食品中微生物间交互模型的研究进展
2020-02-08张文敏方太松耿方琳董庆利
张文敏,方太松,王 翔,耿方琳,刘 箐,董庆利,
(1.上海理工大学医疗器械与食品学院,上海 200093;2.陕西中医药大学公共卫生学院,陕西 咸阳 712000)
食品安全问题与人们的生活息息相关,其中由微生物导致的食品腐败和食源性疾病问题不容忽视[1]。食品预测微生物学可运用数学模型描述食品在流通过程中环境因素对微生物生长、残存和死亡动态的影响[2]。数学模型是食品预测微生物学研究的核心,基于预测模型得到的腐败菌生长情况可用于预测食品的货架期[3], 得到的食源性致病菌生长数据是微生物暴露评估不可缺少的部分[4]。数学模型的正确选择对准确预测微生物的生长动态至关重要。构建正确的微生物生长模型可以给食品安全决策者提供更为精确的微生物生长信息,对于保障食品质量和食品安全具有十分重要的意义。
自20世纪以来,预测微生物学专家们探究并引入多种数学模型到食品预测微生物学领域中,随之出现了多种模型分类方法[5-6]。另外,快速高效并且省时省力的分子生物学技术,如实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction, qPCR)、DNA杂交探针和变性梯度凝胶电泳技术,也被引进食品预测微生物学领域,用来监测微生物的生长、残存和死亡信息[7]。然而,无论是新数学模型的提出还是微生物监测方法的改进,大多数研究只局限于探究单一微生物在纯培养或消除本底菌的食品中的生长繁殖特点,这明显不符合实际食品中多种微生物共存的情况。并且传统预测模型建立的条件之一是假设微生物的生长环境(如温度、pH值、水分活度)在微生物整个生存期间保持不变,这与微生物在实际食品环境中的生长与存活情况差别较大。
自1990年以来,食品预测微生物学发展的主要方向之一是关注微生物在实际食品环境的生长动态[8-9]。实际食品基质中多种微生物共同存在,“微生物-微生物”及“微生物-环境”之间均存在着相互作用。这些相互作用包括相互抑制、互不影响和相互促进3 种关系。Ye Keping等[10]的研究发现在真空冷却包装的猪肉中,腐败菌之一的乳杆菌(Lactobacillusspp.)可抑制单增李斯特菌的生长。Ye等[11]的另一项研究发现冷却生猪肉中,背景菌的存在对单增李斯特菌的生长没有明显影响。