黄海獐子岛几种大型红藻的分类研究❋
2020-01-09王艺晓陈彦伟丁兰平刘金梅姜晶晶黄冰心
王艺晓, 陈彦伟, 杨 楠, 丁兰平, 刘金梅, 姜晶晶, 黄冰心
(天津师范大学生命科学学院, 天津市动植物抗性重点实验室, 天津 300387)
大连獐子岛地处太平洋与亚欧大陆间的中纬地带,素有“黄海明珠”之美誉。岛上拥有丰富的藻类资源,得天独厚的地理位置为海藻的生长提供了适宜的场所。
红藻是重要的海洋大型藻类类群,具有很高的研究和利用价值。其中,杉藻目海藻富含卡拉胶或琼胶[1],可用于食品、药品制造和工农业生产中[2]。角叉菜属(Chondrus)和马泽藻属(Mazzaella)均属于杉藻目、杉藻科。蜈蚣藻属(Grateloupia)隶属于红藻门、隐丝藻目、海膜科[3]。蜈蚣藻可食用,并因其色泽光鲜,被用于合成天然色素,可作为生产琼胶的原料[4],也可用于制作清热解毒的药物,其中繁枝蜈蚣藻(Grateloupiaramossissima) 和带形蜈蚣藻(Grateloupiaturuturu)还具有抗病毒活性[5-6]。
角叉菜属由Stackhouse于1797年建立[7],种间差异比较小[8]。目前,角叉菜属已报道104种及65个种下单位,其中仅19种得到确认,主要分布于北太平洋和北大西洋[9],中国已发现10种,即扩大角叉菜(Chondrusarmatus(Harvey) Okamura),沟状角叉菜(Chondruscanaliculatus(C.Agardh) Greville),皱叶角叉菜(ChondruscrispusStackhouse),皱叶角叉菜纤细变种(Chondruscrispusvar.filiformis(Hudson) Lyngbye,Chondruscrispusvar.lonchophorusMontagne),日本角叉菜(ChondrusnipponicusYendo),角叉菜(ChondrusocellatusHolmes),树枝角叉菜(Chondruspinnulatus(Harvey) Okamura),宽叶角叉菜(Chondrusplatynus(C.Agardh) Ruprecht)和异色角叉菜(ChondrusverrucosaMikami[10])。1904年,Gepp[11]报道了产自山东威海的(Cystocloniumarmatus(C.armatus))。而后,曾呈奎、杭金欣、周贞英等[12-14]先后报道了产自广东、福建和浙江的C.ocellatus。1998年,栾日孝[7]等报道了产自大连的C.armatus和C.yendoi(楔形角叉菜)。1999年,夏邦美[15]报道了产自山东的C.nipponicus,并指出曾呈奎、杭金欣等[12-13]报道的产于东南沿海的Chondrusocellatus不属于角叉菜属。2001年,苏乔、栾日孝等[16]报道了产自大连的C.ocellatus和Mazzaellajaponica(日本马泽藻)。2002年苏乔、栾日孝等[17]指出产于中国东南沿海的角叉菜C.ocellatus应为Grateloupiaimbricata(复瓦蜈蚣藻),最初被归类在角叉菜属的C.yendoi应为Mazzaellajaponica。其中,由于角叉菜(C.ocellatus)的形态变异很大,Mikami[18]在前人研究的基础上将其划分为4个变型,即C.ocellatusf.aequalis、f.crispoides、f.ocellatus和f.parvus。中国该属的物种虽然不多,但以往的鉴定是否正确,学者们仍然存有疑虑[17]。
马泽藻属由De Toni于1936年建立,目前,本属已报道31种及4个种下单位,28种得到确认[9]。中国仅发现3种,即报道产于台湾的近源马泽藻(Mazzaellaaffinis(Harvey) Fredericq),报道产于台湾的异果马泽藻(Mazzaellaheterocarpa(Postels et Ruprecht) Fredericq)和报道产于辽宁的日本马泽藻(Mazzaellajaponica(Mikami) Hommersand[10])。其藻体主要特征为叉状分枝、表面有光泽且平滑。本属在中国黄、渤海沿岸有分布,多生长于中、低潮带的岩石上[19]。近年来,在生长季节,本属海藻在中国北部沿海的生长呈爆发趋势,但涉及物种的相关报道非常少。
