Differential cell stress responses to food availability by the nestlings of Asian Short-toed Lark (Calandrella cheleensis)
2020-01-11LiangZhangLidanZhaoXinjieZhangWeiLiangandShupingZhang
Liang Zhang, Lidan Zhao, Xinjie Zhang, Wei Liang and Shuping Zhang*
Abstract
Keywords: Asian Short-toed Lark nestling, Cell stress, Food availability, HSP70, HSP90
Background
In temperate zone, passerine birds time their breeding to track the availability of their food (Lack 1968), because birds have to raise their young at times with highest food abundance to achieve the highest reproductive output(Nager and van Noordwijk 1995; Dias and Blondel 1996).Many studies have found higher food availability results in increased growth rates (Keller and van Noordwijk 1993), and larger fledgling mass improves offspring survival (Gebhardt-Henrich and van Noordwijk 1991; Verboven and Visser 1998). The underlying mechanism of this relationship between food availability and offspring fitness remains largely unexplored. Some studies have explained this relationship from the point of malnutrition. It has been found in some passerine bird species such as Great Tit (Parus major) that the optimal food(e.g. Lepidoptera) has higher quality nutritional profile than the alternative food (e.g. Diptera and Coleoptera) (Arnold et al. 2010). Some studies found plasma metabolite level can predict changes in the individual body condition, growth rates and survival rate of nestlings (Nadolski et al. 2006; Amat et al. 2007; Albano et al.2011). However, there is little work to reveal the physiological responses of the nestlings hatched asynchronized with the food abundance peak, which is important for understanding the causal link between the timing of breeding and the fitness of offspring.
Food shortage is a major environmental factor inducing stress of wild animals (Merino et al. 2002; Herring et al. 2011). Understanding the stress status of the nestlings hatched in different food conditions is important to reveal the biological mechanism linking the food availability and fitness of offspring. Cells are the fundamental units of organisms, and the survival of the whole organism depends on the ability of cells to maintain homeostasis. The cellular stress response, an important molecular mechanism involved in maintaining cellular homeostasis, can offset the effects of stress and thereby allow other processes to restore cellular homeostasis (Kultz 2005; Somero 2010). Heat shock proteins (HSPs) are molecular chaperones that help maintain regular cellular functions by playing a crucial role in protein folding, unfolding, aggregation, degradation and transport (Beissinger and Buchner 1998;Sørensen et al. 2003; Arya et al. 2007). HSPs increase in response to acute and chronic stress, injury, and disease(Santoro 2000; Al-Aqil et al. 2013; Zulkifli et al. 2014).In particular, HSP60, 70 and 90 have been shown to be indicators of stress in several avian species, including Barn Swallows (Hirundo rustica), Broiler Chickens,and House Finches (Haemorhous mexicanus) (Merino et al. 2002; Beloor et al. 2010; Hill et al. 2013; Wein et al. 2016). Therefore, the levels of HSPs can be used as markers to reflect physiological status of the nestlings hatched under different food condition.
Asian Short-toed Lark (Calandrella cheleensis) is the most common passerine species living on the grasslands of northeastern Inner Mongolia of China (Tian et al. 2015; Zhang et al. 2017). The breeding of the species begins in mid-April and ends in mid-June, and the grasshopper nymph (Orthoptera) is the main component (about 80%) of the nestlings’ diet (Tian et al. 2015).Because there is only one significant nymph abundance peak once a year in the study area (Chen and Gong 2005;Guo et al. 2009), and the hatching time of nestlings has to be synchronized with the time of nymph abundance peak. According to our previous study, the species displays significantly within year individual variation in the timing of breeding behavior (Zhang et al. 2017; Zhao et al. 2017). Although there was an obvious hatching peak in one breeding season, there were some nestlings hatched outside this peak (Zhang et al. 2017). In addition,the synchronization between the nestling hatching peak and nymph abundance peak varied with the spring temperature. In 2014 and 2016, the peak of nestling hatching was asynchronized with the nymph abundance peak for the extremely high and low spring temperature, and nestling’s food shortage induced by such mismatch could finally reduce the survival rate of nestlings (Zhang et al.2017). Therefore, this species is an appropriate species to study the effect of food availability on the physiological status of nestlings.
