利用Illumina MiSeq测序平台分析沐川乌骨黑鸡空肠微生物多样性
2020-02-22廖娟王钢喻世刚梁梓
廖娟 王钢 喻世刚 梁梓
摘要:为探索沐川乌骨鸡肠道微生物多样性,采用Illumina MiSeq测序平台对10羽沐川乌骨鸡的空肠内容物样本中微生物的16S rDNA-V4变异区进行测序,利用UPARSE等软件分析和统计样品中的操作分类单元(OTUs)数量,并进行物种注释及丰度分析,揭示样品的物种构成。结果表明,共获得667 466条tags,在97%相似水平上进行OTU分类,被归类于3 154个OTUs,且由稀释曲线可知此次测序结果较全面地覆盖了沐川乌骨黑鸡空肠微生物群落。门水平上,占优势的门主要有厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、变形菌门(Proteobacteria),但这3种优势菌门在不同样品中所占比例均有所差异。在属水平上,在10个样品中所占比例均大于1%的菌属有乳杆菌属(Lactobacillus)和Romboutsia菌属。
关键词:沐川乌骨黑鸡;Illumina MiSeq测序;空肠;微生物;多样性
中图分类号: S831;S182 文献标志码: A 文章编号:1002-1302(2020)24-0178-04
在动物的身体部位中,胃肠道中定殖着数量庞大、种类繁多的微生物,这类复杂的微生物群落被称为肠道微生物[1]。肠道微生物菌群在宿主营养、发育、免疫及代谢等过程中起着重要作用[2-3]。近年来,肠道微生物多样性成为很多学者的研究热点,肠道微生物多样性的研究方法较多,早期主要采用传统的纯培养方法[4],但是肠道中有很多厌氧型细菌,传统纯培养方法无法分离培养。随着科学技术的发展,末端限制性片段长度多样性分析、变性梯度凝胶电泳、温度梯度凝胶电泳等技术广泛运用于肠道微生物的分析中,这些方法虽能检测一些厌氧型微生物,但这些研究技术获得的信息覆盖量仍不能充分反映出肠道微生物的多样性。Illumina MiSeq技术因其方便、快速、准确率高和信息覆盖量大等优点,被广泛应用于兔[5]、人[6]、蜜蜂[7]等的肠道微生物研究中。林奕岑等[8]、徐帅等[9]采用Illumina MiSeq技术分别对普通鸡的盲肠和回肠的微生物多样性进行分析,但对沐川乌骨黑鸡肠道微生物分析尚未见报道。沐川乌骨黑鸡是四川省乐山市的优良地方品种资源,在沐川县具有悠久的种源历史,属药肉兼用型品种,因其全身乌黑而得名[10],其生产性能及健康程度与复杂的肠道微生物组成密切相关。本研究利用Illumina MiSeq测序平台分析了沐川乌骨黑鸡空肠微生物多样性,旨在为相关研究提供理论依据。
1 材料与方法
1.1 试验动物
试验采用1日龄健康的沐川乌骨黑母鸡,购于四川省乐山市沐川县黑凤凰乌骨鸡业有限公司,采用相同的饲养和管理方式。
1.2 试验方法
1.2.1 样品采集 于30日龄时,随机选取10羽健康个体,无菌操作采集每羽沐川乌骨黑鸡的空肠内容物,依次编号K1~K10,并快速置于液氮罐中,带回实验室放入-80 ℃冰箱保存。
1.2.2 总DNA提取 采用SDS方法提取空肠内容物样本基因组DNA,利用1%琼脂糖凝胶电泳检测DNA的纯度和浓度,采用无菌水稀释至1 ng/μL。