蜈蚣藻属是海膜科里最大的属,被认为是世界上最难区分的属之一[20],目前,本属已报道149种及30个种下单位,96种得到确认[9]。蜈蚣藻属的分类鉴定是以藻体形态、内部结构、繁殖器官等特征为基础,但藻体形态通常受环境影响而发生改变[21]。近年来,王宏伟教授[33,37]等专家学者对中国蜈蚣藻属开展了丰富的研究工作,发表了一系列研究成果,本文对此不再重复相关内容。
在以上研究的基础上, 本文对大连獐子岛角叉菜属、马泽藻属和蜈蚣藻属海藻进行了分类研究。该研究结果丰富了该海域海藻的物种多样性,为獐子岛地区角叉菜属、马泽藻属和蜈蚣藻属物种资源的利用提供分类学依据。
1 材料和方法
1.1 材料
以2017年8月3—6日采集自大连獐子岛的角叉菜属、马泽藻属和蜈蚣藻属标本为实验材料,包括:浸液(用5%甲醛海水液和90%酒精液浸泡)和腊叶标本,编号保存。
1.2 方法
1.2.1 外形特征比较 主要观察藻体的轮廓、高度、宽度、颜色、主轴及分枝特点、藻体顶端及基部特点、以及生殖器官的形状、大小、发生位置等。
1.2.2 显微特征比较 将样品制成临时装片,放置于荧光显微镜Leica DM5000B和体式显微镜Nikon SMZ25下观察并拍照。观察、比较藻体皮层厚度;内外皮层细胞层数、大小、形状;髓部是否中空、髓部细胞形状、大小等。
1.2.3 物种鉴定 结合相关文献和数据,与上述特征进行比对,鉴定物种。
2 结果
2.1 角叉菜属
2.1.1 角叉菜(ChondrusocellatusHolmes) 由Holmes[22]首次命名(见图1)。

(a.藻体外观;b.楔形基部;c.分枝小育枝;d.顶端分枝;e.分枝顶端;f.皮层细胞;g~i.藻体纵切面观,髓丝。 a.External morphology of a frond; b.Cuneate base; c.Branches give birth to young branches; d.Top branches; e.Top of the branches; f.Cortical cells; g~i.Longitudinal section view of frond, medullary filaments.)
图1 角叉菜的形态学特征
Fig.1 Morphological characteristicsChondrusocellatus
特征描述:藻体红褐色,偶尔带绿,丛生,高8.2 cm。基部扁平楔形,具亚圆柱状短柄。多次叉状分枝呈扇形,枝端微尖或钝圆,主枝和分枝上生有副枝和小育枝。藻体内部结构分为皮层和髓部,皮层厚80~100 μm,皮层细胞向内逐渐变大。外皮层细胞椭圆或近圆形,5~8层,最外层细胞长2.5~4 μm,宽约2 μm,含色素体,向内细胞长4~6 μm,宽2~5 μm。内皮层细胞近圆形或星形,2~4层,长6~8 μm,宽 4~6 μm。髓部由丝状细胞构成,呈网状排列,多与藻体表面平行,长25~78 μm,宽1~5 μm。
习性: 固着于中、低潮带石沼中的岩石上。北太平洋特有物种。
模式标本产地:日本。
标本编号:20180804004A,采集自大连獐子岛褡裢岛。
2.1.2 角叉菜小型变型(C.ocellatusf.parusMikami) 由Mikami[18]首次命名(见图2)。
特征描述:藻体紫红色,高2.4 cm。基部楔形,固着器不规则圆盘状,2~3次二叉状分枝,腋角钝圆,分枝顶端不规则浅裂。藻体内部结构分为皮层和髓部,皮层细胞由外向内逐渐变大,外皮层细胞椭圆形,长2~7 μm,宽1~3 μm,约6层,排列紧密。内皮层细胞近圆形或不规则形,长2~7 μm,宽1~3 μm,2~3层。髓部由髓丝组成,多数与表面平行。孢子囊数目众多,分布于整个藻体上,常彼此相互汇合在一起。孢子囊成熟时呈椭圆形,长30~60 μm,宽25~40 μm,十字形分裂。
习性:大多在潮间带及石沼内生长,本变型在辽东半岛的分布不普遍,量少。分布于东亚地区。
模式标本产地:日本。
标本编号:20170805001B,采集自大连獐子岛大圈。

(a.藻体外观;b.基部固着器;c.叉状分枝;d~e.分枝顶端;f~g.藻体纵切面观;h~i.髓丝和近一步发育的孢子囊。a.External morphology of a frond; b.Base fixator; c.Dichotomous branches; d~e.Top of the branches; f~g.Longitudinal section view of frond; h~i.Medullary filaments and developed sporangiums.)
