鲈鲤仔鱼饥饿试验及不可逆生长点的确定
2020-01-21张建明姚金忠姜伟郭文涛
张建明 姚金忠 姜伟 郭文涛
摘要:【目的】全面掌握鱸鲤仔鱼早期培育过程的摄食特点和饥饿耐受力,为其苗种规模化培育提供技术支撑。【方法】在18.5~21.9 ℃水温条件下,利用人工繁殖孵化获得的鲈鲤初孵仔鱼进行饥饿试验,研究饥饿胁迫对鲈鲤仔鱼生长、发育、存活、卵黄囊吸收、摄食率及摄食强度的影响,并确定鲈鲤仔鱼的不可逆生长点(PNR)。【结果】饥饿对鲈鲤仔鱼全长、体长、体高及肛前长等生长指标有极显著影响(P<0.01,下同),对眼径和鳔等组织器官发育也有明显影响。鲈鲤仔鱼5日龄开口摄食,进入混合营养期,10日龄时仔鱼卵黄囊基本消耗殆尽,鲈鲤仔鱼混合营养期维持5 d。鲈鲤仔鱼5日龄开始摄食外源性营养,初次摄食率约20%,随后逐步提高,至13日龄时达最高值(100%),之后开始下降,在17日龄时下降至40%,低于最大初次摄食率的50%。根据PNR定义,即17日龄是鲈鲤仔鱼的PNR。在5~11日龄期间(饥饿7 d内),鲈鲤仔鱼的初次摄食强度和摄食能力随着饥饿天数的增加呈上升趋势,12日龄后(饥饿7 d以上)则随着饥饿天数的增加呈迅速下降趋势。饥饿组仔鱼的累计死亡率在17日龄(饥饿13 d)时剧增,到18日龄(饥饿14 d)时其累计死亡率达88.00%,而摄食组仔鱼的累计死亡率仅为14.75%;至19日龄(饥饿15 d)时饥饿组仔鱼的累计死亡率高达100.00%,摄食组仔鱼的累计死亡率仍为14.75%。饥饿组鲈鲤仔鱼的50.00%累计死亡率出现日龄为18日龄,即在PNR出现之后。【结论】在18.5~21.9 ℃水温条件下,鲈鲤仔鱼混合营养期为5 d,PNR发生在17日龄。鲈鲤仔鱼可在5日龄后开始投喂,最佳投喂时间为8日龄,但最晚投喂时间不宜超过13日龄。
关键词: 鲈鲤;仔鱼;饥饿;摄食率;摄食强度;不可逆生长点(PNR)
中图文分类号: S965.199 文献标志码: A 文章编号:2095-1191(2020)10-2565-08
Experimental starvation on Percocypris pingi larvae and determination of the point of no return
ZHANG Jian-ming1,2, YAO Jin-zhong3*, JIANG Wei1,2, GUO Wen-tao1,2
(1Institute of Chinese Sturgeon Research, China Three Gorges Project Corporation, Yichang, Hubei 443100, China; 2Hubei Key Laboratory of Three Gorges Project for Conservation of Fishes,Yichang, Hubei 443100, China;
3China Three Gorges Corporation Operation and Administration Center for River Basin Hydro Complex,
Yichang, Hubei 443100, China)
Abstract:【Objective】To study the feeding characteristicsandstarvation tolerance of Percocypris pingi during the early development of larvae, and provide technical support for the large-scale breeding of larvae. 【Method】Under the condition of water temperature of 18.5-21.9 ℃, the experimental ecology method was used to conduct starvation experiments on the newly hatched larvae obtained by artificial breeding and hatching. Analyze and study the effects of starvation stress on the growth, development, survival, yolk sac absorption, feeding rate and feeding intensity of larvae, and initially determine the point of no return(PNR) of larvae. 【Result】The growth index of the full length, body length, body height and anal length of the larvae of P. pingi had extremely significant effect(P<0.01, the same below), and had obvious effects on the development of tissues and organs such as eye diameter and sputum. The larvae of P. pingi were fed at 5 d of age and entered the mixed nutrition period. At 10 d of age, the yolk sac of the larvae was basically consumed, and the mixed nutrition period for larvae of P. pingi were 5 d. The larvae of P. pingi began to eat exogenous nutrition at the age of 5 d. The initial feeding rate was about 20%, and then the initial feeding rate gradually increased. The initial feeding rate of the 13-day-old larvae reached the highest value of 100%. Then, the initial feeding rate began to decline. By the 17 d of age, the initial feeding rate dropped to 40%, which was lower than 50% of the maximum initial feeding rate. According to the definition of PNR , the 17 d of age was PNR for larvae of P. pingi. The larvae of P. pingi initial feeding intensity and feeding ability increased with the increase of starvation days during 5-11 d of age(starved within 7 d). After 12 d of age(starved over 7 d), the initial feeding intensity and feeding ability of larvae decreased rapidly with the increase of hunger days. The cumulative mortality rate of starved group increased sharply on 17 days old(starved for 13 d). When the starved group was 18 days old(starved for 14 d), the cumulative mortality rate reached 88.00%, and the cumulative mortality rate of the larvae in the feeding group was 14.75%. At 19 d of age, the cumulative mortality rate of the hunger fish in the starved group was 100.00%, while the cumulative mortality rate of the larvae in the feeding group was 14.75%. 50.00% cumulative mortality of larvae in the hunger-stricken fish in starved group was 18 days old, after PNR. 【Conclusion】Under the condition of water temperature of 18.5-21.9 ℃, the mixed nutrition period for larvae of P. pingi were 5 d, and PNR occurs at 17 d of age. Larvae of P. pingi can be fed after 5 d of age. The best feeding time is 8 d of age, but the latest feeding time should not exceed 13 d of age.