1.2.3 基因文库构建及上机测序 以稀释后的样品基因组 DNA 为模板,使用16S rRNA-V4区特异性引物515F(5′-GTGCCAGCMGCCGCGGTAA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)进行PCR扩增。PCR产物采用2%琼脂糖凝胶进行电泳检测后,切下目标条带,用Thermo Scientific 公司GeneJET胶回收试剂盒进行回收。
使用建库试剂盒(Thermofisher公司的Ion Plus Fragment Library Kit 48 rxns建库试剂盒)进行文库的构建,构建好的文库经过定量、检测合格后,使用Thermofisher的Ion S5TMXL进行上机测序。测序工作在诺禾致源生物信息有限公司完成。
1.3 生物信息学分析
测序完成后,原始数据使用Cutadapt对低质量部分进行剪切[11],再根据Barcode从得到的reads中拆分出各样品数据,截去Barcode和引物序列得到的原始数据(raw reads)后,利用UCHIME Algorithm 软件去除嵌合序列[12],得到有效数据(clean reads)。利用Uparse软件对所有样品的全部clean reads按97%的相似性进行操作分类单元(operational taxonomic unit,简称OTU)聚类。并对用Mothur方法与SILVA的SSUrRNA数据库进行物种注释分析[13-14]。采用Qiime软件计算每个样品的Alpha多样性。
2 结果与分析
2.1 各样品序列及OTUs信息
本试验基于Illumina MiSeq测序平台对10羽沐川乌骨黑鸡的空肠内容物菌群V4区进行了测序,一共获得667 466条tags,在97%的相似水平上进行OTU分类,被归类于3 154个OTUs。其中,tags数量最高的是样品K3,为86 261条;tags数量最低的是样品K9,为47 679条。
2.2 Alpha多样性分析
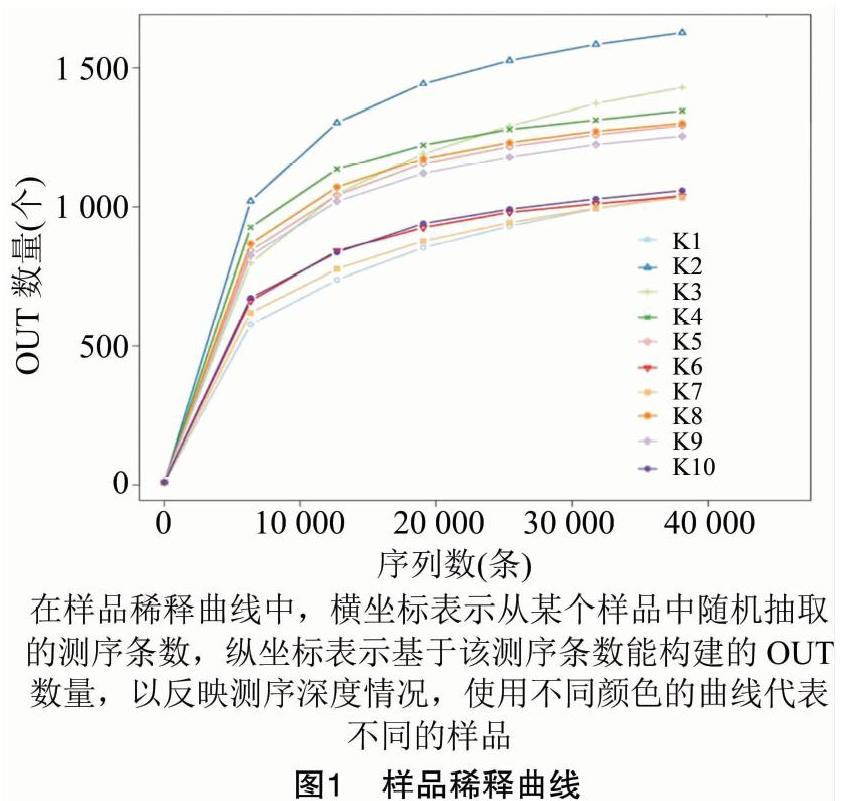
采用Alpha多样性指数分析样品内的微生物群落的丰富度和多样性。从样品稀释曲线(图1)可知,随着测试深度的不断加深,稀释曲线逐渐趋于平坦,说明测序数据量渐进合理,10个样本的覆盖率(good's coverage)均达到了99.0%以上,表明更多的数据量仅会产生少量新OTUs,16S rRNA基因测序深度足以反映样品的微生物多样性。
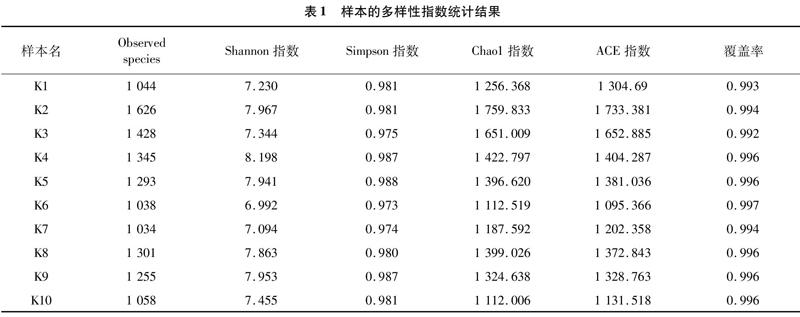
采用Chao1、ACE、Shannon、Simpson等指数表示,样品中微生物群落的Alpha多样性。Chao1或ACE指数越大,表明样品群落丰富度越高;Shannon指數越高,表明群落物种多样性越高;Simpson指数越高,表示其群落均匀度越高。由表1可知,样品K2物种丰富度相对较高,样品K4物种多样性相对较高。
3.3 物種注释
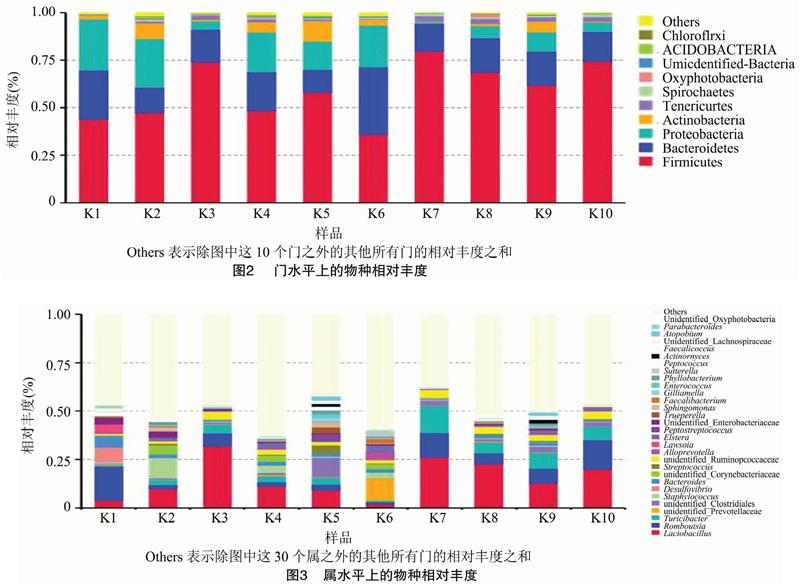
根据物种注释结果,选取每个样品在门水平上最大丰度排名前10的物种和在属水平上最大丰度排名前30的物种,生成物种相对丰度柱形累加图,以便直观查看各样品在门和属分类水平上相对丰度较高的物种及其比例。由图2和图3可知,10个样品共获得45个菌门,559个菌属。
