Bergmann’s rule and Allen’s rule in two passerine birds in China
2020-01-11LiqingFanTianlongCaiYingXiongGangSongandFuminLei
Liqing Fan, Tianlong Cai, Ying Xiong, Gang Song and Fumin Lei,4*
Abstract
Keywords: Appendage size, Body size, Geographic variation, Thermoregulation
Background
Bergmann’s rule (1847), one of the best-known ecogeographic patterns, states that populations or closely related species of endotherms typically have a larger body size in colder climates as a result of thermoregulation (James 1970; Salewski and Watt 2017). Bergmann proposed the conventional heat conservation hypothesis to explain this pattern, which assumed that the volume of endothermic animals is a limiting factor for heat production and the surface area is a limiting factor for heat dissipation. Further, he posited that the lower surface area to volume ratio occurring in the larger body size endotherms would facilitate heat retention, which is favored in lower temperature environments or at higher latitudes (Mayr 1956;James 1970; Salewski and Watt 2017). Environmental factors other than temperature have been proposed as contributing to geographic variations in body size, such as humidity (James’s hypothesis) (James 1970), seasonality (the seasonality hypothesis) (Boyce 1979; Searcy 1980;Lindstedt and Boyce 1985) and precipitation (the primary productivity hypothesis) (Rosenzweig 1968).
There is no agreement about whether Bergmann’s pattern is general or valid (Blackburn et al. 1999; Meiri and Dayan 2003; Watt et al. 2010; Teplitsky and Millien 2014).Empirical studies have found the predicted pattern at both the intraspecific and interspecific levels (Meiri 2011;Salewski and Watt 2017) in numerous mammals (Ashton et al. 2000; Rodriguez et al. 2008) and birds (Olson et al.2009; Sun et al. 2017). At the same time, animals that do not conform to Bergmann’s rule have also been reported(Yom-Tov et al. 2002; Freeman 2017; Sargis et al. 2018).For instance, decreases in body mass with increasing temperature within species have been found in only 14%of 952 endothermic species (Riemer et al. 2018).
Allen’s rule (1877) is an extension of Bergmann’s rule,predicting that appendage size in endotherms, including limbs, tails and ears, become larger in warm climates for similar thermoregulatory reasons. Studies have found that this pattern is valid for some mammals (Yom-Tov and Yom-Tov 2005; Betti et al. 2015) and birds (Laiolo and Rolando 2001; VanderWerf 2012), but not in some cases (Wiedenfeld 1991; Bidau et al. 2011; Du et al. 2017).One of the most representative examples of Allen’s rule is bill size, which has been demonstrated to serve as an efficient radiator in birds (Scott et al. 2008; Tattersall et al. 2009; Campbell-Tennant et al. 2015). Smaller bills with less surface area help birds keep a constant temperature in colder environments, while the larger surface area of larger bills in warmer climates raises the effi-ciency of heat dissipation (Symonds and Tattersall 2010).For instance, Tattersall et al. (2016) found that 64 of 110 bird species have a smaller bill at cooler environments at the intraspecific level. In addition to bills, the featherless legs of birds are also reported to play a role in heat exchange (Martineau and Larochelle 1988; Arad et al.1989; Maloney and Dawson 1994) and several studies have revealed a profound effect of temperature on tarsus length (Laiolo and Rolando 2001; VanderWerf 2012). For example, Nudds and Oswald (2007) found that terns and gulls showed interspecific geographic variation predicted by Allen’s rule in the length of exposed leg elements, but not in feathered element length.
For animals living in seasonal climates, the need for dissipating heat in the summer may tradeoff with the need for conserving heat in the winter. It remains unclear which season is the critical period for thermoregulation.Studies have found that selection on bill size for its thermoregulation function varies with climatic regions. For instance, bill size of Song Sparrows (Melospiza melodia)increases with high summer temperatures in California(Greenberg and Danner 2012) and decreases with cold winter temperatures in eastern North America (Danner and Greenberg 2015), suggesting that variation in bill size is selected for summer heat dissipation and winter heat retention in these two regional climates respectively.Friedman et al. (2017) further found that beak size across 158 Australasian species was positively correlated with winter minimum temperature but not with summer maximum temperature.
Bergmann emphasized that it would be easier to find a geographic size cline among similar animals (Salewski and Watt 2017). Therefore, intraspecific comparisons are powerful in testing the relationships between climate and body trait variations (Mayr 1956; Shelomi 2012). The Oriental Magpie (Pica serica) and the Oriental Tit (Parus minor) are species within, respectively, Pica pica (Haring et al. 2007; Lee et al. 2003; Zhang et al. 2012; Song et al.2018) and Parus major (Kvist et al. 2003; Päckert et al.2005) complex. Oriental Magpies and Oriental Tits are widespread across eastern and central China, abundant and resident or dispersed at short distances, providing two ideal groups to test Bergmann’ rule and Allen’s rule at the intraspecific level. In this study, we examined the geographical variations of several major morphological traits in Oriental Magpies and Oriental Tits to evaluate whether they support Bergmann’s rule and Allen’s rule.We also test correlation between morphological variation and climate variables, including temperature, precipitation and seasonality, to further explain the mechanisms of these two rules.