Key words: Percocypris pingi; larvae; starvation; feeding rate; feeding intensity; point of no return(PNR)
Foundation item:National Key Research and Development Program of China(2016YFC0502210); National Natural Science Foundation of China(51779016); Science and Technology Planning Project of China Three Gorges Project Corporation(ZHX2017025,JG/18001B)
0 引言
【研究意义】鲈鲤(Percocypris pingi)又称青脖、金甲鱼或花鱼,隶属于鲤科(Cyprinidae)鲃亚科(Barbinae)鲈鲤属(Percocypris),在我国长江上游的干支流、西江流域、南盘江等水系均有分布(詹会祥等,2016)。由于过度捕捞及环境污染等因素的影响,鲈鲤资源逐年衰减,在长江鱼类受威胁程度定量分析中已达到长江一级急切保护程度(刘军,2004),為四川省珍稀保护鱼类。在鱼类生活史中,仔鱼期是鱼类对环境条件最敏感的生活阶段,极易受各种外界因素的影响而发生死亡,其中饥饿是引起仔鱼死亡最主要的原因之一(叶忠平等,2012;段国庆等,2013)。初次摄食期遭受饥饿胁迫引起死亡是自然状况下仔鱼数量变动的主要原因,而不可逆生长点(PNR)是衡量仔鱼耐饥饿能力的常用评判标准(夏连军等,2004)。孵化出膜后的仔鱼需经历从内源性营养过渡到外源性营养,在此期间若不能建立主动摄食机制度过耐受期,将无法恢复摄食能力,到达PNR而最终死亡(杨育凯等,2019)。PNR是从生态学角度测定仔鱼耐饥饿能力的一项关键指标,是初次摄食仔鱼耐受饥饿的时间临界点。因此,通过饥饿试验确定仔鱼的饥饿耐受力及其PNR,对研究鱼类早期生活史及指导苗种培育均具有重要意义。【前人研究进展】至今,有关鱼类仔鱼饥饿耐受力及其PNR的研究已有较多报道。夏连军等(2004)研究发现,在水温(22.0±1.0)℃、盐度31.0‰的条件下,黄鲷(Dentex tumifrons)仔鱼孵化后第4 d开始摄食外部食物,幼体发育进入混合营养期,混合营养期仅维持3 d;黄鲷仔鱼最高初次摄食率出现在孵出后第7 d,PNR出现第8~9 d。李强等(2009)研究指出,在(20.0±1.5)℃水温条件下白甲鱼(Onychostoma sima)仔鱼到达PNR的时间为13.5 d,且PNR出现时间与50%累计死亡率出现时间基本相同,说明第13.5 d是白甲鱼仔鱼早期发育的敏感阶段。张永泉等(2009)研究表明,在10.0~11.0 ℃水温条件下哲罗鱼(Hucho taimen)仔鱼的初次摄食时间在21日龄,至29日龄时摄食率达100%,PNR出现在39~40日龄,因此初次摄食仔鱼最佳投喂时间在25日龄,即仔鱼上浮后第4 d。马建波等(2012)通过开展杂交鳢仔鱼饥饿试验得知,在(26.0±1.0)℃水温条件下杂交鳢仔鱼孵化后第2 d开始摄食,进入混合营养期,混合营养期维持5 d,PNR出现在仔鱼孵出后的第7~8 d。段国庆等(2013)研究表明,黄鳝(Monopterus albus)仔鱼在25~28 ℃水温条件下于出膜后第6 d开始向外界摄食,8日龄时卵黄囊吸收殆尽,5~8日龄为混合营养阶段,16日龄抵达PNR,即黄鳝仔鱼的最佳投喂时间在8日龄。贾瑞锦等(2014)研究表明,在水温(22.9±0.8)℃、盐度30‰~32‰的条件下,条纹锯鮨(Centropristis stria-ta)仔鱼在孵化后2 d开口摄食,从内源营养期转入混合营养期(持续1 d),3日龄时进入外源营养期,其PNR出现在5日龄,即条纹锯仔鱼孵出后2~5 d为摄食的关键期。李艳秋等(2014)研究显示,在10.0~11.0 ℃水温条件下,太平洋鳕(Gadus macrocephalus)仔鱼在孵化出膜后第5 d开始开口摄食,卵黄囊体积随仔鱼生长不断减小,仔鱼初次摄食率为30%,至7日龄的初次摄食率达90%,但只维持1 d,PNR出现在9日龄,因此太平洋鳕仔鱼最佳投饵时间为5~7日龄。