图2 角叉菜小型变型的形态学特征
Fig.2 Morphological characteristics ofC.ocellatusf. parus Mikami
2.2 马泽藻属
2.2.1 日本马泽藻(Mazzaellajaponica) 由Hommersand[23]首次命名(见图2)。
特征描述:藻体紫红色,表面光滑,具荧光,叶片状,高5.2 cm。盘状固着器,藻体下部具楔形短柄,分枝不规则,枝宽1~6 cm。藻体内部结构由皮层和髓部组成,皮层厚50~80 μm,细胞由外向内逐渐变大,6~8层,含色素体。外皮层细胞近椭圆形,长3~10 μm,宽2~4 μm,排列紧密。内皮层细胞近圆形或星形,长8~13 μm,宽8~10 μm,约2层。髓部由髓丝组成,多与表面平行,宽约3 μm。
习性:常生长于潮间带岩石沼泽,部分也可在低于低潮线的岩石中生长。东亚特有种类。
模式标本产地:日本。
样品编号:20170805006A,采集自大连獐子岛大道沟。

(a.藻体外观;b~c.分枝顶端;d.叶柄;e~f.藻体纵切面观;g.髓丝;h~i.皮层细胞。a.External morphology of a frond; b~c.Top of the branches; d.Petiole; e~f.Longitudinal section view of frond; g.Medullary filaments; h~i.Cortical cells.)
图3 日本马泽藻的形态学特征
Fig.3 Morphological characteristics ofMazzaellajaponica
2.3 蜈蚣藻属
2.3.1 亚洲蜈蚣藻(Grateloupiaasiatica) 由Kawguchi[33]首次命名(见图4、5)。
特征描述:藻体紫红色,扁压状,较小个体高5.5 cm,基部具圆柱形短柄,主枝1~2回羽状分枝,主枝顶端为两叉分枝,各叶片末端一般尖细。较大个体高9.5 cm,主枝直立,线形,基部固着器不规则圆盘状,羽状分枝,枝端尖细。藻体内部结构分皮层和髓部,皮层细胞由外向内逐渐变大,5~7层,厚50~70 μm。外皮层细胞近椭圆形或近圆形,3~5层,长3~8 μm,宽2~5 μm。内皮层细胞近椭圆形或多角形,2~3层,长9~17 μm,宽5~10 μm。髓部髓丝纵行分布,宽3~9 μm。
习性:生长在潮间带石沼中,秋冬季节成熟。分布于东亚、东南亚及欧洲的法国。
模式标本产地:日本。
样品编号:20170806008(较小个体),采集自大连獐子岛褡裢岛;20170804008(较大个体),采集自大连獐子岛二大滩。

(a.藻体外观;b.主枝顶端;c.指形小枝;d~e.藻体基部横切面观;f~h.皮层细胞;i.基部横切面髓丝。a.External morphology of a frond; b.Top of the main branches; c.Finger twigs;d~e.Transverse section view of frond base; f~h.Cortical cells; i.Medullary filaments in transverse section at base.)