Ginovart等[12]发现酸奶中的嗜热链球菌(Streptococcus thermophilus)可产生甲酸盐和CO2气体,这些产物可促进保加利亚乳杆菌(Lactobacillus bulgaricus)的生长;同时保加利亚乳杆菌也可产生一些蛋白酶促进嗜热链球菌的生长。因此,在构建微生物预测模型时,需要考虑微生物生长的实际环境。近年来,预测微生物学专家们已将微生物间交互模型引入到食品预测微生物学领域,包括不考虑微生物相互作用机制的描述性数学模型和考虑微生物相互作用机制的机械型交互模型。
本文首先简要介绍了传统的预测微生物数学模型,然后阐述了实际食品系统中微生物与环境之间的交互作用现象,介绍了描述性和机械性两类微生物间交互模型,并且分析了这两类模型的推导过程,最后对食品中微生物间交互模型的应用前景进行了展望,以期为预测微生物学的相关研究提供参考。
1 传统的预测微生物数学模型
1.1 传统的数学模型分类
1922年,Esty等[13]首先应用数学模型构建了肉毒梭菌(Clostridium botulinum)在加热作用下的失活模型。1922年—1983年是预测微生物学处于缓慢平稳的发展时期。1983年,Roberts等[14]正式提出将“食品预测微生物学”作为一门学科。同时,得益于计算机技术的发展,预测微生物学专家们将多种数学模型引入到食品预测微生物学领域,并按不同的方法进行分类[15]。
预测微生物模型分类方式有以下几种:生长模型和失活模型、概率模型和确定性模型、经验模型和机理模型。目前被广为接受的是Buchanan等[16]基于变量的不同将预测微生物模型分为一级模型、二级模型和三级模型的方法。一级模型描述微生物数量变化(生长或死亡)和时间的关系;二级模型描述一级模型中的微生物动力学参数和环境参数(如温度、水分活度、添加剂、pH值及气体包装条件)的关系;三级模型是在一级模型和二级模型的基础上通过计算机编程得到的软件,它使得非专业人士可以快速准确地获得预测微生物学相关信息。目前,传统预测微生物学的模型构建方法以及食品预测微生物学软件都已经比较完善(表1)。

表 1 主要的预测微生物生长模型及软件平台Table 1 Major PFM growth models and software platforms
1.2 传统的数学模型建模方法