王晓龙等(2017)研究表明,花鲈(Lateolabrax maculatus)初孵仔鱼在水温(21±1)℃、盐度26‰的条件下于孵化后4日龄开始摄食外源食物进入混合营养期,混合营养期仅维持2 d,PNR出现在9~10日龄,其最佳投喂时间在4日龄。陈淑吟等(2018)研究表明,黑鲷(Acanthopagrus schlegelii ♀)×真鲷(Pagrosomus major ♂)杂交F2代仔鱼在(21.8±0.4)℃水温条件下的混合营养期为4~5 d,6日龄时抵达PNR,饥饿仔鱼5日龄时呈负生长,最适开口投饵时期为仔鱼开口后3 d内。宋勇等(2019)研究显示,在(20.0±1.0)℃水温条件下,叶尔羌高原鳅(Triplophysa yarkandensis )仔鱼的混合营养期为3~4日龄,7日龄时的初次摄食率最高(90%),PNR出现在8~9日龄,其最佳投喂时间为仔鱼开口后4 d内。杨育凯等(2019)研究表明,在水温18~19 ℃条件下黑棘鲷(Acanthopagrus schlegelii)仔鱼在3日龄开口摄食,至4日龄时卵黄囊消耗殆尽,6日龄时油球消失,混合营养期仅维持3 d,属于易遭受饥饿胁迫的鱼类品种,即育苗中的最适投饵时间在仔鱼开口后4 d内。【本研究切入点】PNR是判断初孵仔鱼耐饥饿能力及确定其最适开口投饵时期的重要依据,但关于鲈鲤仔鱼饥饿试验和PNR的研究至今未见相关报道。【拟解决的关键问题】以鲈鲤初孵仔鱼为研究对象,对饥饿过程中仔鱼卵黄消耗、初次摄食率、初次摄食强度、生长状况及部分组织器官发育变化等进行分析并确定其PNR,旨在掌握鲈鲤仔鱼早期培育过程的摄食特点和饥饿耐受力,为其苗种规模化培育提供技术支撑。
1 材料与方法
1. 1 试验材料
1. 1. 1 初孵仔鱼来源 2019年3—4月在中国长江三峡集团有限公司中华鲟研究所乌东德实验站进行试验。对人工驯养鲈鲤亲鱼进行人工催产,通过挤压腹部获得成熟卵子和精子,干法授精获得受精卵,流水孵化获得初孵仔鱼。
1. 1. 2 仔鱼培育 初孵仔鱼置于静水微充气的玻璃纤维缸(2.0 m×0.8 m×0.6 m)中培育,试验用水为无天然生物饵料且经沉淀静置、充分过滤后的金沙江江水;每日换水1次,换水量为1/2。试验期间培育水温遵循自然水温节律,维持在18.5~21.9 ℃,溶解氧>6.0 mg/L。
1. 1. 3 生物饵料 试验用生物饵料为丰年虫(Artemia salina),由市售丰年虫卵在水温25.0~28.0 ℃、盐度20‰条件下孵化获得丰年虫幼体。
1. 2 试验方法
1. 2. 1 仔鱼饥饿试验 取同一批次的孵化仔鱼置于玻璃纤维缸中进行暂养,开口前1 d随机分为3组(T1、T2和T3),每组500尾仔鱼。5日龄开始投饵,其中,T1组每日投喂3次丰年虫,作为正常摄食组;T2和T3组不投喂饵料,作为饥饿组。每日观察记录各组仔鱼行为活动及死亡情况,并分别从T1和T2组随机抽取10尾仔鱼,测量记录其全长、体长、体高、肛前长、眼径、鳔长、鳔高、卵黄囊长和高等生物学数据。
(1)卵黄囊体积和鳔体积计算公式(李艳秋等,2014;杨育凯等,2019):
V=(4/3)·π·(r/2)2·R/2
式中,r为卵黄囊(鳔)高,R为卵黄囊(鳔)长。
(2)开鳔率计算公式:
开鳔率(%)=鳔充气仔鱼数/样本仔鱼数×100
1. 2. 2 初次摄食率和初次摄食强度 鲈鲤仔鱼开口摄食后,每日从T3组中随机抽取20尾仔鱼放入微充气且盛有足量丰年虫的1 L水体塑料盆中。摄食2 h后取出仔鱼,在解剖镜下解剖观察其摄食情况,并对每尾仔鱼摄食的丰年虫进行计数,摄入的丰年虫卵当作丰年虫计数,计算仔鱼初次摄食率和摄食强度。
初次摄食率(%)=摄食仔鱼数/样本仔鱼数×100
初次摄食强度(个/尾)=初次摄食仔鱼消化道内
丰年虫数/样本仔鱼数
1. 2. 3 PNR确定 以鲈鲤仔鱼孵化后的日龄表示,每日测定饥饿试验组仔鱼的初次摄食率,当仔鱼初次摄食率低于最高初次摄食率的50%时,即为PNR的出现时间。
1. 3 统计分析