图4 亚洲蜈蚣藻(20170806008)的形态学特征
Fig.4 Morphological characteristics ofGrateloupiaasiatica(20170806008)

(a.藻体外观;b.基部固着器;c.分枝;d.主枝顶部;e.藻体基部横切面观;f~h.皮层细胞;i.基部横切面示髓丝。a.External morphology of a frond; b.Base fixator; c.Branches; d.Top of main branches; e.Transverse section view of frond base; f~h.Cortical cells; i.Medullary filaments in transverse section at base.)
图5 亚洲蜈蚣藻(20170804008)的形态学特征
Fig.5 Morphological characteristics ofGrateloupiaasiatica(20170804008)
2.3.2 链状蜈蚣藻(Grateloupiacatenata) 由Yendo[25]首次命名(见图6)。
特征描述:藻体红棕色,高11 cm,主枝线状,末端尖细,不规则羽状分枝,两侧有长短不同的小枝,顶端尖细或生出更小的分枝,基部隘缩。藻体髓部中空,外皮层5层细胞,最外一层细胞近椭圆形,长10~12.5 μm,宽2.5~4 μm,呈栅栏状整齐排列,向内细胞近圆形或不规则形,逐渐变大,直径5~17 μm;内皮层为短圆柱形细胞,底面直径约3 μm,多垂直于横切面。
习性:藻体生长于中、高潮带的岩石上,5—8月为生长发育期。东亚特有种类。
模式标本产地:日本。
样品编号:20170805012B,采集自大连獐子岛大圈。
2.3.3 青岛蜈蚣藻(Grateloupiaqingdaonsis) 由夏邦美[35]首次命名(见图7)。
特征描述:藻体紫红色,高13 cm,丛生。主枝直立,扁形,末端叉状分枝,2~3回羽状分枝,羽枝长,对生、互生或偏生,排列紧密,向上弯曲如镰刀状,羽枝上生有小育枝,分枝末端尖细或两叉分枝。藻体内部结构分皮层和髓部。皮层细胞8~20层,厚80~150 μm。外皮层细胞4~18层,近椭圆形,长5~10 μm,宽3.5~5 μm。内皮层细胞2~3层,星形、近圆形或不规则形,长5~15 μm,宽4~11 μm。髓部髓丝交错分布,长9~67 μm,宽3~5 μm。
习性:生长在低、中潮带的岩石或沙砾上。
模式标本产地:主模式标本为LI83-446,雌配子体,系1983年7月14日李伟新采自中国山东省青岛。
标本编号:20170804015A,采集自大连獐子岛东邦。
2.3.4 带形蜈蚣藻(Grateloupiaturuturu) 由Yamada[26]首次命名(见图8)。
特征描述:藻体淡红色,带淡黄色,黏滑,扁平带状,高18 cm,宽5~6.5 cm,基部楔形,有短柄,固着器圆盘状,藻体边缘波浪状。藻体由皮层和髓部组成。皮层细胞由外向内逐渐变大,6~7层,厚50~60 μm。外皮层细胞3层,椭圆形,长5~10 μm,宽3~5 μm;内皮层细胞2~3层,椭圆形,长6~10 μm,宽5~8 μm;中央髓丝垂直于表面分布,宽2~5 μm。
习性:通常在中、高潮带的岩石上生长,6—8月为成熟期。东亚、美洲、非洲、澳洲和欧洲均有分布。
模式标本产地:日本。
标本编号:20170806017,采集自大连獐子岛褡裢岛。
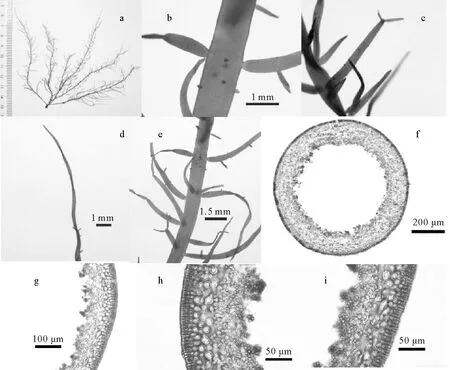
(a.藻体外观;b.小枝基部;c.小枝顶端;d.主枝末端;e.主枝分枝;f.藻体横切面观;g~i.皮层细胞。a.External morphology of a frond; b.Branchlets base; c.Top of branchlets; d.End of branchlets; e.Branch of main branches; f.Transverse section view of frond; g~i.Cortical cell.)