预测微生物学成立的前提是微生物种群对环境条件的反应是可重现的。预测微生物学的核心在于建立正确的数学模型,然后利用已建立的数学模型预测在特定环境下微生物未来的生长和死亡动态。一级模型种类很多(表1),表达式也较复杂,但模型的实质是一元函数Nt=f(t,μ,λ,a,b,c…),其中因变量Nt是菌落浓度的对数(lg(CFU/g)或者ln(CFU/g),下文中均用lg(CFU/g)),自变量t是时间/h。参数μ是微生物生长的比生长速率/(lg(CFU/g))-h、λ是迟滞时间/h,a、b和c等字母代表其他参数。二级模型是构建一级模型中的微生物生长动力学参数和环境参数的函数关系。不同温度(T)条件下,传统的预测微生物建模过程见图1。
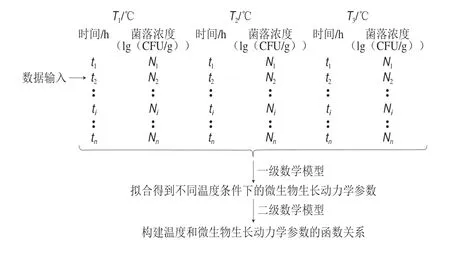
图 1 不同温度条件下单一纯菌生长模型的构建Fig. 1 Development of pure bacterial growth models at different temperatures
2 食品中微生物间交互模型
2.1 食品中微生物间交互作用
实际食品环境是一个复杂的系统,多种微生物共存于同一个食品基质。食品中“微生物-微生物”之间相互影响的关系见图2。微生物生长的“环境”包括食品基质的物化环境和食品中微生物之间的相互作用。在食品基质中,食品的环境会影响微生物的生长繁殖。同样地,微生物生长繁殖消耗掉大部分营养物质以及产生的代谢产物可以改变食品环境。另外,微生物与微生物之间也存在着协同或者拮抗作用。假如细菌N1以μ1的速率生长繁殖,细菌N2以μ2的速率生长繁殖(图2),如果其中一种细菌产生了促进细菌生长的物质,使得细菌N1和N2形成互相促进生长的协同作用,那么μ1和μ2的值就会增加;如果其中一种细菌产生了抑制细菌生长的物质,那么μ1和μ2的值就会减小。食品环境和其他微生物的存在对所要研究的微生物生长都可能产生影响;因此,构建微生物在食品中的生长模型时,需要考虑微生物实际生存的环境。
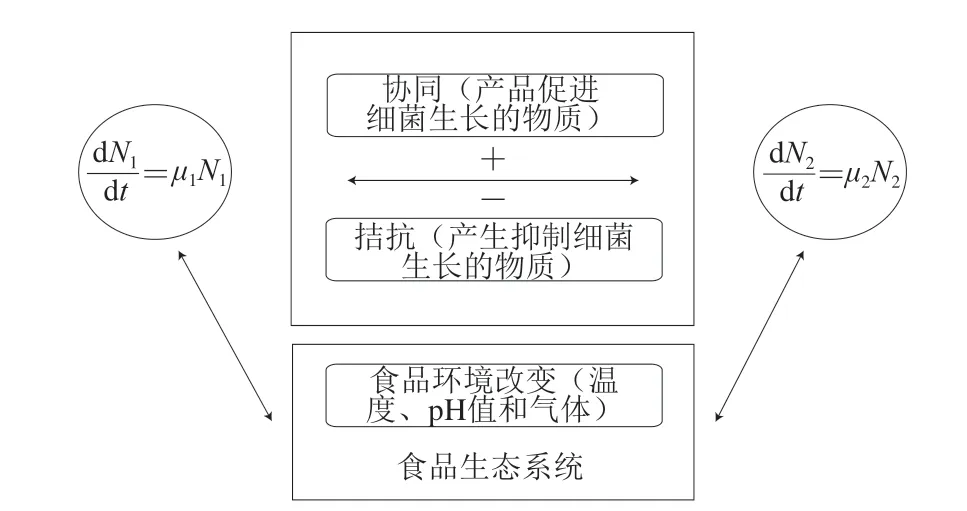
图 2 食品生态系统中微生物间交互作用图解[24]Fig. 2 Schematic representation of interactions between microorganisms in a food ecosystem[24]