采用Excel 2013和SPSS 19.0對试验数据进行统计分析及制图,并以t 检验比较分析饥饿组和摄食组仔鱼的各项生物学指标。
2 结果与分析
2. 1 饥饿组和摄食组鲈鲤仔鱼的生长变化情况
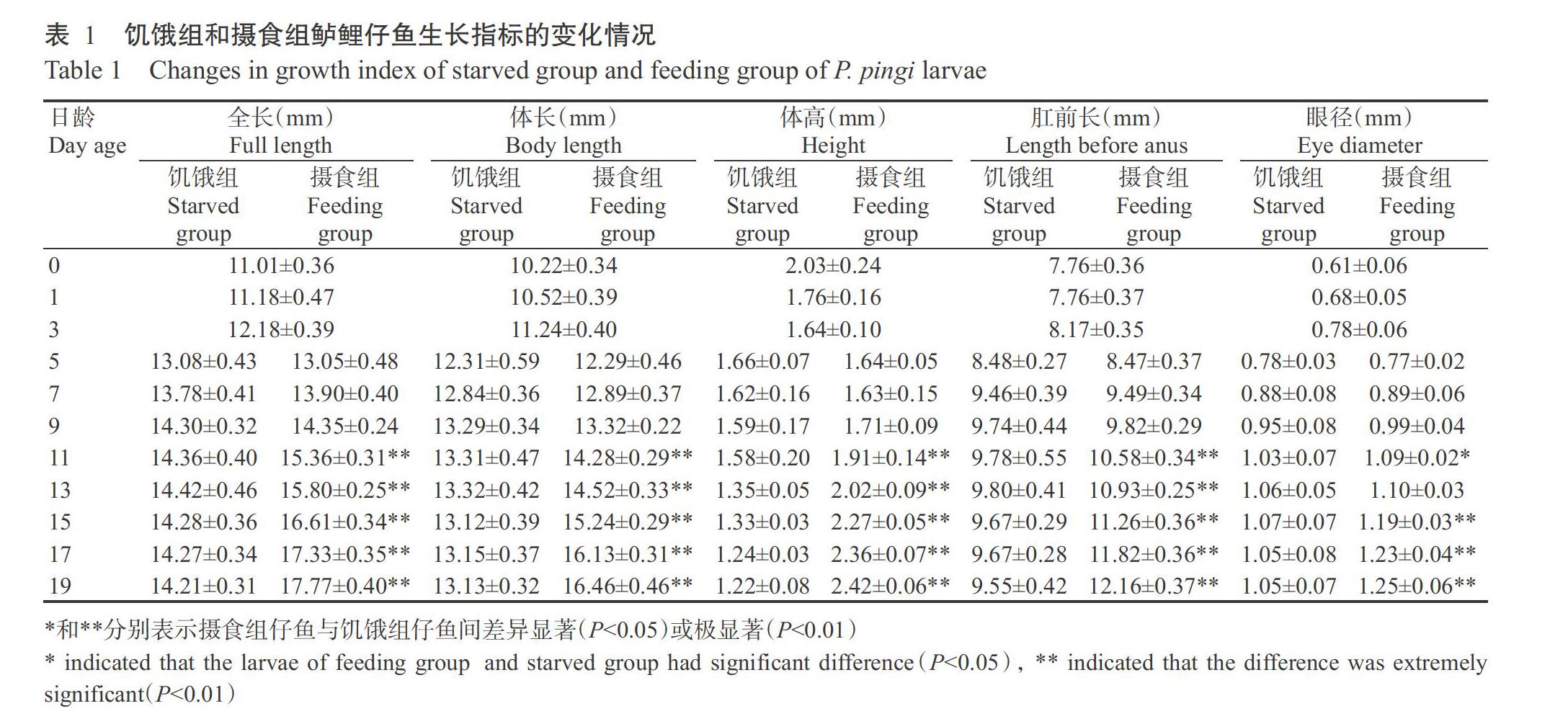
在18.5~21.9 ℃水温条件下,鲈鲤初孵仔鱼全长11.01±0.36 mm、体长10.22±0.34 mm、体高2.03±0.24 mm、肛前长7.76±0.36 mm、眼径0.61±0.06 mm(表1)。随着日龄的增加,仔鱼依靠卵黄等内源性营养不断生长,至3日龄时,仔鱼全长12.18±0.39 mm、体长11.24±0.40 mm、体高1.64±0.10 mm、肛前长8.17±0.35 mm、眼径0.78±0.06 mm。本研究对4日龄仔鱼进行分组,5日龄时摄食组开始投喂丰年虫饵料。9日龄的生长数据指标显示,摄食组仔鱼的全长、体长等生长指标均高于饥饿组仔鱼,但差异不显著(P>0.05,下同);11日龄后两组仔鱼的全长、体长等生长指标变化差异极显著(P<0.01,下同),眼径差异显著(P<0.05)。摄食组仔鱼由内源性营养完全过渡到外源性营养后,其生长速度加快,仔鱼整体较粗壮,至19日龄即试验结束时,仔鱼全长17.77±0.40 mm、体长16.46±0.46 mm、体高2.42±0.06 mm、肛前长12.16±0.37 mm、眼径1.25±0.06 mm。而饥饿组仔鱼在10日龄后内源性营养基本消耗殆尽,又未及时建立外源性营养,仔鱼全长、体长、肛前长、眼径等生长几乎停滞,仔鱼消瘦明显;至17日龄时多数仔鱼头部向下缓慢游动、对外界刺激反应迟钝;至19日龄时仔鱼全长14.21±0.31 mm、体长13.13±0.32 mm、体高1.22±0.08 mm、肛前长9.55±0.42 mm、眼径1.05±0.07 mm。
2. 2 饥饿组和摄食组鲈鲤仔鱼卵黄囊的吸收情况
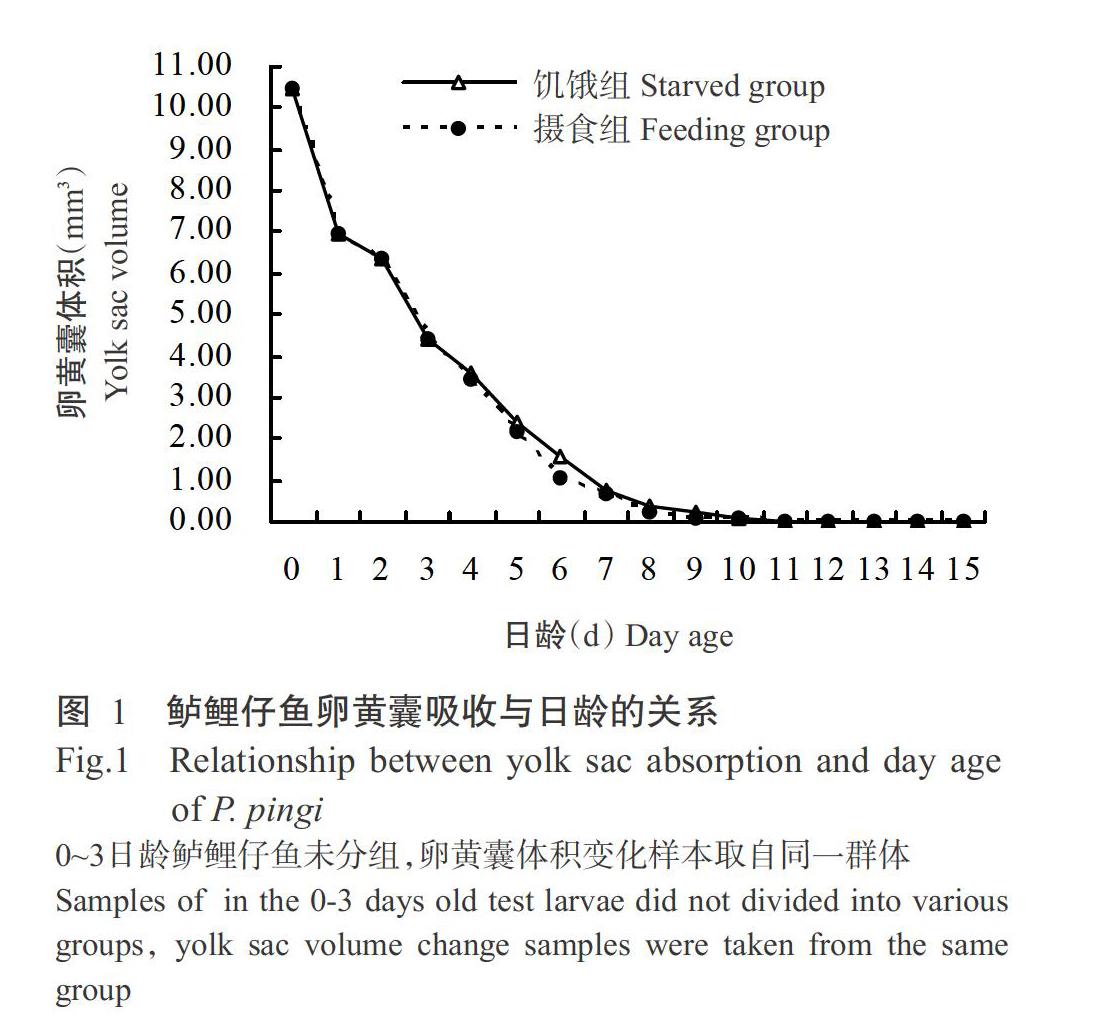
由图1可看出,鲈鲤初孵仔鱼卵黄囊体积平均为10.44±2.25 mm3。在18.5~21.9 ℃水温条件下,鲈鲤仔鱼卵黄囊体积随着日龄的增加及生长发育而迅速消耗,以0~4日龄阶段尤为明显;至4日龄时,摄食组仔鱼卵黄囊体积为3.43±0.52 mm3,仅占初孵仔鱼卵黄囊体积的32.85%,饥饿组仔鱼卵黄囊体积为3.57±0.43 mm3,仅占初孵仔鱼卵黄囊体积的34.20%。5日龄开始投饵后,两组仔鱼的卵黄囊消耗及卵黄囊体积变化略有差异,表现为摄食组仔鱼卵黄囊消耗速度略快于饥饿组仔鱼,卵黄囊体积明显小于饥饿组仔鱼;至9日龄时,饥饿组和摄食组仔鱼的卵黄囊进一步缩小,其体积分别为0.19±0.14和0.10±0.04 mm3;至10日龄时,两组仔鱼的卵黄囊基本消耗殆尽。即在18.5~21.9 ℃水温条件下,鲈鲤仔鱼混合营养期维持5 d。
2. 3 饥饿组和摄食组鲈鲤仔鱼开鳔率及鳔体积的变化情况
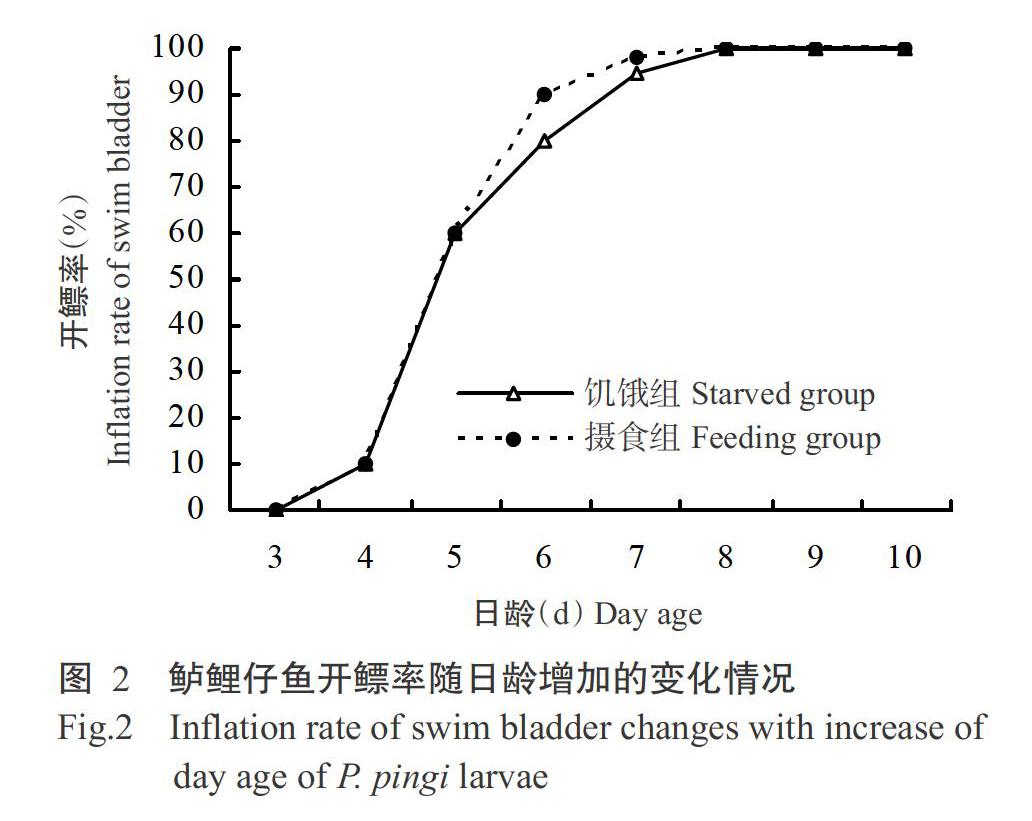
仔鱼鳔的形成及充气是其准备开口摄食的重要标志之一(李艳秋等,2014)。在18.5~21.9 ℃水温条件下,随着鲈鲤仔鱼的发育,在4日龄时即可观察到仔鱼鳔组织出现,表明仔鱼卵黄囊尚未吸收完毕时已做好开口前的准备;5日龄时仔鱼开鳔率迅速增长,至7日龄时两组仔鱼的开鳔率均超过95%(图2)。在仔鱼鳔体积(图3)方面,4~12日龄期间两组仔鱼鳔体积整体上均呈增长趋势,但鳔体积变化差异较小,饥饿组仔鱼鳔体积范围在0.01±0.01~0.41±0.10 mm3,摄食组仔鱼鳔体积范围在0.02±0.02~0.51±0.17 mm3。13日龄后,摄食组仔鱼鳔体积随着日龄的增加继续生长增大,至19日龄时其鳔体积为0.79±0.12 mm3;而饥饿组仔鱼鳔体积在13日龄后开始逐步缩小,至19日龄时其鳔体积仅为0.08±0.04 mm3。
2. 4 鲈鲤仔鱼初次摄食率、摄食强度及PNR的确定
