,CRESS W A,VAN S J. Dissecting the roles of osmolyte accumulation during stress[J].Plant Cell Environment,1998,21:535-553.
[25]付晓云,梁茵,黄彦青. 污染水体中水生植物的生理效应研究[J].西北林学院学报,2013,28(1):63-66,72.
[26]沙翠芸,孟庆瑞,王静,等. 两种彩叶植物对铅胁迫的生理响应[J]. 西北林学院学报, 2011, 26(4):36-40.
[27]李雅洁,张其安,陆晓民. 不同外源物质对低温弱光次生盐渍化复合逆境下黄瓜幼苗生长、抗氧化系统及光合作用的影响[J]. 江苏农业学报,2018,34(2):404-410.
[28]袁孟玲,岳堃,王红,等.增强UV-B辐射对芒果成年树光合作用及其产量与常规品质的影响[J].南方农业学报,2018, 49(5):930-937.
[29]高祺,李明,朴琳,等. 拔节期弱光和渍水胁迫对春玉米光合作用、根系生长及产量的影响[J].江苏农业学报,2018,34(6):1276-1286.
[30]朱新广,张其德. NaCl对光合作用影响的研究进展[J].植物学通报,1999,16(4):332-338.
[31]刘凤歧,刘杰淋,朱瑞芬,等. 4种燕麦对NaCl胁迫的生理响应及耐盐性评价[J].草业科学,2015,24(1):183-189.
(责任编辑:张震林)