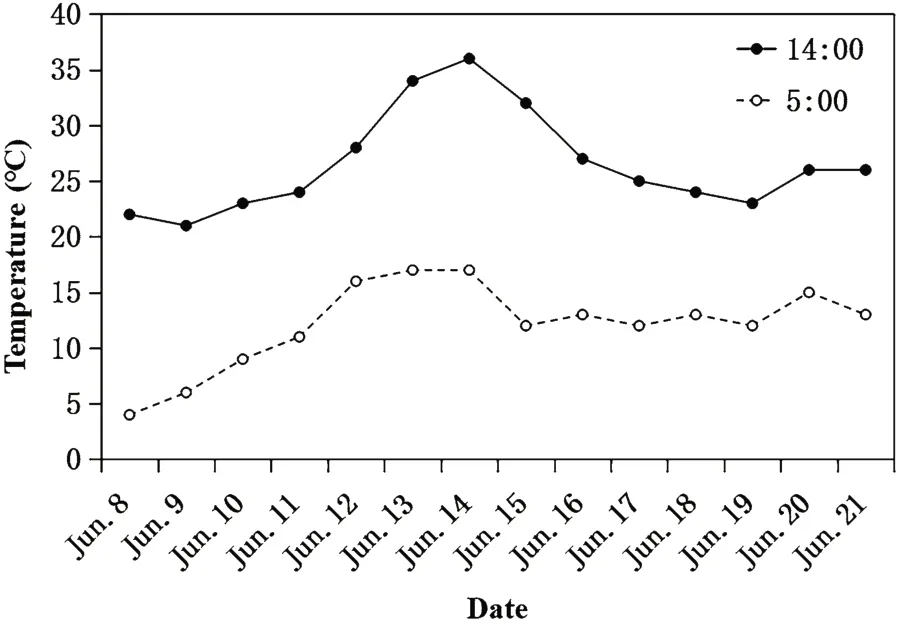
Fig. 1 Temperature at 5:00 and 14:00 of the days sampling the blood of Asian Short-toed Lark nestlings
In this paper, we hypothesize that the nestlings of Asian Short-toed Lark hatched under poor optimal food condition would activate cell stress response. To test this hypothesis, we compared the gene expression of HSP70 and HSP90 in the blood cells of Asian Short-toed Lark nestlings hatched under different nymph abundance conditions in 2017.
Methods
Study site
Our study area was in the Hulun Lake National Nature Reserve (47°45′50″-49°20′20″N, 116°50′10″-118°10′10″E),which is in the northeastern part of the Inner Mongolian Autonomous Region, China. This is a semiarid, steppe region where the mean annual temperature, precipitation and potential evaporation are - 0.6 °C, 283 mm and 1754 mm, respectively. The dominant plant species are Stipa krylovii, Leymus chinesis and Cleistogenes squarrosa. Winter is longer than summer and the approximate average maximum daytime temperatures in January and July are - 20.02 °C and 22.72 °C, respectively.
Field study design
Asian Short-toed Larks nest on the ground. From 20 April to 27 June in 2017, we surveyed nests of larks and grasshopper nymph in the study area every day. From 8 June to 21 June, we took blood samples of 64 four-day-old nestlings from different nests and recorded their hatching date. Considering the daily temperature variation(Fig. 1), each sampled nestling was blooded on 5:00 and 14:00 respectively. The relative abundance of grasshopper nymphs in the study area was quantified by catching them in an insect net on 10 parallel, 2 m × 100 m sampling transects, spaced 10 m apart, every 2 days.Captured nymphs were dried and weighed to determine their biomass. The mean daily nymph biomass was the average daily biomass obtained from all 10 transects.Annual variations in the timing of hatching and daily nymph biomass are shown in Fig. 2.