门水平上,占优势的门主要有厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、变形菌门(Proteobacteria),这3个菌门总比例在每个样品中均在84%以上,但这3个优势菌门在不同样品中所占比例有所差异。厚壁菌门在10个样品中所占比例均最高,均高于35%,其中K7样品中高达79%。
在属水平上,共获得559个菌属,其中56个属尚未被注释出具体的菌属名。在10个样品中所占比例均大于1%的菌属有乳杆菌属(Lactobacillus)和Romboutsia菌属。不同菌属在不同样品中所占比例差异较大,其中,样品K1和K6与其他样品差异最为明显。
3.4 聚类分析
为研究不同样品间的相似性,基于Weighted Unifrac距离采用非加权组平均法(UPGMA)对样品进行聚类分析。由图4可知,K1和K6这2个样品的菌群与其他样品组成差异较大。
3 讨论
传统的分离培养方法仅限于分离能在培养基上生长的微生物,而通过该方法鉴定的微生物仅占所研究样品中所有微生物物种的1%~10%[15]。因此,传统的分离培养技术分析所得到的结果并不能反映所研究样品中微生物的多样性。近年来,Illumina MiSeq测序技术广泛应用于各种动物肠道微生物多样性的研究,该技术可在短时间内获得大量数据,而且检测数据完整性好[16]。
本研究采用Illumina MiSeq测序技术对沐川乌骨黑鸡空肠微生物菌群进行分析,样品稀释曲线分析表明,10个样本的覆盖率均达到99.0%,说明此次测序数据比较全面,能够比较全面地反映10个研究对象的空肠微生物组成情况。但10羽沐川乌骨黑鸡即使在相同的饲养和管理方式下,其空肠微生物的种类和数量也不完全相同,这可能与每个检测
个体的宿主属性(如身体情况、饮食习惯、自身激素水平等)有关[2]。
有关肠道微生物的研究均表明,厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、变形菌门(Proteobacteria)为相对丰度最高的菌门[8-9,17],本研究结果与之一致。Torok等进行了肉鸡饲喂试验,结果表明厚壁菌门、拟杆菌门、变形菌门与肉鸡生产性能相关[18]。厚壁菌门在健康的火鸡[19]、肉鸡[9]、蛋鸡[17]的肠道中也是含量最多的菌群,厚壁菌门与粗饲料的消化吸收有关,对宿主体内的物质和能量代谢有重要作用。拟杆菌菌群发酵产物有醋酸盐,可以刺激宿主黏液相关基因的表达,促进肠道黏液的产生[20]。
在物种注释结果中属水平上有56个属尚未被注释出具体的菌属名,这表明有关鸡肠道微生物在属水平上的研究还不够深入。肠道微生物是一个十分复杂的系统,即使同一批在相同的饲养和管理下的鸡群,不同菌属在不同个体肠道中所占比例差异也较大,每种菌属在宿主的新陈代谢过程中均发挥着不同作用。随着高通量测序的广泛应用,对肠道微生物的研究不断深入,肠道微生物的功能研究将成为热点,筛选出可提高宿主生产性能的有益微生物,对提高养殖经济效益具有重要意义。
参考文献:
[1]Gerritsen J,Smidt H,Rijkers G T,et al. Intestinal microbiota in human health and disease:the impact of probiotics[J]. Genes & Nutrition,2011,6(3):209-240.