Methods
Data collection
We measured wing length (carpal joint to the tip of the longest primary feather unflattened, ± 0.01 mm), total bill length (± 0.01 mm) and tarsus length (joint of tibiotarsus and tarsometatarsus to the distal edge of the last undivided scute on the anterior surface of the leg,± 0.01 mm) of 165 specimens of Oriental Magpies and 410 specimens of Oriental Tits from collections of the Zoological Museum of Institute of Zoology, Chinese Academy of Sciences. All measurements were performed by Liqing Fan. Data corresponding to juveniles or molting adults was excluded from analysis. Specimens of Oriental Magpie were collected from 21°27.5′N to 40°59.0′N at latitude and 0 to 3500 m a.s.l. at elevation between 1953 and 2009 at 65 localities (Additional file 1: Table S1), and Oriental Tits were collected from 23°9.6′N to 47°1.7′N at latitude and 0 to 1565 m a.s.l. at elevation between 1951 and 2009 at 52 localities in Chinese mainland (Additional file 2: Table S2).
Body mass (± 1 g), location data and collection date were obtained from the specimen labels. We georeferenced the site at which each specimen was captured and obtained the climate variables for each locality from WorldClim Version 1.4 (http://www.world clim.org/) for 30-year means (1960-1990) with 30 s spatial resolution(Hijmans et al. 2005), including annual mean temperature(bio1), temperature seasonality (bio4), maximum temperature of warmest month (bio5), minimum temperature of coldest month (bio6), temperature annual range(bio7), mean temperature of warmest quarter (bio10),mean temperature of coldest quarter (bio11), annual precipitation (bio12), precipitation seasonality (bio15), precipitation of warmest quarter (bio18) and precipitation of coldest quarter (bio19).
Statistical analyses
We checked the relationships of body size (body mass and wing length) and appendage size (bill length and tarsus length) with the geographic and environmental variables. We used bio1, bio4-7, bio10-12, bio15, bio18 and bio19 to test for Bergmann’s rule, and used bio1, bio5,bio6, bio10 and bio11 for Allen’s rule.
Traits in males and females may show different geographic patterns due to sexual selection and sexual dimorphism (Mccollin et al. 2015), and the effect of sex on morphological measurements was analyzed with linear mixed-effect models (LME) using the restricted maximum likelihood (REML) method (fixed effect: sex,random effects: collection site) (Pinheiro et al. 2015). If the bird displayed sexual dimorphism in morphological measurements, data were analyzed separately for each sex. We checked the correlations of the traits in each sex.Then, we regressed bill length and tarsal length against wing length and extracted the residuals from the models(residual of bill length/tarsus length). The residuals were subsequently used as size-independent appendage variables. Linear mixed-effect models were also performed to examine the impact of latitude, elevation and climate variables on each morphological measurement with collection site as random effect. In models with multiple climate variables as predictors, we used Akaike’s Information Criterion (AIC, Burnham and Anderson 2002)to identify the best mode with the lowest AIC score. We then performed Collinearity Diagnostics and discarded the models if the variance inflation factor > 10, eigenvalue< 0.05 or condition index > 10. All statistical analysis were conducted using R 3.4.4. (R Development Core Team 2012).
Results
Male Oriental Magpies were significantly larger than females in all of the morphological measurements (all t ≤ - 6.153, p < 0.001; Table 1). In Oriental Tits, males did not differ from females in body mass (t = - 1.117,p = 0.264) and bill length (t = - 0.269, p = 0.788), but the males had larger wing length (t = - 8.147, p < 0.001)and tarsus length (t = - 3.218, p = 0.001; Table 1) thanthe females. Therefore, data were analyzed separately for each sex for both species.
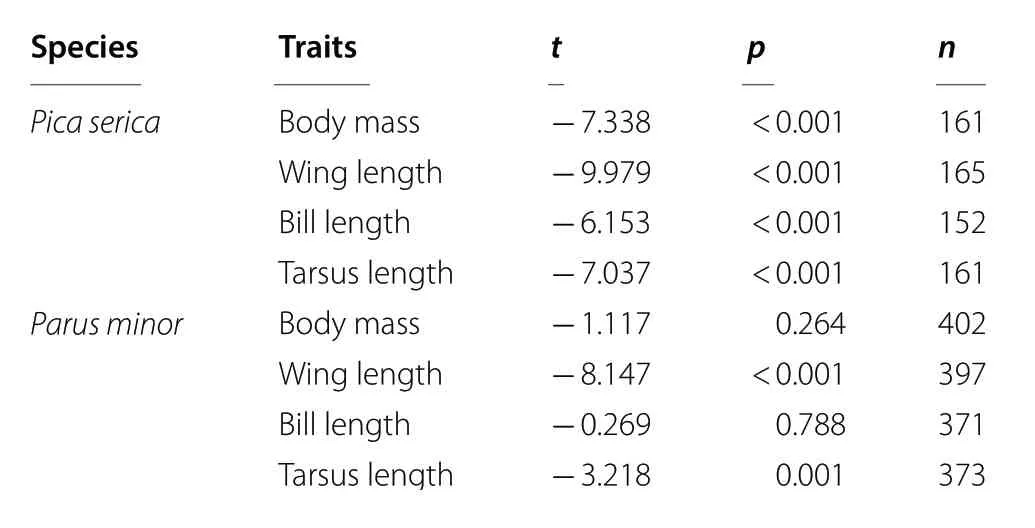
Table 1 Effect of sex on morphological measurements in the Oriental Magpie (Pica serica) and the Oriental Tit(Parus minor)
Oriental Magpies
There were significant positive correlations between each pair of morphological measurements in both sexes for Oriental Magpies (all Spearman’s rank Correlation r ≥ 0.236, p < 0.05; Table 2) except correlations between wing length and bill length, and correlations between wing length and tarsus length in males (both p > 0.05;Table 2).