Jameson[25]于1962年首先发现微生物之间存在交互作用。20世纪末,很多学者发现处于同一环境中的微生物,若其中一种优势微生物达到最大菌落浓度,另一种微生物的生长会受到影响,可能失活、停止生长或者以减小的速度继续生长。2000年,Ross等[26]正式将这种现象称作Jameson-effect理论。之后,很多学者开始关注微生物之间的交互效应,例如烟熏鱼中单增李斯特菌和腐败菌的交互作用[27]、牛奶中发酵剂对金黄色酿脓葡萄球菌(Staphylococcus aureus)的抑制作用[28]及猪肉中单增李斯特菌和假单胞菌[29]的竞争生长作用。
另外,处于食物链中同一位置的动物之间存在着竞争食物的关系,这种现象可用Lotkae-Volterra效应描述。Dens等[30]于2003年将Lotkae-Volterra模型引入到预测微生物学领域描述微生物之间的交互作用。Lotkae-Volterra模型建立的理论基础是:消耗同一种营养物质的两种微生物之间存在着交互作用,当营养物质耗尽,两种微生物同时停止生长。Vimont等[31]用Lotkae-Volterra模型构建了大肠杆菌和背景菌在牛肉中的生长模型。Giuffrida等[32]研究了单增李斯特菌在意大利香肠中的生长情况。Mounier等[33]研究了奶酪中“酵母菌-酵母菌”及“酵母菌-微生物”之间的交互作用。
2.2 食品中微生物间交互模型
微生物之间的交互作用有3 种:相互促进、互不影响或者相互抑制作用。目前发现最多的是食品基质中存在着优势菌群抑制弱势菌群的现象[34-37]。例如在发酵香肠中乳酸菌快速生长繁殖,抑制了大肠杆菌和单增李斯特菌的生长[38]。乳酸菌对其他微生物的抑制作用可能是由于抑菌代谢产物如乳酸、细菌素和CO2等的产生,还可能是微生物之间的营养竞争和细胞接触[39-41]作用。预测微生物学家已经注意到微生物间的相互作用会改变微生物的生长形式,用传统的预测微生物模型描述微生物的生长已经不太合适。近些年来,两菌交互模型如描述性模型和机械性模型已经被引入到预测微生物学领域(表2)。
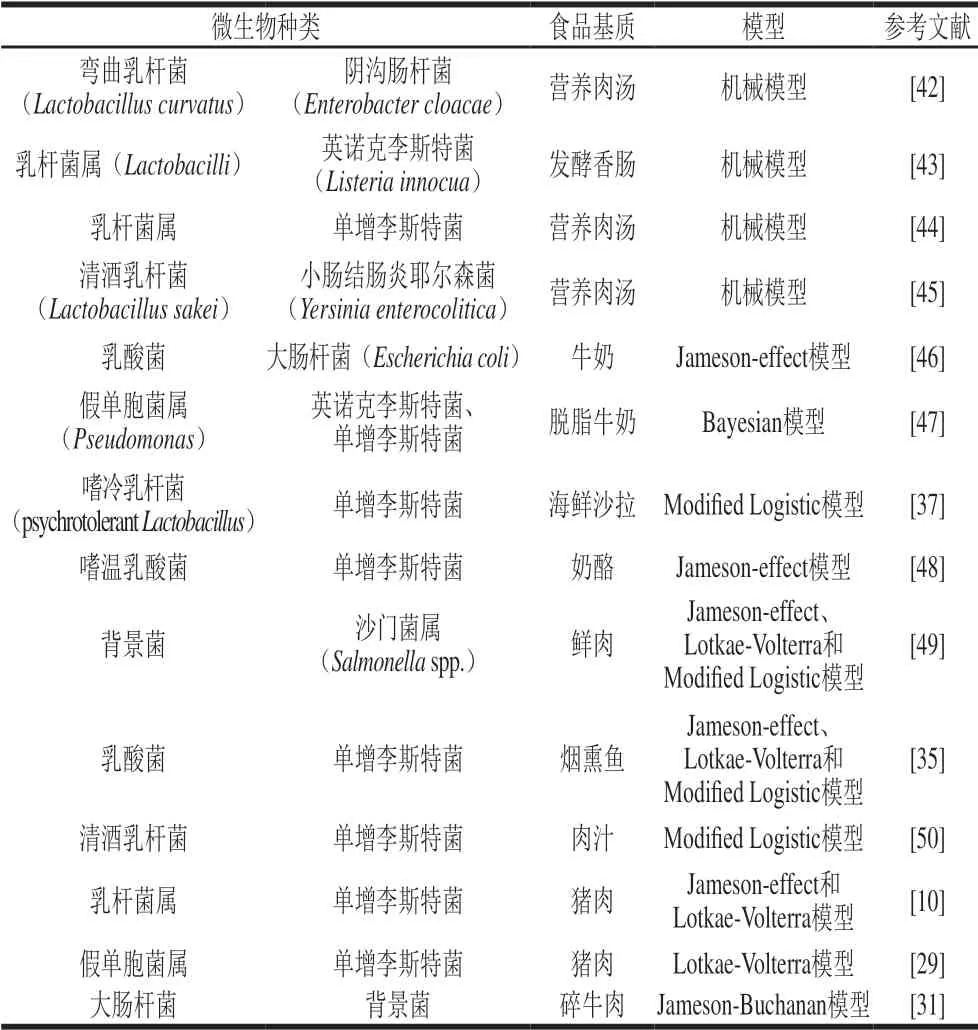
表 2 应用交互模型描述微生物生长动态Table 2 Application of interaction models to describe microbial growth
2.2.1 描述性微生物间交互模型