[2]Hanning I,Diaz-Sanchez S. The functionality of the gastrointestinal microbiome in non-human animals[J]. Microbiome,2015,3(1):51.
[3]Bergmann G T. Microbial community composition along the digestive tract in forage-and grain-fed bison[J]. BMC Veterinary Research,2017,13(1):253.
[4]Barnes E M,Mead G C,Barnum D A,et al. The intestinal flora of the chicken in the period 2 to 6 weeks of age,with particular reference to the anaerobic bacteria[J]. British Poultry Science,1972,13(3):311-326.
[5]Zeng B,Han S S,Wang P,et al. The bacterial communities associated with fecal types and body weight of Rex rabbits[J]. Scientific Reports,2015,5:9342.
[6]Yu X,Wu X,Qiu L,et al. Analysis of the intestinal microbial community structure of healthy and long-living elderly residents in Gaotian Village of Liuyang City[J]. Applied Microbiology and Biotechnology,2015,99(21):9085-9095.
[7]Jia H R,Geng L L,Li Y H,et al. The effects of Bt Cry1Ie toxin on bacterial diversity in the midgut of Apis mellifera ligustica(Hymenoptera:Apidae)[J]. Scientific Reports,2016,6(1):24664.
[8]林奕岑,徐 帅,倪学勤,等. 利用Illumina MiSeq测序平台分析肉鸡盲肠微生物多样性[J]. 中国农业大学学报,2016,21(12):65-73.
[9]徐 帅,林奕岑,周梦佳,等. 基于高通量测定肉鸡回肠微生物多样性及PICRUSt基因预测分析[J]. 动物营养学报,2016,28(8):2581-2588.
[10]Yu S,Wang G,Liao J,et al. Transcriptome profile analysis of mechanisms of black and white plumage determination in Black-Bone chicken[J]. Cellular Physiology and Biochemistry,2018,46(6):2373-2384.
[11]Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads. Embnet Journal,2011,17(1):10-12.
[12]Edgar R C,Haas B J,Clemente J C,et al. UCHIME improves sensitivity and speed of chimera detection[J]. Bioinformatics,2011,27(16):2194-2200.
[13]Wang Q,Garrity G M,Tiedje J M,et al. Naive bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy[J]. Applied and Environmental Microbiology,2007,73(16):5261-5267.
[14]Quast C,Pruesse E,Yilmaz P,et al. The SILVA ribosomal RNA gene database project:improved data processing and web-based tools[J]. Nucleic Acids Research,2013,41(1):590-596.
[15]Haraszthy V I,Zambon J J,Sreenivasan P K,et al. Identification of oral bacterial species associated with halitosis[J]. Journal of the American Dental Association,2007,138(8):1113-1120.
[16]Williams S T,Foster P G,Littlewood D. The complete mitochondrial genome of a turbinidvetigastropod from MiSeq Illumina sequencing of genomic DNA and steps towards a resolved gastropod phylogeny[J]. Gene,2014,533(1):38-47.
[17]張亚楠,魏单平,韩瑞丽,等. 高产期不同产蛋水平蛋鸡肠道微生物群落特征[J]. 中国兽医学报,2017,37(6):1179-1185.
[18]Torok V A,Hughes R J,Mikkelsen L L,et al. Identification and characterization of potential performance-related gut microbiotas in broiler chickens across various feeding trials[J]. Applied and Environmental Microbiology,2011,77(17):5868-5878.
[19]Lu J,Domingo J S. Turkey fecal microbial community structure and functional gene diversity revealed by 16S rRNA gene and metagenomicsequences[J]. Journal of Microbiology,2008,46(5):469-477.
[20]Wrzosek L,Miquel S,Noordine M L,et al. Bacteroides thetaiotaomicron and Faecalibacteriumprausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent[J]. BMC Biology,2013,11(1):61.陈逍遥,赵琼瑜,彭振辉,等. 乌鳖黑色素提取工艺及其结构性状[J]. 江苏农业科学,2020,48(24):182-188.