We did not find support for Bergmann’s rule in magpies. Neither body mass nor wing length in either sex—excepting wing length of males, which increased with elevation significantly (t = 2.177, p = 0.030)—showed latitudinal or elevational cline, or related to climate variables(Fig. 1; Table 3).
There were no significant relationships between residual of tarsus length in males and bio5 (maximum temperature of warmest month) or bio10 (mean temperature of warmest quarter), nor between residual of tarsus length in females and bio5. Beyond that, residual of bill length and residual of tarsus length in both sexes all significantly decreased with latitude (all t ≤ -4.42, p < 0.001;Fig. 1) and increased with temperature variables (all t ≥ 2.173, p < 0.05; Table 4), concordant with the patterns predicted by Allen’s rule. In all of these significant relationships, temperature variables explained 9.9-38.1%of all variations in bill length, and 8.1-36.6% of all variations in tarsus length (Table 4). Residual of bill length and residual of tarsus length in both sexes did not vary with elevation with the exception of residual of bill length in females (t = - 2.757, p = 0.006; Table 4). According to the AIC value and Collinearity Diagnostics, the bestfit models explaining geographic variation in bill length for male and female magpies included, respectively,bio6 (minimum temperature of coldest month) and bio1(annual mean temperature), and accounted for 38.1% and 25.6% of the total variance. The best predictors for tarsus length in males and females were bio6 and bio11 (meantemperature of coldest quarter), respectively, explaining 22.6% and 36.6% of the total variance (Table 4; Additional file 3: Table S3).
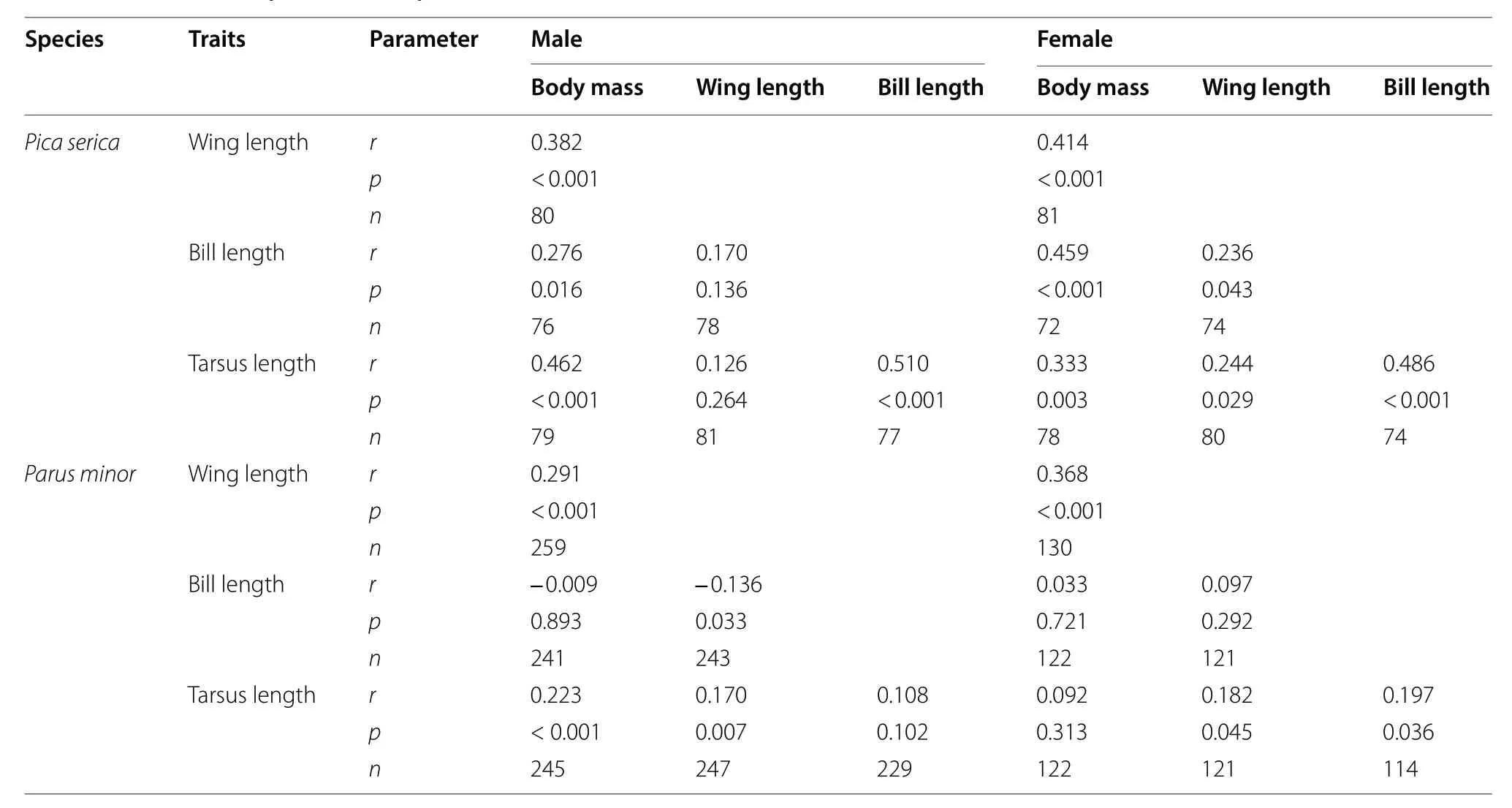
Table 2 Spearman’s rank correlation between morphological measurements in the Oriental Magpie (Pica serica)and the Oriental Tit (Parus minor)
Oriental Tits
In male Oriental Tits, body mass was positively correlated with wing length (Spearman’s rank correlation r = 0.291,p < 0.001) and tarsus length (r = 0.223, p < 0.001), and wing length was correlated with bill length (r = - 0.136,p = 0.033) and tarsus length (r = 0.170, p = 0.007). In females, body mass was correlated with wing length(r = 0.368, p < 0.001), and tarsus length was correlated with wing length (r = 0.182, p = 0.045) and bill length(r = 0.197, p = 0.036; Table 2).