图6 链状蜈蚣藻的形态学特征
Fig.6 Morphological characteristicsGrateloupiacatenata

(a.藻体外观;b.基部;c.小育枝;d.部分分枝末端;e.主枝末端;f.藻体横切面观;g~h.皮层细胞;i.髓丝。a.External morphology of a frond; b.Base; c.Young branches; d.End of partial branches; f.Transverse section view of frond; g~h.Cortical cells; i.Medullary filaments.)
图7 青岛蜈蚣藻的形态学特征
Fig.7 Morphological characteristicsGrateloupiaqingdaonsis

(a.藻体外观;b~c.基部;d~e.藻体边缘;f~h.藻体纵切面观。a.External morphology of a frond; b~c.Base; d~e.Margin of frond; f~h.Longitudinal section view of frond.)
图8 带形蜈蚣藻的形态学特征
Fig.8 Morphological characteristicsGrateloupiaturuturu
3 讨论
角叉菜属在辽东半岛集中分布在大连的各个海区,其它的海区极少。由于其生境差异明显,即使是同一种变型,其形态差异也很大,但它们都有相似的内部结构:皮层细胞由外向内逐渐变大,外皮层细胞5~7层,内皮层细胞2~3层;髓部由网状、稍与表面平行的髓丝组成;四分孢子囊由髓细胞发育而成。目前在中国已发现的3种角叉菜(角叉菜、日本角叉菜和扩大角叉菜)的区别在于:(1)日本角叉菜和扩大角叉菜分枝方式是二叉状分枝或双二叉状分枝,而角叉菜只有二叉状分枝;(2)日本角叉菜和扩大角叉菜的高(10~15 cm),纵切面细胞的宽(7~12 μm)和长(7~15 μm)都比角叉菜的大(对应分别是4~10 cm,4~5.5 μm,5~6 μm)[8]。上述关于角叉菜高度和细胞大小的报道与本文中角叉菜的相关描述基本吻合。此外,角叉菜的另一特征——囊果,在藻体各部分都可能生长(基部除外),一面凹陷,一面凸出[36]。但本次采集的角叉菜未发现囊果。王宏伟等[27]在2014年对大连水域中采集的日本角叉菜的雌配子进行鉴定,发现囊果出现在9月—次年3月;10月中旬—11月中旬大连海水平均温度分别为17.8和13.2 ℃,此间雌配子体上成熟囊果明显多于其他月份。根据这一研究结果推测,本次未能采集到带有囊果的角叉菜可能和采集的月份(本次采集时间为8月上旬)和水温有关。
角叉菜小型变型由Mikami[18]建立,中国关于角叉菜小型变型的报道很少,仅李熙宜等[37]描述其藻体矮小、2~3 cm、扇形、1~3次近双叉分枝和四分孢子囊发育情况与角叉菜原变形相同。本样品特征基本与Mikami[18]和李熙宜[28]等的相符。在中国北部沿海,角叉菜的外形变化很大,过去都归入本种中,引起了一些争议,可能存在如Mikami介绍的不同变型或者属于不同的物种,希望后续能结合分子分析进一步明确。
马泽藻属和角叉菜属亲缘关系很近[38],日本马泽藻分枝无规则,而角叉菜属分枝方式则为有规律的叉状分枝,且日本马泽藻表面具荧光、叶片状。这些外形上的特征可以帮助区别马泽藻和角叉菜属海藻。本文日本马泽藻的内部结构与张晓明[30]等的基本相符,例如皮层厚50~80 μm,细胞5~7层(本文为6~8层),外皮层细胞椭圆形,长2~10 μm,宽2~3 μm,(本文为长3~10 μm,宽2~4 μm),内皮层细胞直径8~15 μm(本文为长8~13 μm,宽8~10 μm)等。