鲈鲤仔鱼初次摄食率变化如图4所示。在18.5~21.9 ℃水温条件下,鲈鲤仔鱼在5日龄开始摄食外源性营养,初次摄食率约20%,随后逐步提高,至13日龄时达最高值(100%),之后开始下降,在17日龄时下降至40%,低于最大初次摄食率的50%。根据PNR定义,即17日龄是鲈鲤仔鱼的PNR。鲈鲤仔鱼初次摄食强度随饥饿天数的增加呈先上升后下降的变化趋势(图5),在5~11日龄期间(饥饿7 d内),仔鱼初次摄食强度和摄食能力随着饥饿天数的增加呈上升趋势,12日龄后(饥饿7 d以上)则随着饥饿天数的增加呈迅速下降趋势。
2. 5 饥饿组和摄食组鲈鲤仔鱼死亡率的变化情况
在18.5~21.9 ℃水温条件下,两组鲈鲤仔鱼的死亡率变化情况如图6所示。在13日龄前(饥饿9 d内),两组仔鱼的死亡率差异不明显,累计死亡率均小于10.00%;从17日龄(饥饿13 d)起饥饿组仔鱼的累计死亡率剧增,到18日龄(饥饿14 d)时饥饿组仔鱼的累计死亡率达88.00%,而摄食组仔鱼累计死亡率仅为14.75%;至19日龄(饥饿15 d)时饥饿组仔鱼的累计死亡率高达100.00%,摄食组仔鱼的累计死亡率仍为14.75%。饥饿组鲈鲤仔鱼的50.00%累计死亡率出现日龄为18日龄,即在PNR出现之后。
3 讨论
3. 1 饥饿对鲈鲤仔鱼生长及发育的影响
仔鱼期是鱼类早期生活史中最敏感、最脆弱的时期,极易受饵料、水温及水质等一系列外界因子的影响,其中饵料是影响早期仔鱼正常发育和存活的重要因素(李艳秋等,2014;陈淑吟等,2018),即由内源性营养顺利过渡到外源性营养是重要保障。本研究结果表明,饥饿对鲈鲤仔鱼的全长、体高等生长指标有极显著影响。在11日龄(摄食7 d)时,摄食组仔鱼生长速度加快,生物学指标增长显著,仔鱼整体较粗壮,而饥饿组仔鱼出现负增长,体高不断变小,呈头大身瘦的体形,两组仔鱼的全长、体高等生长指标变化差异极显著,与白甲鱼(李强等,2009)、太平洋鳕(李艳秋等,2014)、黑棘鲷(杨育凯等,2019)等鱼类的饥饿试验结果一致。此外,饥饿对鲈鲤仔鱼眼径和鳔组织的发育具有极显著影响,当内源性营养消耗殆尽又无外源性营养供给时,饥饿组仔鱼眼径和鳔组织等器官出现发育停滞甚至负增长的现象,与陈淑吟等(2018)针对黑鲷(♀)×真鲷(♂)杂交F2代仔鱼的研究结果相似。
3. 2 鲈鲤仔鱼混合营养期
鱼类仔鱼一般需经过内源性营养期、混合营养期和外源性营养期3个阶段。鱼类混合营养期的长短受其种类影响而存在明显差异。大西洋鳟(Salmo salar)仔鱼的混合营养期长达30 d(Hansen and M?ller,1985),哲罗鱼仔鱼的混合营养期约10 d(张永泉等,2009),黑莓鲈(Pomoxis nigromacufatus)仔鱼的混合营养期为2 d(秦志清,2014),而豹纹鳃棘鲈(Plectropomus leopardus)仔鱼的混合营养期仅为0.5 d(杨育凯等,2017)。本研究结果显示,鲈鲤仔鱼5日龄开口摄食,10日龄卵黄囊基本消耗殆尽,混合营养期约维持5 d。鱼类混合营养期除受其种类影响产生差异外,水温等环境因子对混合营养期也具有显著影响(万瑞景等,2004;单秀娟和窦硕增,2008)。为了减少水温及卵黄囊吸收速度等因素的影响,常采用仔鱼开口摄食至卵黄囊吸收完毕时长与仔鱼孵化至开口摄食时长之比评价不同种类仔鱼混合营养期的相对状态(宋勇等,2019):基准比值>1,表明仔鱼在卵黄囊吸收完毕前开口摄食,摄食能力较强,不易遭受饥饿胁迫威胁;基准比值<1,表明仔鱼卵黄囊已吸收完毕但尚未开口摄食,摄食能力较弱,易遭受饥饿胁迫威胁。本研究中,鲈鲤仔鱼开口摄食至卵黄囊吸收完毕的时长为5 d,仔鱼孵化至开口摄食的时长为5 d,基准比值为1,表明鲈鲤仔鱼摄食能力适中。此外,摄食组鲈鲤仔鱼在5日龄开口摄食后,其卵黄囊消耗速度相对变快,而饥饿组仔鱼由于未获得外源性营养,其卵黄囊吸收速度相对变慢。这可能是鲈鲤仔鱼面对饥饿胁迫作出的生理适应,与在大泷六线鱼(Hexagrammos otakii)(菅玉霞等,2014)、黑莓鲈(秦志清,2014)、黑棘鲷(杨育凯等,2019)上的研究结果相似。鲈鲤仔鱼初次摄食强度以12日龄为分界点,12日龄前初次摄食强度随着饥饿天数的增加呈上升趋势,12日龄后初次摄食强度随着饥饿天数的增加呈下降趋势,与李艳秋等(2014)对太平洋鳕仔鱼的研究结果相似。因此,要提高鱼苗成活率,育苗过程中应及时观察仔鱼卵黄囊吸收情况,准确把握开口摄食时间,适时进行投喂,避免饥饿胁迫影响仔鱼的生长发育及存活。