不同于传统的预测微生物模型,在交互模型中,每一个时间点(ti)对应着两种微生物的菌落浓度(N1i和N2i),微生物间交互模型的建立过程如图3所示。描述性交互模型不考虑微生物间交互作用的机制,直接将微生物菌落浓度作为一个抑制条件,用简单的数学模型量化其他微生物对目标微生物生长的影响。描述性交互模型主要包括Jameson-effect模型和Lotkae-Volterra模型,这两种模型是基于传统的Baranyi模型和Buchanan三阶段线性模型扩展而来的。不同温度条件下,微生物间建模过程如图3所示。
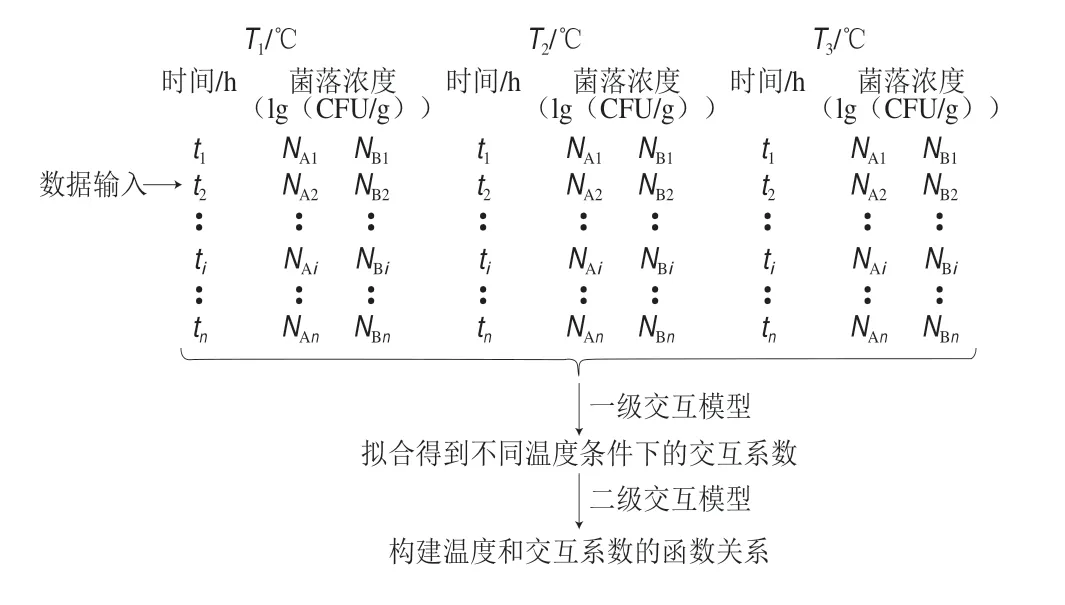
图 3 不同温度条件下微生物间交互模型的构建Fig. 3 Development of microbial interaction models at different temperatures
为了进一步了解传统一级模型和一级交互模型之间的关系,现将Jameson-Baranyi模型、Jameson-Buchanan模型和Lotkae-Volterra模型公式进行推导,具体如下。
微生物生长模型的一般形式见公式(1)[8]。
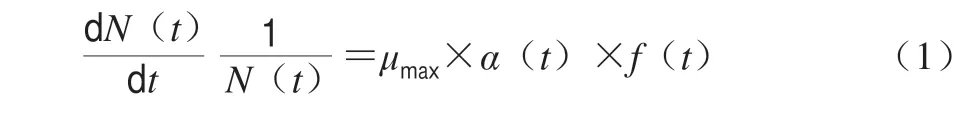
式中:α(t)是调整函数;f(t)是抑制函数;μmax是微生物的最大比生长速率。
根据微生物自身生理特性不同,两菌有不同的调整函数;因为两菌在一个食品基质中,它们具有同一个抑制函数,所以描述性交互模型的一般形式如式(2)所示。
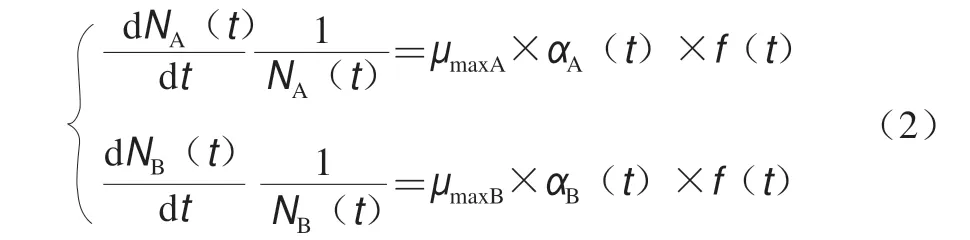
在Baranyi模型中,适应函数和抑制函数分别见 式(3)、(4)。
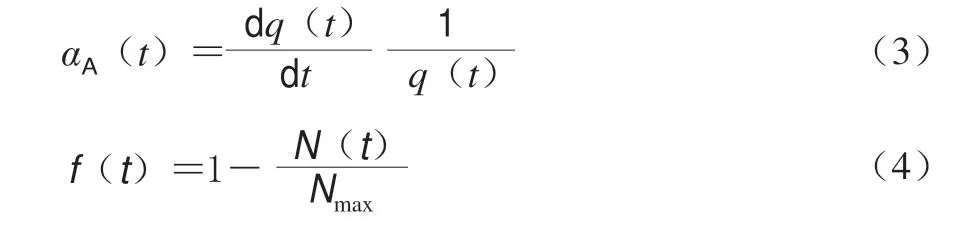
式中:q是微生物的生理状态λ是迟滞期。
在Buchanan三阶段模型中[16],适应函数和抑制函数分别见式(5)、(6)。

式中:tmax是微生物达到最大菌落浓度所需的时间。
在Lotkae-Volterra模型中,适应函数αA(t)同 式(3),抑制函数[51]见式(7)。
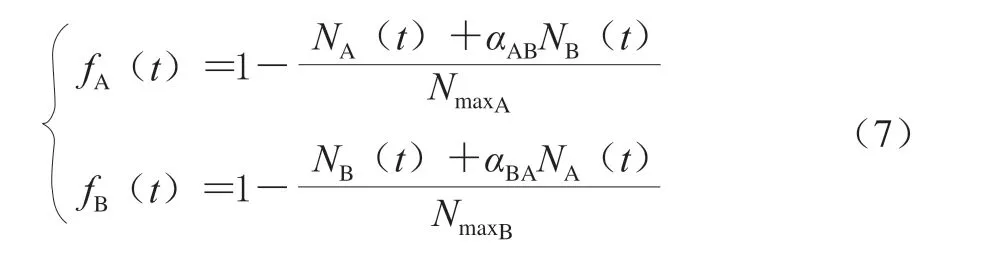
因此Jameson-Baranyi模型见式(8)。
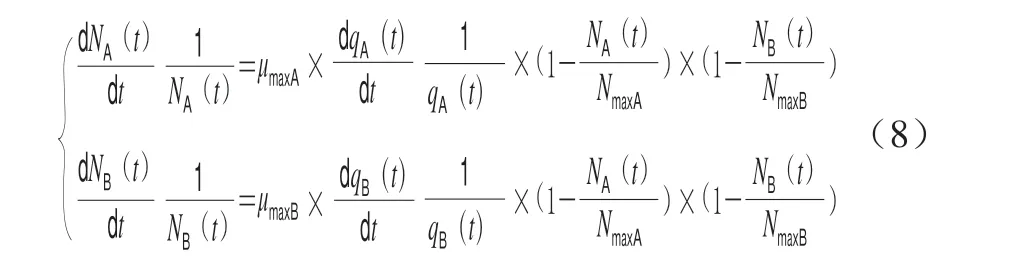
Jameson-Buchanan模型见式(9),其中tmax是优势微生物达到最大菌落浓度的时间。