Body mass in tits in both sexes showed little variation along latitudinal, elevational or the environmental gradients (all p > 0.05). In both sexes, wing length was significantly related to latitude (both t ≥ 9.536, p < 0.001)and the climate variables (all |t| ≥ 2.305, p < 0.05; Fig. 2;Table 3). Residual of bill length and residual of tarsus length were not related to latitude, elevation or the climate variables in either sex (Fig. 2; Table 4). According to the AIC value and Collinearity Diagnostics, the primary driver of wing length of tits for both sexes was bio6 (minimum temperature of coldest month; Table 3; Additional file 3: Table S3), explaining 38.2% and 36.5% of the variances for males and females respectively.
Discussion
Our results show that body size of Oriental Magpies did not follow Bergmann’s rule, as no geographic variation was found. Nevertheless, appendage size (bill length and tarsus length) of Oriental Magpies tend to be smaller in higher latitudes, concordant with Allen’s rule. Whereas Oriental Tits followed Bergmann’ rule instead of Allen’s rule, with longer wings at higher latitudes. The different patterns of morphological traits with latitudinal gradients in these two birds suggest that species may adopt different thermoregulation strategies to adapt to environment. Similar findings have been reported previously,for example, the Red-billed Chough (Pyrrhocorax pyrrhocorax) and the Alpine Chough (Pyrrhocorax graculus) follow both Bergmann’s rule and Allen’s rule (Laiolo and Rolando 2001), and the Red Squirrel (Tamiasciurus hudsonicus) follows Allen’s rule but not Bergmann’s rule(Lindsay 1987), whereas the Wedge-tailed Shearwater(Puffinus pacificus) conforms to Bergmann’s rule rather than to Allen’s rule (Bull 2006). In addition to changes in body size or appendage size, changes in the thickness of fur or feathers and in certain behaviors (panting, bathing,ptilo-erection and microsite selection) could be effective mechanisms that allow animals to cope with temperature gradients (Scholander 1955; Hafez 1964). In this study,the decline of appendage size in Oriental Magpies with the decrease in temperature may help the magpies retain heat in cold climates; this would have an effect similar to the increase in body size in Oriental Tits in cold temperatures.