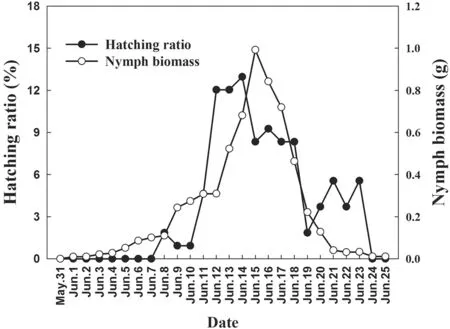
Fig. 2 Annual trends of grasshopper nymph biomass and nestling hatching ratio at Hulun lake national nature reserve in 2017
Nestling blood samples
About 100 µL of whole blood was collected from 64 fourday-old nestlings to measure HSP70 and HSP90 gene expression levels. A brachial wing vein of each nestling was punctured with a disinfected 23 G needle within 1-3 min of capture and blood that exuded from the puncture site was collected into heparinized microcapillary tubes. The skin around the puncture site was disinfected with medical alcohol before, and after, puncturing.Pressure was applied to the puncture site for 1 min with an alcohol-soaked cotton wool swab to staunch bleeding. Blood sampling procedures complied with ARRIVE guidelines and were approved by the Animal Research Ethics Committee of the Hainan Provincial Education Centre for Ecology and Environment, Hainan Normal University (permit no. HNECEE-2013-002). The study is carried out with the permissions of Hulun Lake National Nature Reserve.
RNA isolation and real-time quantitative PCR
Total RNA from the blood samples was isolated from 1 mL TRIzol reagent in 200 µL chloroform, centrifuged at 12,000 r/min for 15 min at 4 °C after which about 500µL of the supernatant was transferred to a clean tube,adding 500 µL isopropanol to the tube and centrifuging at 12,000 r/min for 15 min at 4 °C. The resultant RNA pellet was washed with 1 mL 75% ethanol (dissolved in DEPC water) two times. The RNA pellet was briefly airdried for 5 min and dissolved in 30 µL DEPC water. Total RNA quality and quantity were evaluated using agarose gel electrophoresis (AGE) and a NanoDrop 2000 spectrophotometer, and 2 µg total RNA sample was reversely transcribed to cDNA using M-MLV Reverse Transcriptase with oligo dT primers. The cDNA was used as a template in RT-qPCR. Primers were designed based on HSP70 and HSP90 mRNA sequences in NCBI using Primer3 software (Table 1). The procedures for RT-qPCR were as follows: pre-degeneration at 95 °C for 1 min,degeneration at 95 °C for 15 s, annealing at 59 °C for 15 s and extension at 72 °C for 40 s, totally 40 cycles. After the reaction had completed, a melting curve analysis from 55 to 95 °C was applied to ensure consistency and specificity of the amplified product. The expression of β-actin was confirmed to be stable under all treatments and this gene was consequently used as the reference gene to normalize mRNA levels among samples. RT-qPCR was performed twice, in triplicate. The values of the average cycle threshold (Ct) were determined and Ct scores for gene transcripts in each sample were normalized using the ΔCt scores for β-actin and expressed as the fold change in gene expression using the equation 2-ΔΔCT.

Table 1 Primer sequences and anticipated size of the amplified products
Statistical analysis
We used “nymph biomass proportion (NBP)”, the percentage of the daily nymph biomass relative to the total biomass measured over all survey days, as a measure of grasshopper nymph abundance in the statistical analysis. The effects of nymph biomass, sample time and their interaction on the blood HSP70 and HSP90 gene expression levels in nestlings are analyzed with a linear mixed model (LMM) in which the individual nestling is a random factor. Regression analysis is used to analyze the relationship between nymph biomass and the gene expression level of HSP70 and HSP90 if the effect of nymph biomass is significant in the LMMs. We used α = 0.05 as the significant level for all statistical tests. All analyses were performed using SPSS version 24 (SPSS Inc., Chicago, USA).