3. 3 鲈鲤仔鱼初次摄食率与PNR
仔鱼摄食能力常以初次摄食率和最高摄食率持续时间进行判定(殷名称,1996)。本研究结果显示,鲈鲤仔鱼5日龄(初次摄食)时的初次摄食率较低,仅为20%,随后迅速上升,13日龄时达最高初次摄食率(100%),维持1 d后初次摄食率开始下降,其最高摄食率持续时间较短,与条石鲷(Oplegnathus fasciatus)(彭志兰等,2010)、细鳞斜颌鲴(Plagiognathops microlepis)(叶忠平等,2012)等鱼类相似,但与赤眼鳟(Squaliobarbus curriculus)(李彩娟等,2016)存在明显差异。PNR是评价鱼类饥饿耐受力的重要指标之一,抵达PNR的时间越短表明仔鱼饥饿耐受力越弱,抵达PNR的时间越长表明仔鱼饥饿耐受力越强(黄海等,2012)。已有研究证实,鲢(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)、草鱼(Ctenopharyngodon idellus)仔鱼PNR均发生在16.5日龄(殷名称,1997),中华鲟(Acipenser sinensis)仔鱼PNR发生在24日龄(庄平等,1999),唐鱼(Tanichthys albonubes)仔鱼PNR发生在8.5日龄(陈国柱和方展强,2007),白甲鱼仔鱼PNR发生在13.5日龄(李强等,2009),哲罗鱼仔鱼PNR发生在39~40日龄(张永泉等,2009),条石鲷仔鱼PNR发生在2.5~3.0日龄(彭志兰等,2010),赤眼鳟仔鱼PNR发生在10~11日龄(李彩娟等,2016)。本研究结果显示,在18.5~21.9 ℃水温条件下鲈鲤仔鱼PNR发生在17日龄,与上述鱼类比较,鲈鲤仔鱼耐饥饿能力与鲢、鳙和草鱼相近,强于唐鱼、赤眼鳟、白甲鱼和条石鲷,但弱于中华鲟和哲罗鱼。陈国柱和方展强(2007)研究发现,可采用PNR有效积温参数客观评价不同鱼类的耐饥饿能力,且多数鱼类仔鱼的PNR有效積温范围在100~250 ℃·d。本研究中,鲈鲤仔鱼的PNR有效积温为347.4 ℃·d,高于多数鱼类仔鱼的PNR有效积温,说明鲈鲤仔鱼具有较强的耐饥饿能力。
3. 4 饥饿对鲈鲤仔鱼存活的影响及最佳投喂时间
饥饿是制约鱼类仔鱼早期存活率的主要因素之一。本研究结果表明,饥饿对鲈鲤仔鱼的存活影响显著,饥饿组仔鱼的累计死亡率在17日龄时迅速增加,至18日龄时高达88.00%,到19日龄时达100.00%,即鲈鲤仔鱼累计死亡率与饥饿时间呈正相关,与唐鱼(陈国柱和方展强,2007)、白甲鱼(李强等,2009)、细鳞斜颌鲴(叶忠平等,2012)等鱼类的研究结果一致。当仔鱼抵达PNR后才出现50.00%的累计死亡率,理论上只要在仔鱼抵达PNR前能得到适宜的饵料补给,就还有恢复的可能性(陈国柱和方展强,2007;李强等,2009)。本研究结果显示,饥饿组鲈鲤仔鱼的50.00%累计死亡率出现在PNR之后,提示在抵达PNR前获得适宜饵料对保障鲈鲤仔鱼的存活具有重要意义。鲈鲤仔鱼具有一定耐饥饿能力,在18.5~21.9 ℃水温条件下,虽然5日龄即开始摄食,但摄食率不高,直到8日龄时其摄食率才超过50%,摄食强度也保持稳步上升;摄食率和摄食强度分别在13日龄和11日龄开始下降。因此,鲈鲤仔鱼可在5日龄后开始投喂,最佳投喂时间为8日龄,但最晚投喂时间不宜超过13日龄。
4 结论
在18.5~21.9 ℃水温条件下,鲈鲤仔鱼混合营养期为5 d,PNR发生在17日龄。鲈鲤仔鱼可在5日龄后开始投喂,最佳投喂时间为8日龄,但最晚投喂时间不宜超过13日龄。
参考文献:
陈国柱,方展强. 2007. 饥饿对唐鱼仔鱼摄食和生长的影响[J]. 动物学杂志,42(5):49-61. [Chen G Z,Fang Z Q. 2007. Effects of starvation on feeding and growth of the larvae of Tanichthys albonubes[J]. Chinese Journal of Zoology,42(5):49-61.]