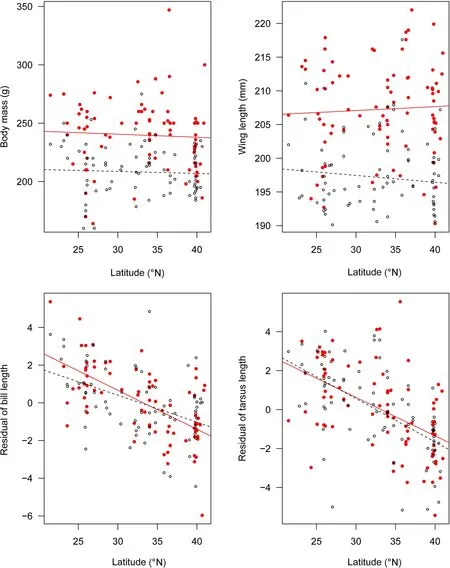
Fig. 1 Linear regressions between the morphological measurements and latitude in male (filled red circles and solid lines) and female (hollow circles and dash lines) Oriental Magpie (Pica serica)
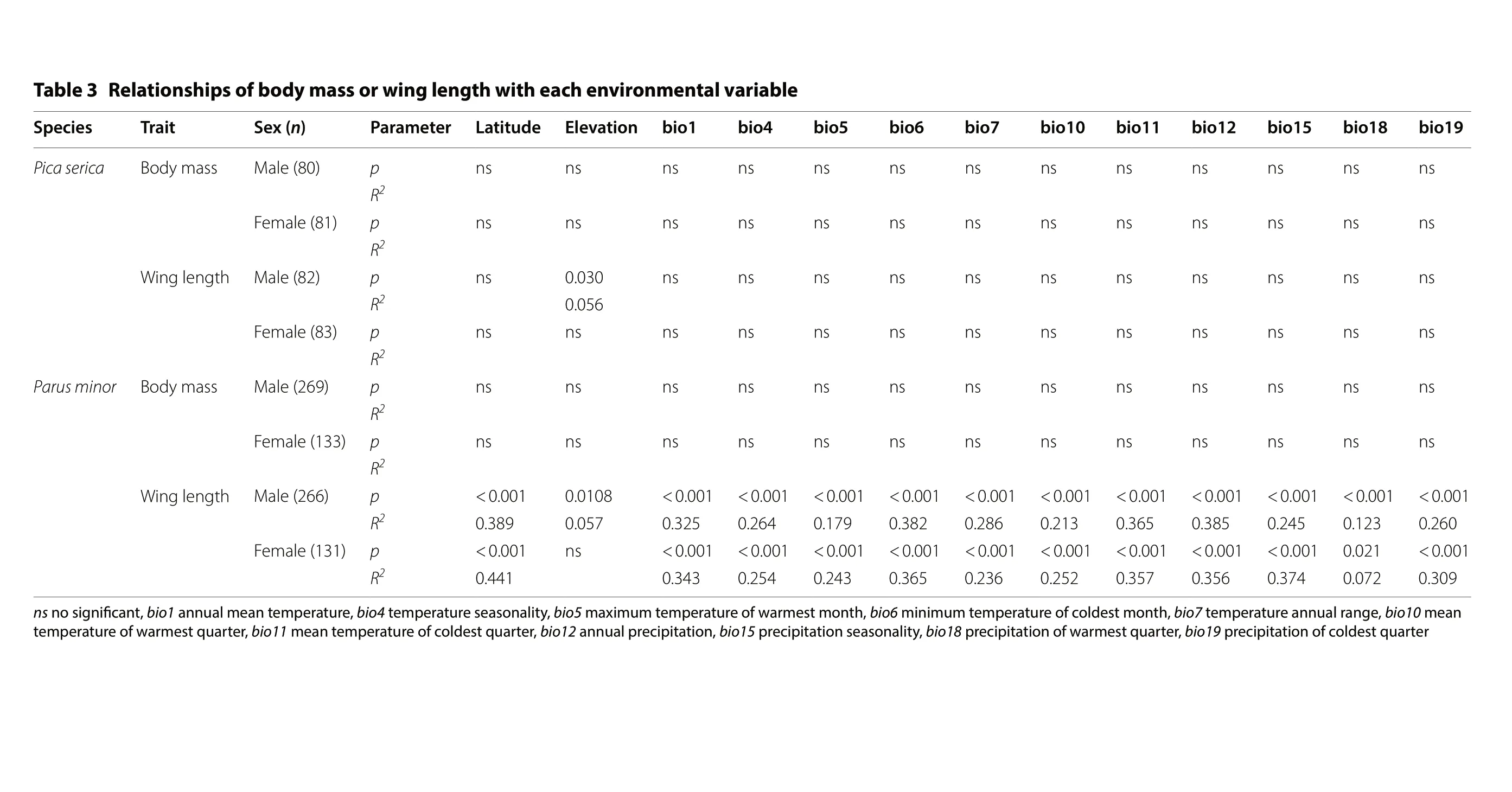
el b air a v l at n e m n ori v n e hc a e hti w ht g n el g ni w r o ss a my d o b f o s pi hs n oit al e R 3 el b aT 9 1 oi b 8 1 oi b 5 1 oi b 2 1 oi b 1 1 oi b 0 1 oi b 7 oi b 6 oi b 5 oi b 4 oi b 1 oi b n oitavel E e d utit aL r et e m ara P)n( x e S tiarT s eic e p S s n s n s n s n s n s n s n s n s n s n s n s n s n p)08( ela M ssa m y d o B acires aciP 2R s n s n s n s n s n s n s n s n s n s n s n s n s n p)18( ela meF 2R s n s n s n s n s n s n s n s n s n s n s n 030.0 s n)28( ela M ht g nelg ni W p 2 650.0 R s n s n s n s n s n s n s n s n s n s n s n s n s n)38( ela meF p2R s n s n s n s n s n s n s n s n s n s n s n s n s n)962( ela M ssa m y d o B p roni m suraP 2R s n s n s n s n s n s n s n s n s n s n s n s n s n)331( ela meF p2R 100.0<100.0<100.0<100.0<100.0<100.0<100.0<100.0<100.0<100.0<100.0<8010.0 100.0<p)662( ela M ht g nelg ni W 2 062.0 321.0 542.0 583.0 563.0 312.0 682.0 283.0 971.0 462.0 523.0 750.0 983.0 R 100.0<120.0 100.0<100.0<100.0<100.0<100.0<100.0<100.0<100.0<100.0<s n 100.0<p)131( ela meF 2 903.0 270.0 473.0 653.0 753.0 252.0 632.0 563.0 342.0 452.0 343.0 144.0 R nae m 01 oi b ,e g nar la u n na er utare p m et 7 oi b ,ht n o m tse dl oc f o er utare p m etm u mi ni m 6 oi b ,ht n o m tse mra w f o er utare p m etm u mixa m 5 oi b ,ytila n osa eser utar