Table 2 Results of linear mixed models (LMMs) on the effects of nymph biomass, sample time and the interactions of the two factors on gene expression levels of blood HSP70 and HSP90 in nestlings of wild Asian Short-toed Lark in the Hulun Lake NatureReserve, the Inner Mongolian Autonomous Region, China
Results
The results of the LMMs for the effects of nymph biomass and sample time on gene expression level of HSP70 and HSP90 indicate that nymph biomass, sample time and interaction of the two factors significantly influenced the blood HSP70 and HSP90 gene expression level of Asian Short-toed Lark nestlings (Table 2). According to this result, the relationships between nymph biomass and HSP70 and HSP90 gene expression level at 5:00 and 14:00 were analyzed separately. Simple linear regression analysis showed that the nymph biomass negatively correlated with the gene expression level of HSP70 (5:00, n = 64,R2= 0.811, Fig. 3a; 14:00, n = 64, R2= 0.636, Fig. 3c) and HSP90 (5:00, n = 64, R2= 0.666, Fig. 3b; 14:00, n = 64,R2= 0.743, Fig. 3d). The gene expression levels of HSP70 and HSP90 increase with the decrease of nymph biomass(Fig. 3). In addition, both HSP70 and HSP90 gene expression levels at 14:00 are significantly higher than those at 5:00 (Fig. 3).
Discussion
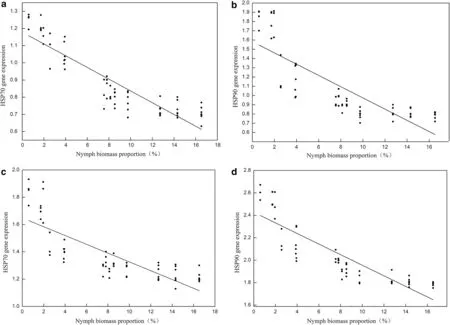
Fig. 3 The simple liner regression between nymph biomass proportion (NBP) and gene expression of HSP70 (a: 5:00, n = 64, R2 = 0.811; c: 14:00,n = 64, R2 = 0.636) and HSP90 (b: 5:00, n = 64, R2 = 0.666; d: 14:00, n = 64, R2 = 0.743) in the blood of Asian Short-toed Lark (Calandrella cheleensis)
The LMM results indicate that the nymph abundance is a factor which significantly influences the gene expressions of HSP70 and HSP90 of Asian Short-toed Lark nestlings.Negative correlation between nymph biomass and gene expressions of HSP70 and HSP90 suggests that poor food condition is an important environment factor inducing cell stress of Asian Short-toed Lark nestlings. Similar results about the relationship between food availability and stress protein level of nestlings have been found in other species such as White Ibis which showed that the HSP60 levels increased predictably in response to food limitation (Herring and Gawlik 2013). Our results and the available study jointly show that food shortage during post embryo development can induce the stress in nestlings. Several studies have compared the response of HSP60, HSP70 and HSP90 to the ecological factors.While some of the studies found HSP60 is more sensitive than other HSPs (Martínez-Padilla et al. 2004; Moreno et al. 2005), other studies found HSP70 was more sensitive to environmental perturbations than HSP60 or HSP90 (Ulmasov et al. 1992; Fader et al. 1994; McMillan et al. 2011). Our study showed that HSP70 and HSP90 of nestlings respond to the optimal food condition similarly.The different responses of the HSPs might be species specific, therefore we suggest that multiple HSPs should be used as cell stress markers in the evaluation of cell stress status.
In addition to the food availability, the LMM results of our study also indicate the sample time is also an important factor inducing the variation of HSPs level. HSP70 and HSP90 gene expressions in the blood sampled at 14:00 were significantly higher than those sampled at 5:00, which could be explained by the daily temperature variation in the habitat. Our study area was in the northeastern part of the Inner Mongolian Autonomous Region, where the daily temperature variation between minimum and maximum is 10-15 °C (Chen and Gong 2005). The temperature at 14:00 was higher than that at 5:00 (Fig. 1). Temperature variation has been found to be an important environment factor inducing the HSPs expression (Arya et al. 2007). Therefore, the expression level of HSPs at 14:00 increasing in our study indicates the nestlings could cope with the temperature variation to maintain homeostasis by cell stress response.The response of blood HSPs to the marked temperature variation in the study area has been found in our previous study of adult Asian Short-toed Larks (Qin et al.2017). These results indicate that cell stress response is an important physiological mechanism for birds to cope with highly stochastic ecosystems not only in adult but also in nestlings. Our LMM model suggested that poor food condition enhanced the level of cell stress of nestlings at both low and high temperature.