陈淑吟,张志勇,许津,吉红九,吴国均,贾超峰,祝斐,孟乾. 2018. 黑鲷(♀)×真鲷(♂)杂交子二代仔鱼早期发育及饥饿不可逆点[J]. 南方水产科学,14(4):122-128. [Chen S Y,Zhang Z Y,Xu J,Ji H J,Wu G J,Jia C F,Zhu F,Meng Q. 2018. Study on larval early development and point of no return of hybrid F2 of Acanthopagrus schiegelii(♀)×Pagrus major(♂)[J]. South China Fishe-ries Science,14(4):122-128.]
段国庆,江河,胡王,凌俊,胡玉婷. 2013. 黄鳝仔鱼饥饿试验与不可逆点的确定[J]. 南方农业学报, 44(6):1036-1040. [Duan G Q,Jiang H,Hu W,Ling J,Hu Y T. 2013. Experimental starvation on Monopterus albus larvae and determination of the point of no return[J]. Journal of Southern Agriculture,44(6):1036-1040.]
黄海,杨宁,张希. 2012. 马拉瓜丽体鱼仔鱼饥饿试验及不可逆点的确定[J]. 南方水产科学,8(2):43-50. [Huang H,Yang N,Zhang X. 2012. Experimental starvation on Cichlasoma managuense larvae and determination of point of no return[J]. South China Fisheries Science,8(2):43-50.]
贾瑞锦,陈超,孔祥迪,李炎璐,宋振鑫,吴雷明,王鲁,曲江波,赵从明,吴坚. 2014. 饥饿对条纹锯鮨仔鱼的存活、摄食及生长的影响[J]. 海洋科学,38(4):44-49. [Jia R J,Chen C,Kong X D,Li Y L,Song Z X,Wu L M,Wang L,Qu J B,Zhao C M,Wu J. 2014. Effects of starvation on suriving,feeding and growth of the yolk-sac larval of Centropristis striata[J]. Marine Sciences,38(4):44-49.]
菅玉霞,房慧,张少春,王雪,胡发文,高凤祥,潘雷,郭文. 2014. 大泷六线鱼仔鱼饥饿试验及不可逆点的研究[J]. 海洋科學,38(3):111-115. [Jian Y X,Fang H,Zhang S C,Wang X,Hu F W,Gao F X,Pan L,Guo W. 2014. Starvation and no return point study of Hexagrammos otakii Jordan et Starks larvae[J]. Marine Sciences,38(3):111-115.]
李彩娟,许郑超,张振早,王辉,凌去非. 2016. 赤眼鳟仔鱼饥饿试验和不可逆点研究[J]. 扬州大学学报(农业与生命科学版), 37(3):66-70. [Li C J,Xu Z C,Zhang Z Z,Wang H,Ling Q F. 2016. Starvation and point of no return of Squaliobarbus curriculus larvae[J]. Journal of Yangzhou University (Agricultural and Life Science Edition),37(3):66-70.]
李强,李孟均,周传江,陈先均. 2009. 饥饿对白甲鱼(Onychostoma sima)仔鱼摄食、生长的影响[J]. 淡水渔业,39(5):32-37. [Li Q,Li M J,Zhou C J,Chen X J. 2009. Effects of starvation on feeding and growth of the larvae of Onychostoma sima[J]. Freshwater Fisheries,39(5):32-37.]
李艳秋,姜志强,孙阳,毛明光,孟祥科. 2014. 太平洋鳕仔鱼饥饿实验及不可逆生长点的确定[J]. 生态学报,34(14):3873-3878. [Li Y Q,Jiang Z Q,Sun Y,Mao M G,Meng X K. 2014. Experimental starvation on Gadus macrocephalus and definition of the point of no return[J]. Acta Ecologica Sinica,34(14):3873-3878.]
劉军. 2004. 长江上游特有鱼类受威胁及优先保护顺序的定量分析[J]. 中国环境科学,24(4):395-399. [Liu J. 2004. A quantitative analysis on threat and priority of conservation order of the endemic fishes in upper reaches of the Yangtze River[J]. China Environmental Science,24(4):395-399.]
马建波,茅沈丽,梁桂洪,邹记兴. 2012. 杂交鳢仔鱼饥饿试验及不可逆点的确定[J]. 水生态学杂志,33(3):90-95. [Ma J B,Mao S L,Liang G H,Zou J X. 2012. Starvation and point of no return in hybrid snakehead larvae[J]. Journal of Hydroecology,33(3):90-95.]
彭志兰,柳敏海,罗海忠,傅荣兵,张凤萍,毛志增. 2010. 条石鲷仔鱼饥饿试验及不可逆点的确定[J]. 水产科学,29(3):152-155. [Peng Z L,Liu M H,Luo H Z,Fu R B,Zhang F P,Mao Z Z. 2010. Starvation and point of no return in striped knifejaw Oplegnathus fasciatus larvae[J]. Fisheries Science,29(3):152-155.]