e p met 4 oi b ,er utare p met nae m la u n na 1 oi b ,t nacifi n gis o n s n r etra u q ts e dl oc f on oitati picer p 91 oi b ,r etra u q tse mra w f on oitati pic er p 81 oi b ,ytila n osaesn oitati picer p 51 oi b ,n oitati pic er p la u n na 21 oi b ,retra u q tse dl oc f o er utare p m et nae m 11 oi b ,retra u q ts e mra w f o er utare p met
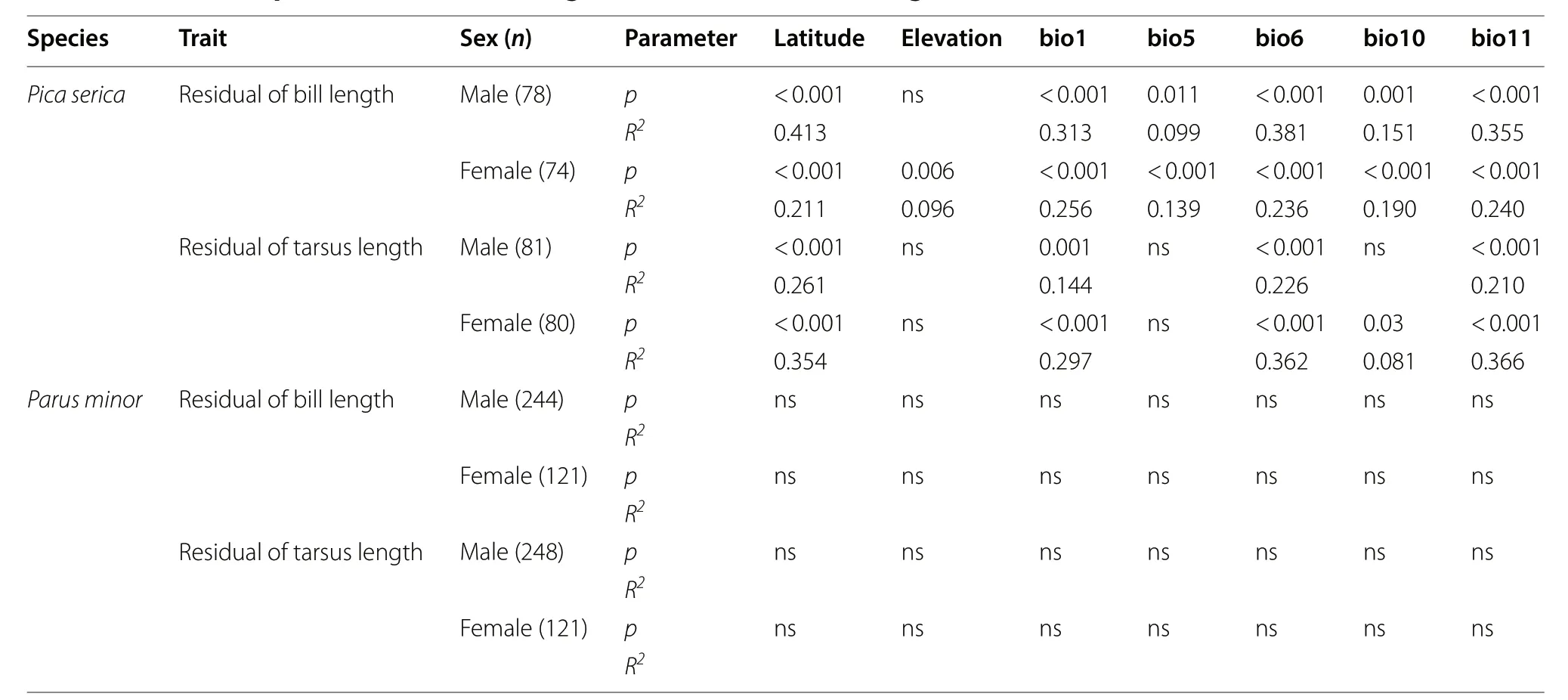
Table 4 Relationships of residual of bill length or residual of tarsus length with each environmental variable
For Oriental Magpies, appendage size in males and tarsus length in females were most strongly related to bio6(minimum temperature of coldest month) and bio11(mean temperature of coldest quarter) respectively, and both were less or not related to bio5 (maximum temperature of warmest month) and bio10 (mean temperature of warmest quarter; see Table 4). These results indicate that heat retention in winter cold environments, especially in the coldest month, rather than heat dissipation in summer warm environments, drives appendage size in magpies. The seasonal climates may have distinct influences on fitness (Greenberg and Danner 2012; Danner and Greenberg 2015). Compared with the relatively cool summer (19.834.8 °C for bio5), Oriental Magpies in this study suffered a hard winter (as low as - 22.7 °C for bio6) in the high latitude areas. Similar to the study on bill length of song sparrows in eastern North America(Danner and Greenberg 2015), winter was the season of critical thermal stress for Oriental Magpies of Chinese mainland, as indicated by their appendage size.