过去,报道产于亚洲的蜈蚣藻(G.filicina)是一个很常见的种类,但2001年王宏伟等[29]认为是一新种—亚洲蜈蚣藻(G.asiaticaKawaguchi et Wang),并确认G.filicina在西太平洋和亚洲没有分布[43]。本次采集到的个体较大的亚洲蜈蚣藻样品(20170804008),与文献[35]所报道的蜈蚣藻(G.filicina)的外形与内部特征相似,差异仅为内皮层细胞大小不同,该文献报道的内皮层细胞长15~20 μm、宽20~25 μm,而本文样品的长9~17 μm、宽5~10 μm。本次采集的个体较小的(20170806008)亚洲蜈蚣藻与两叉蜈蚣藻(G.dichodoma)的特征相似。但戚贵成等(2009)[20]认为两叉蜈蚣藻在辽宁沿海并无分布,过去报道的应该并入亚洲蜈蚣藻,二者区别在于亚洲蜈蚣藻主枝 1~2回或 2~3 回羽状分枝、分枝末端尖细,两叉蜈蚣藻主枝叉羽状或叉状分枝、顶端为两叉。本文大、小个体的样品符合王宏伟、戚贵成等[20,24]报道的亚洲蜈蚣藻的特征,故都鉴定为亚洲蜈蚣藻。本文对亚洲蜈蚣藻的内部结构进行了具体描述,但并未发现其生殖器官。王宏伟等[37]报道采集于4—7月的藻体没有生殖器官,8月开始出现四分孢子囊、囊果等生殖器官,且生殖器官首先出现在较大个体上。而本文样品的采集时间在8月初,亚洲蜈蚣藻的生殖器官可能仍在发育的初期,特征不明显。
依据藻体中空的特点,李伟新等[31-32]将蜈蚣藻节荚变型Grateloupiafilicinaf.lomentaria和中空变型Grateloupiafilicinaf.porracea移出蜈蚣藻属,另建一新属——管形藻属Sinotubimorpha,但WANG 等[33]对蜈蚣藻的节荚变型、中空变型和管形藻Sinotubimorphacatenata进行形态学和 rbcL 序列分析后,认为它们为同物异名,并将它们归类成蜈蚣藻属中一个独立种GrateloupiacatenataYendo(1920) (链状蜈蚣藻)。本文接受WANG 等[33]的观点,且相关样品的外部形态和内部结构特征符合李伟新等[31-32]和WANG 等[33]所描述的蜈蚣藻中空变型、管形藻和链状蜈蚣藻。
关于青岛蜈蚣藻的报道非常少,仅《中国海藻志》[35]对其进行了形态学描述。本文青岛蜈蚣藻与文献[35]描述基本相符,例如:2~3回羽状分枝,羽枝长,排列紧密,向上弯曲如镰刀状,皮层细胞8~22层(本文为8~20层)等。
本文带形蜈蚣藻样品的内部结构与文献[35]描述基本相符,差异主要在藻体大小和颜色方面,文献报道其藻体可高达40~100 cm、甚至高于100 cm,颜色鲜红、淡红或深红带绿,本文样品的藻体高度未达到40 cm且颜色淡红带黄。然而,本文带形蜈蚣藻样品的这些外部形态特征与文献[35]报道的裂叶蜈蚣藻Grateloupialatissima相似。戚贵成[20]等认为中国裂叶蜈蚣藻与新西兰的带形蜈蚣藻属于同一亚群,二者碱基差异为种内差异,并暂时命名其为带形蜈蚣藻裂叶变形Grateloupiaturuturf.latissima。本文样品的内部结构和大部分外形特征与文献[35]中对带形蜈蚣藻的描述相近,暂命名其为带形蜈蚣藻,但仍需进一步研究。