Although the HSPs can maintain the homeostasis of the cells, overexpression of these stress proteins is known to have deleterious consequences (Feder and Hofmann 1999). Their synthesis represents a significant energetic cost (Hamdoun et al. 2003), which can lead to reduced fecundity (Parsons 1996). In addition, their response usually results in a concomitant reduction in the synthesis of antibodies, thus synthesizing more HSPs to mitigate stress has been found in passerine birds to be traded-offagainst mounting humoral and cell-mediated immune responses (Morales et al. 2006). The result of our previous study on the Asian Short-toed Lark showed that the survival rate of the nestlings in 2014 and 2016 when the peak of nestling hatching mismatched with the peak of nymph abundance peak was significantly lower than that in 2015 when the two peaks thoroughly match (Zhang et al. 2017), which suggests that overexpression of HSPs could be related with lower nestling survival. As a bird species living in the harsh weather condition, the nestlings of Asian Short-toed Lark have to cope with the marked weather variation by cell stress response, and poor food condition could impose additional stress to make HSPs overexpression, which consequently could result in negative effect on the nestling survival. Therefore, over cell stress response may be one of physiological factor mediating the effect of optimal food availability and the nestling’s survival.
Shifts in phenology induced by climate change are altering ecological relationships and processes around the world (Visser and Both 2005; Cleland et al. 2007;Forrest and Miller-Rushing 2010) and there is a growing body of literature on resultant trophic mismatches between nestlings and their food resources (Visser et al.1998; Visser and Both 2005; Thackeray et al. 2010).Although such trophic mismatches have been found to result in nestlings having lower body mass, smaller body size and reduced survival rates (Verboven and Visser 1998; Visser et al. 1998; Durant et al. 2007; Ovaskainen et al. 2013; Dunn and Møller 2014), there is little information to reveal the physiological mechanism how the optimal food condition influences the fitness of the nestlings. Our research indicates that cell stress biomarkers have the potential to respond directly and predictably to optimal food conditions. Under the phenological mismatch scenario, the birds living in the fluctuating environment would encounter more stress challenges, which implicates that these birds could be more vulnerable to the phenological mismatch. Therefore, we suggest the vulnerability of the species to the phenological mismatch induced by climate change should be estimated combining the physiological and ecological data.
Conclusion
The results of this study indicate that poor food condition is an important environment factor inducing cellular stress of Asian Short-toed Lark nestlings. The interactive effect of the nymph abundance and sample time on the HSPs response may be related with the daily temperature variation of the grassland. Over cell stress response may be one of physiological factor mediating the effect of food availability and the nestling’s fitness.
Acknowledgements
We are grateful to Manquan Gui, Muren Wu, Songtao Liu in Hulun Lake National Nature Reserve for their help on the field study.
Authors’ contributions
SZ and WL conceived the study and designed the experiments. L. Zhaang,L. Zhao and XZ conducted the experiments. LZ wrote the first draft of the article. SZ supervised the research and revised the draft. All authors read and approved the final manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (No. 31872246).
Availability of data and materials
The data used in the present study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
Our experimental procedures complied with the current laws on animal welfare and research in China and had the approval of the Animal Research Ethics Committee of Hainan Normal University. In addition, all procedures followed standard protocols, such as the ARRIVE guidelines for reporting animal research.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Author details
1College of Life and Environment Sciences, Minzu University of China, Beijing 100081, China.2Ministry of Education Key Laboratory for Ecology of Tropical Islands, College of Life Sciences, Hainan Normal University, Haikou 571158,China.
Received: 25 February 2019 Accepted: 17 October 2019
杂志排行

Avian Research的其它文章
- Avian influenza virus surveillance in migratory birds in Egypt revealed a novel reassortant H6N2 subtype
- Optimal analysis conditions for sperm motility parameters with a CASA system in a passerine bird, Passer montanus
- The role of temperature as a driver of metabolic flexibility in the Red-billed Leiothrix (Leiothrix lutea)
- Optimal diet strategy of a large‑bodied psittacine: food resource abundance and nutritional content enable facultative dietary specialization by the Military Macaw
- Exotic parrots breeding in urban tree cavities: nesting requirements, geographic distribution, and potential impacts on cavity nesting birds in southeast Florida
- Drivers of nest survival rate in a southern Tunisian population of Laughing Doves(Spilopelia senegalensis)