Body size of Oriental Tits, like appendage size of Oriental Magpies, was closely related to climate conditions in the winter. Wing length of tits in both sexes were driven by bio6 (minimum temperature of coldest month;Table 3), supporting the heat conservation hypothesis (Mayr 1956; James 1970). Environmental variables were highly correlated with each other for Oriental Tits in this study (Additional file 4: Table S4), and bio6 explained nearly as much of the variance of wing length in males as bio12 (annual precipitation; 38.2% vs 38.5%)and of the variance of wing length in females as bio15(precipitation seasonality; 36.5% vs 37.4%; Table 3).Precipitation is a major limiting factor for net primary productivity, and increased precipitation elevates net primary productivity. According to the primary productivity hypothesis, increase of net primary productivity would lead to an increase in food availability and thus an increase in body mass (Rosenzweig 1968; Yom-Tov and Geffen 2011). While, wing length in male tits decreased with bio12 (t = - 11.9; Additional file 3: Table S3), showing an opposite pattern predicted by the primary productivity hypothesis. Wing length in female tits increased with bio15 (t = 6.863; Additional file 3: Table S3), consistent with the seasonality hypothesis, which suggests that a larger body size would allow for higher fasting endurance and would be advantageous in regions with greater seasonality (Boyce 1979). These results indicate that temperature in the winter (bio6) and seasonality (bio15)limit natural selection for tits living at high latitudes, who become larger to retain their heat and survive food shortages in the winter.
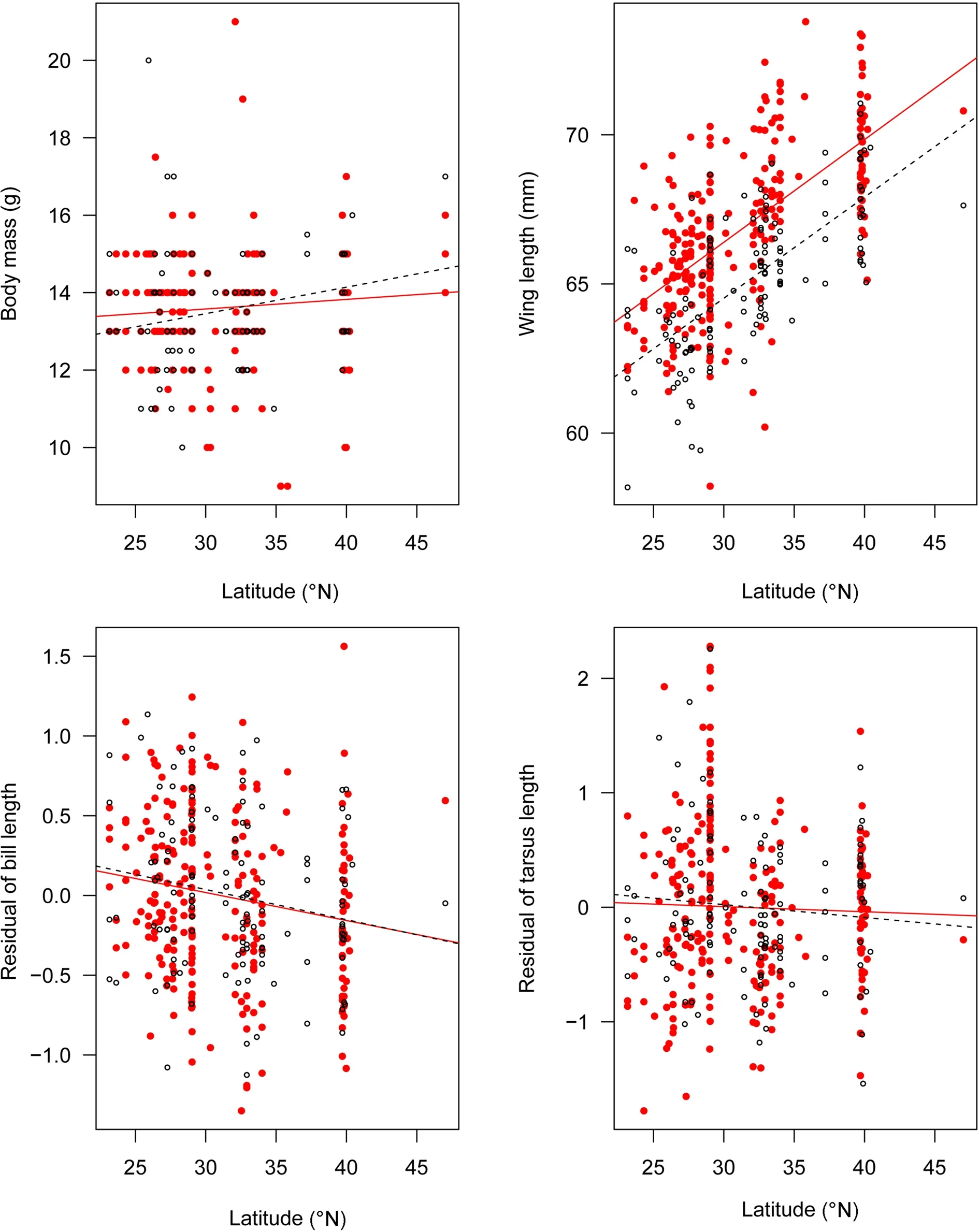
Fig. 2 Linear regressions between the morphological measurements and latitude in male (filled red circles and solid lines) and female (hollow circles and dash lines) Oriental Tit (Parus minor)
Body mass and wing length are widely used as proxies for bird body size (Snow 1954; Gosler et al. 1998).In Oriental Tits, body mass provided little evidence for Bergmann’s rule. In contrast, wing length showed clear patterns of Bergmann’s rule in both sexes, with temperature being the main driver of the variance. It is not surprising that we did not detect a strong geographic cline in body mass, given that the samples we used in this study were collected throughout the year (Additional file 2:Table S2), and, as we know, body mass fluctuates considerably during the breeding cycle (Croxall and Ricketts 1983) and seasonally (Marks and Leasure 1992; Yom-Tov et al. 2002). Besides, Oriental Tits are resident but normally undergo seasonal elevational movements, which may mitigate the potential climatic effects on morphology. In this study, wing length is a satisfactory indicator of body size for Oriental Tits, for they do not disperse over long distances.