秦志清. 2014. 黑莓鲈(Pomoxis nigromacufatus)仔鱼饥饿试验及PNR的确定[J]. 福建农业学报,29(12):1167-1173. [Qin Z Q. 2014. Experimental starvation on Pomoxis nigromacufatus larvae and definition of the point of no return[J]. Fujian Journal of Agricultural Sciences,29(12):1167-1173.]
单秀娟,窦硕增. 2008. 饥饿胁迫条件下黑鮸(Miichthys miiuy)仔鱼的生长与存活过程研究[J]. 海洋与湖沼, 39(1):14-23. [Shan X J,Dou S Z. 2008. Effects of delayed first feeding on growth,survival and biochemical composition of croaker Miichthys miiuy larvae[J]. Ocea-nologia et Limnologia Sinica,39(1):14-23.]
宋勇,江志香,马卓勋,任道全,陈生熬. 2019. 叶尔羌高原鳅仔鱼饥饿试验及不可逆点的确定[J]. 水生态学杂志,40(2):102-107. [Song Y,Jiang Z X,Ma Z X,Ren D Q,Chen S A. 2019. Starvation experimental on Triplophysa yarkandensis larvae and the determination of the point of no return(PNR)[J]. Journal of Hydroecology,40(2):102-107.]
万瑞景,李显森,庄志猛,蒙子宁. 2004. 鳀鱼仔鱼饥饿试验及不可逆点的确定[J]. 水产学报,28(1):79-83. [Wan R J,Li X S,Zhuang Z M,Meng Z N. 2004. Experimental starvation on Engraulis japonicus larvae and definition of the point of no return[J]. Journal of Fisheries of China,28(1):79-83.]
王晓龙,温海深,张美昭,李吉方,张凯强,王伟,刘阳,田源,常志成. 2017. 花鲈初孵仔鱼饥饿不可逆点的确定及摄食节律研究[J]. 中国海洋大学学报(自然科学版),47(5):57-64. [Wang X L,Wen H S,Zhang M Z,Li J F,Zhang K Q,Wang W,Liu Y,Tian Y,Chang Z C. 2017. Determination of irreversible starvation point and feeding rhythm of Lateolabrax maculatus larvae[J]. Periodical of Ocean University of China,47(5):57-64.]
夏连军,施兆鸿,陆建学. 2004. 黄鲷仔鱼饥饿试验及不可逆点的确定[J]. 海洋渔业,26(4):286-290. [Xia L J,Shi Z H,Lu J X. 2004. Experimental starvation on Dentex tumifrons larvae and definition of the point of no return[J]. Marine Fisheries,26(4):286-290.]
杨育凯,林黑着,王志勇,谢仰杰,郭奕惠,黄小林,李涛. 2019. 黑棘鲷仔鱼饥饿实验及不可逆点的确定[J]. 渔业科学进展,41(2):87-94. [Yang Y K,Lin H Z,Wang Z Y,Xie Y J,Guo Y H,Huang X L,Li T. 2019. Experimental starvation of Acanthopagrus schlegelii larvae,and de-finition of the point of no return[J]. Progress in Fishery Sciences,41(2):87-94.]
杨育凯,虞为,林黑着,李涛,黄小林,黄忠,戚常乐. 2017. 豹纹鳃棘鲈仔鱼饥饿实验和不可逆点研究[J]. 南方水产科学, 13(6):90-96. [Yang Y K,Yu W,Lin H Z,Li T,Huang X L,Huang Z,Qi C L. 2017. Experimental starvation of Plectropomus leopardus larvae and study of point of no return[J]. South China Fisheries Science,13(6):90-96.]
叶忠平,吕业坚,林岗. 2012. 细鳞斜颌鲴仔鱼饥饿试验与不可逆点的确定[J].南方农业学报,43(4):536-539. [Ye Z P,Lü Y J,Lin G. 2012. Experiments on starvation of Plagiognathops microlepis larvae and its PNR determination[J]. Journal of Southern Agriculture,43(4):536-539.]
殷名称. 1996. 鱼类早期生活史阶段的自然死亡[J]. 水生生物学报,20(4):363-372. [Yin M C. 1996. Natural mortality of early life stages of fish[J]. Acta Hydrobiologica Sinica,20(4):363-372.]
殷名称. 1997. 鲢、鳙、草、银鲫卵黄囊仔鱼的摄食生长、耐饥饿能力[C]//中国鱼类学会. 鱼类学论文集(第六辑). 北京:科学出版社:69-79. [Yin M C. 1997. Feeding,growth and starvation tolerance of the yolk-sac larvae of Hypophthalmichthys molitrix,Hypophthalmichthys nobilis,Cten-opharyngodon idellus and Carassius auratus gibelio[C]// Chinese Ichthyological Scoiety. Proceedings of Fish Scien-ce(Vol. 6). Beijing: Science Press: 69-79.]
詹會祥,杨德国,李正友,王艳艳,周礼敬,吴兴兵,杨林. 2016. 金沙鲈鲤人工繁殖技术研究[J]. 水生态学杂志,37(4):84-88. [Zhan H X,Yang D G,Li Z Y,Wang Y Y,Zhou L J,Wu X B,Yang L. 2016. Artificial propagation of Percocypris pingi[J]. Journal of Hydroecology,37(4):84-88.]
张永泉,尹家胜,杜佳,贾钟贺,佟广香. 2009. 哲罗鱼仔鱼饥饿实验及不可逆生长点的确定[J].水生生物学报, 33(5):945-950. [Zhang Y Q,Yin J S,Du J,Jia Z H,Tong G X. 2009. ExperImental starvatian on Hucho taimen and definition of the point of no return[J]. Acta Hydrobio-logica Sinica,33(5):945-950.]
庄平,章龙珍,张涛,张征,曹文宣. 1999. 中华鲟仔鱼初次摄食时间与存活及生长的关系[J]. 水生生物学报,23(6):560-565. [Zhuang P,Zhang L Z,Zhang T,Zhang Z,Cao W X. 1999. Effects of delaying first feeding time on the survival and growth of larval Chinese sturgeon,Acipen-ser sinensis[J]. Acta Hydrobiologica Sinica,23(6):560-565.]
Hansen T J,M?ller D. 1985. Yolk absorption,yolk sac constrictions,mortality,and growth during first feeding of Atlantic salmon(Salmo salar) incubated on astro-turf[J]. Canadian Journal of Fisheries and Aquatic Sciences,42(6):1073-1078.
(责任编辑 兰宗宝)