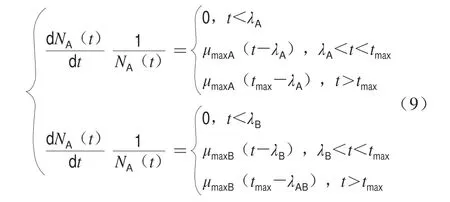
Lotkae-Volterra模型见式(10)。
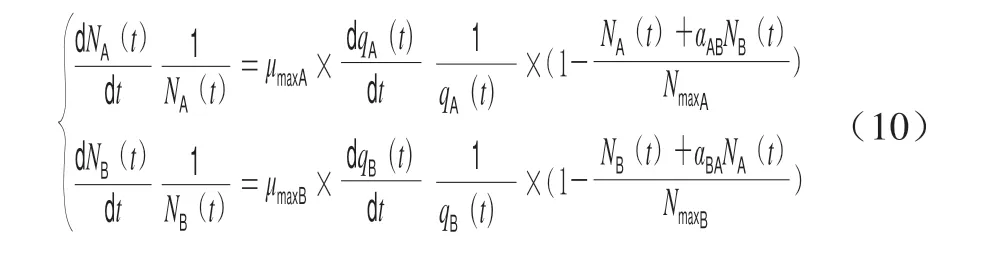
式中:αAB和αBA反映了两个微生物种群的交互情况。当αAB(αBA)>1时,说明微生物B(A)达到最大菌落浓度后,微生物A(B)浓度减少;当αAB(αBA)=1时,说明微生物B(A)和A(B)无相互作用。当αAB(αBA)<1 时,说明微生物B(A)达到最大菌落浓度后,微生 物A(B)浓度增加。
Jameson-effect模型的成立是基于“同一系统中两种细菌生长过程中的减速是同时进行”这一假设的,这个假设非常简单,因此,Jameson-effect模型的形式也不复杂[10]。但是这一假设并不一定适用于所有微生物之间的交互情况,因此在拟合过程中效果并不是很好。如Ye Keping等[10]在研究猪肉中乳杆菌对单增李斯特菌的抑制作用时发现,乳杆菌Nmax的预测值比实测值低。可根据细菌的实际生长情况对Jameson-effect模型进行改良,以提高其拟合优度。Le Marc等[52]在研究牛奶中乳酸菌对金黄色葡萄球菌的抑制作用时发现,乳酸菌达到最大菌落浓度前,金黄色葡萄球菌停止了生长,因此在模型中增加了一个“关键细胞浓度”的参数(小于最大细胞浓度);且发现修正后的模型更适用于描述乳酸菌对金黄色葡萄球菌的抑制作用,修正后的模型说明金黄色葡萄球菌比乳酸菌对营养限制或者代谢产物更加敏感。Mejlholm等[37]在研究蛋黄酱海鲜沙拉中乳杆菌对单增李斯特菌的抑制作用时,在抑制函数中加了一个抑制系数“γ”,若γ<1,说明在乳杆菌停止生长时,单增李斯特菌浓度继续增加;若γ>1,表示单增李斯特菌浓度减小;若γ=1,表示单增李斯特菌停止生长。这种改良形式的Jameson-effect模型能够很好地描述两种微生物之间的交互作用。
Lotkae-Volterra模型含有两个参数αAB和αBA,因此解微分方程的过程比较复杂。并且有研究表明Lotkae-Volterra模型能够高估细菌的迟滞期时间,低估生长速率,这种预测结果是“失败-危险”的[10],因此Lotkae-Volterra模型不适宜直接作为一级交互模型来模拟两种细菌的交互作用。但是Lotkae-Volterra模型中参数αAB和αBA反映了两个微生物种群的交互情况,可以用来评估处于同一生态系统中的两种细菌是否具有相互作用,进而再进行后续研究。还有研究将Lotkae-Volterra模型改良后作为三级模型,用来描述环境因子对细菌竞争作用程度的影响[53]。