Only a few morphological measurements (wing length of male magpies, residual of bill length of female magpies and wing length of male tits) showed an elevational cline,all with only a little part of the total variance attributed to elevation (R2≤ 9.6%; Tables 3, 4). Temperature varies along both elevational gradients and latitudinal gradients.Nevertheless, the variation of some climate conditions,such as air pressure and solar radiation, with elevation are entirely different from those that occur with latitude(Körner 2007). Therefore, biological traits along elevational gradients do not necessarily show similar clines as those along latitudinal gradients (Zhang and Lu 2012;Hille and Cooper 2015). For example, body mass of Torrent Ducks (Merganetta armata) followed Bergmann’s rule along latitudinal gradients but not along elevational gradients (Gutierrez-Pinto et al. 2014). And body mass and wing length of Eurasian Tree Sparrow (Passer montanus) showed an opposite pattern, increasing with elevation but not with latitude (Sun et al. 2017). More specimens from high elevations should be studied to further investigate elevational variation in morphological measurements.
Conclusions
Oriental Magpies followed Allen’s rule in relation to variation in bill length and tarsus length across latitudes,and latitudinal variation in wing length in Oriental Tits supported Bergmann’s rule. Minimum temperature of coldest month (bio6) was the best climate variable that predicted geographic variation in bill length and tarsus length in male Oriental Magpies, and also wing length in male and female Oriental Tits, and bill length and tarsus length in female Oriental Magpies were best predicted by, respectively, Annual mean temperature (bio1) and mean temperature of coldest quarter (bio11), supporting the conventional heat conservation hypothesis and demonstrating that the morphological measurements reflect selection for heat conservation in the winter rather than for heat dissipation in the summer.
Supplementary information
Supplementary informationaccompanies this paper at https://doi.org/10.1186/s4065 7-019-0172-7.
Additional file 1: Table S1.Sampling information of the Oriental Magpie(Pica serica).Additional file 2: Table S2.Sampling information of the Oriental Tit(Parus minor).
Additional file 3: Table S3.AIC and t values of the linear mixed-effect models for the relationships of morphological measurements with each climate variable.
Additional file 4: Table S4.Pearson’s correlation coefficient (r) between climate variables.
Acknowledgements
Special thanks to Peng He from the National Zoological Museum of China for providing locality and other information on specimens. We also thank the assistance of two anonymous reviewers for useful comments on our manuscript.
Authors’ contributions
LF and FL conceived the research project, LF collected the data, LF and YX analyzed the data, and LF, TC and GS led the writing. All authors read and approved the final manuscript.
Funding
This study was funded by Strategic Priority Research Program of the Chinese Academy of Sciences (XDA19050202), the National Natural Science Foundation of China (NSFC 31672299) and Collaborative Innovation Center for
Research and Development of Tibetan Agricultural and Animal Husbandry Resources.
Ethics approval and consent to participate
The experiments comply with the current laws of China.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Author details
1Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China.2College of Life Science, University of Chinese Academy of Sciences, Beijing 100049, China.
3Key Laboratory of Forest Ecology in Tibet Plateau of Ministry of Education,Institute of Tibet Plateau Ecology, Tibet Agriculture & Animal Husbandry University, Nyingchi 860000, China.4Center for Excellence in Animal Evolution and Genetics, Chinese Academy of Sciences, Kunming 650223, China.
Received: 28 April 2019 Accepted: 18 August 2019
杂志排行

Avian Research的其它文章
- Avian influenza virus surveillance in migratory birds in Egypt revealed a novel reassortant H6N2 subtype
- Optimal analysis conditions for sperm motility parameters with a CASA system in a passerine bird, Passer montanus
- The role of temperature as a driver of metabolic flexibility in the Red-billed Leiothrix (Leiothrix lutea)
- Differential cell stress responses to food availability by the nestlings of Asian Short-toed Lark (Calandrella cheleensis)
- Optimal diet strategy of a large‑bodied psittacine: food resource abundance and nutritional content enable facultative dietary specialization by the Military Macaw
- Exotic parrots breeding in urban tree cavities: nesting requirements, geographic distribution, and potential impacts on cavity nesting birds in southeast Florida