2.2.2 机械性模型
机械性模型将微生物整个生长过程中环境条件的动态变化及微生物之间的交互机制考虑进去,比如微生物生长过程中食品环境p H 值的变化和营养物质的消耗等。微生物生长模型的一般形式为:Bernaerts等[24]认为,在整个食品基质中,n种微生物共同生长时符合公 式(11)所示的规律。
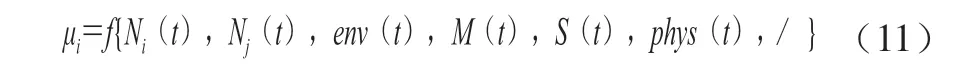
由式(11)可知,μi的大小受到环境中微生物菌种(i,j,…,i≠j)、环境因素(env)、代谢物(M)、营养物质(S)及物理状态(phys)等各种因素影响。根据具体环境分析微生物的生长受到哪些因素影响,当μmax>0时,说明微生物在生长,当μmax<0时,说明微生物处于失活情况。
在机械性模型中,由某一种环境参数“x”引起微生物“i”的变化可用式(12)[18]表示。

式中:μiopt(x)是不存在抑制因子时,微生物的最优比生长速率。当γ=1时,说明微生物的生长不受抑制;当γ=0时,说明微生物的生长完全受到抑制。另外,环境“x”一般随时间一直变化。
以环境pH值的变化以及微生物营养竞争为例介绍机械性模型的使用方法。Martens等[42]在研究弯曲乳杆菌对阴沟肠杆菌的影响时,考虑到了弯曲乳杆菌会产生酸性代谢产物,进而影响环境的pH值,他在描述环境中pH值的变化时用到了式(13)。
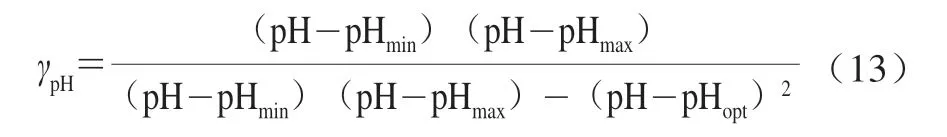
式中:pHopt、pHmin和pHmax分别是适合微生物生长的最优pH值、最小pH值和最大pH值。
Martens等[42]还考虑到了微生物之间的营养竞争作用,他认为营养消耗的规律符合莫纳德生长动力学 (式(14))。

式中:S是营养物质水平;KS是莫纳德常数。
当然,还可能是其他环境参数的变化影响了微生物的生长。根据“γ概念”,如果微生物的生长受到多种环境因素的抑制,可将γ函数相乘(如式(13)和 式(14))[54-55]。Leroy等[43]根据“γ概念”构建了清酒乳杆菌CTC 494菌株在生长过程中通过消耗葡萄糖、产生乳酸及产生细菌素来抑制单增李斯特菌的模型。Martens等[42]计算出了弯曲乳杆菌和阴沟肠杆菌(Enterobacter cloacae)的交互作用开始时间。另外,Couvert等[56]研究了CO2、温度及pH值对单增李斯特菌和荧光假单胞菌生长能力的影响,发现CO2气体能够溶解于水中改变“环境”,进而影响致病菌的生长。Poschet等[57]也用这种办法构建了乳酸乳球菌(Lactococcus lactis)和英诺克李斯特菌(Listeria innocua)的共同生长模型。机械性交互模型可将影响微生物生长的每一个因素都考虑进去,能够很好地描述微生物间交互影响的过程,但是模型建立的过程比较复杂,需要极强的数学与计算机能力。
3 结 语
食品预测微生物学的主要目的是运用数学模型对微生物的生长进行定量分析,可帮助人们在没有进行微生物检测的前提下,预测微生物的生长和死亡,为食品安全提供重要保障。食品预测微生物学在食品货架期、食品安全的预测和管理中有很大的应用价值。构建正确的数学模型可为食品安全决策者提供准确的微生物信息。微生物在食品中绝不是单独存在的,预测微生物学未来的发展方向必将是模拟微生物在实际食品中的生长情况。近些年,预测微生物学家已将微生物间交互模型引入到食品预测微生物学中。本文介绍了微生物间交互模型的理论基础,并且分析了微生物间交互模型的推导过程,为今后微生物间交互模型的相关研究提供参考。尽管已有较多文献研究微生物间交互模型,但要使得微生物间交互模型服务于食品安全控制还有很多工作需要完善,对微生物间交互模型未来的研究方向展望如下: 1)目前微生物间交互模型还多集中于两种微生物之间的相互作用,但是实际食品中肯定是多种微生物共存的,研究多种微生物之间的相互作用更具有实际意义;2)已有的食品货架期预测模型和微生物暴露评估研究中用到的数学模型大多还是单一菌种在纯培养状态下的数学模型,要准确预测微生物的生长情况,还需将两种甚至多种细菌之间的交互模型实际应用到食品质量和食品安全控制中;3)当多种微生物共存时,传统的平板计数方法可能无法将微生物分离开来,因此需要将分子生物学方法如qPCR技术和DNA杂交探针技术等引入到预测微生物学中;4)实际食品中微生物的菌落浓度较低,因此构建单细胞水平下的微生物间交互模型更具有实际意义;5)微生物间交互模型的数学形式比较复杂,模型建立的过程也比较繁琐,因此可将微生物间交互模型结合到已有的微生物预测软件中,使得非专业人士也可以快速准确地获得预测微生物学相关信息的指导,为数据分析奠定工作基础。
