时间序列遥感影像支持下杭州湾南岸植被动态监测
2020-01-09刘永超李加林赵冰雪许文轩何改丽刘永学
刘永超, 李加林, 张 宇, 赵冰雪, 许文轩, 何改丽, 刘永学*
时间序列遥感影像支持下杭州湾南岸植被动态监测
刘永超1,2,3, 李加林4, 张 宇5, 赵冰雪1,6, 许文轩1, 何改丽1, 刘永学1,2,3*
(1.南京大学 地理与海洋科学学院, 江苏 南京 210023; 2.自然资源部海岸带开发与保护重点实验室, 江苏 南京 210024; 3.南京大学 中国南海研究协同创新中心, 江苏 南京 210023; 4.宁波大学 东海研究院, 浙江 宁波 315211; 5.南京师范大学 地理科学学院, 江苏 南京 210023; 6.池州学院 地理与规划学院, 安徽 池州 247000)
海岸带植被是连通陆海生态系统的关键要素, 在全球变暖、海平面上升及人类活动加剧等背景下, 掌握海岸带植被演化规律是支撑未来海岸带弹性发展的基础. 以太平洋西岸中部长江三角洲杭州湾南岸为研究区, 利用时间序列Landsat遥感影像对1984至2018年杭州湾南岸植被动态进行监测. 结果表明: 杭州湾南岸植被面积从1984年的7.26km2增加至2018年的468.35 km2, 主要经历了生长发育到迅速扩张再到缓慢增长等演化阶段; 在围垦、人工引种及泥沙淤积等影响下, 海岸植被垂直于海堤沿海岸呈带状分布并伴有稀疏碎斑, 其景观构成与演替特征显著; 研究也显示了Google Earth Engine对于海岸植被长时序持续监测的优势与潜力. 研究揭示的人类活动加剧背景下杭州湾南岸植被变化特征, 对维护滨海湿地生态系统安全及大湾区可持续发展有重要意义.
时间序列分析; 植被动态; 围垦; 人工引种; 杭州湾南岸
海岸植被具有海岸防护、生物栖息、水质净化和蓝色碳汇等功能, 在全球变暖、海平面上升及人类活动加剧背景下[1], 成为偏离海陆生态系统中心横跨海陆间的狭长生态脆弱带. 1980年前后长江口人工引种的北美洲互花米草[2]由保滩促淤到侵占土著植被生境的变化, 以及海岸人工地貌逐渐增多[3-4], 严重冲击了海岸带生态系统演化[5]. 目前, 海岸带研究以岸线景观[6-8]、蓝碳[9-10]、围垦[11]、盐沼[12]、脆弱性[13-14]、海草[15]、植被光谱[16]和植被护岸功能[17]等内容为主. 随着陆海交互频繁发展, 潮汐周期性浸没以及海堤与滨海公路围栏影响的加剧[18], 野外大面积连续同步观测困难, 使传统实地调研手段难以进行. 加之海岸带地物光谱特征混淆严重[12], 单时相遥感影像难以准确表征植被分布特征[19].
卫星传感器支撑下的地理信息迅速增长, 催生了“大数据科学范式”[20], 推动了遥感技术更加高效地对地观测研究, 尤其是2010年Google、卡内基梅隆大学、美国USGS开发推出Google Earth Engine (GEE)云计算平台, 通过JavaScript API或Python离线编程方式获取和处理共享的海量数据, 可以减少影像下载和预处理等过程, 提高了研究效率[21]. 已有研究借助于GEE探讨了全球植被覆盖[22]、陆表水体[23]、水稻[24]、潮滩轨迹[25]以及湿地景观格局演变[26]等资源环境演化信息, 展现了GEE遥感地学分析的强大功能与优势.
选取太平洋西岸中部长江三角洲杭州湾南岸为研究区, 该区域以平原型淤泥质海岸为主, 海涂广阔, 岸线曲折, 资源丰富, 同时经济发达, 城镇密集, 人地交互作用剧烈, 是我国海岸带研究的典型区域. 本文基于GEE云平台, 揭示了杭州湾南岸1984年至2018年海岸植被覆盖变化特征, 丰富了GEE海岸带研究领域案例, 可为维护滨海湿地生态系统安全及大湾区可持续发展提供决策参考.
1 数据来源与研究方法
1.1 研究区概况
杭州湾位于太平洋西岸中部、长江三角洲浙江东北部区域, 西接钱塘江, 东入东海. 入海泥沙在湾内外堆积, 部分经波浪与潮汐动力随沿岸流南移, 使杭州湾南岸成为全球淤泥质河口海岸带规模最大、发育最完善的河口型潮滩之一, 并入选国际重要湿地名录. 该区域是长江三角洲平原与浙东南火山岩低山丘陵的接壤地带, 地形以平原和低山丘陵为主, 属于亚热带季风湿润气候, 年均气温16~21℃, 红黄壤相间分布, 长有草本、灌木和乔木等植被, 也是长三角城市群重要组成部分, 行政上涉及宁波市的余姚、慈溪和镇海区域(图1).

图1 研究区地理位置
1.2 数据来源及处理
在GEE中通过JavaScript API在线访问研究区行列号118/039区域, 数据包括1984至2018年Landsat 5 TM, Landsat 7 ETM+和Landsat 8 OLI系列表面反射率遥感影像(表1), 空间分辨率30m, 时间分辨率为16d, 该数据已经使用LEDAPS进行了大气校正, 包括使用CFMASK生成的云、影、水和雪掩膜以及每像素饱和度掩膜等[21].
表1 遥感影像信息

1.3 研究方法
(1)海岸带植被指数计算. 归一化植被指数(Normalized Difference Vegetation Index, NDVI)是区分植被与非植被(光滩、水体)的指示[27]. 增强植被指数(Enhanced Vegetation Index, EVI)是改进的植被指数, 它减少了大气影响, 消除了冠层背景信号[28], 在高生物量区域NDVI饱和时, 具有更高的敏感性. 陆地表面水指数(Land Surface Water Index, LSWI)对植被含水和土壤水有很强的敏感性.

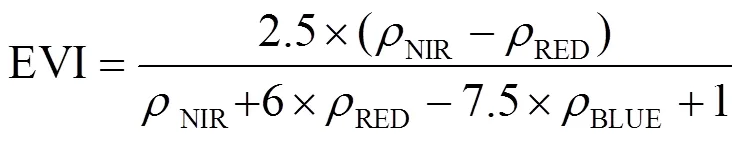

(2)海岸带植被提取算法. NDVI与叶面积指数关联较大, 但在灌木层上却表现为饱和[29]. EVI会根据蓝色波段反射率来调整红色波段反射率, 顾及大气污染、土壤条件变化和冠层背景反射率. LSWI用于识别植被和排除水体. 因此采用EVI≥0.1, NDVI≥0.2且LSWI>0来识别植被信息[30]. 由于潮汐动力周期性、沿海植被物候变化和劣质遥感观测的影响, 从一个图像或合成图像获取的湿地分类结果不能捕捉提取海岸带地物信息. 因此, 为减少潮汐动力和植被物候对海岸带制图的影响, 研究基于时间序列Landsat影像有效频率来识别海岸带植被区和非植被区(潮滩和海水). 参考中国海岸带潮滩轨迹研究[29], 使用植被频率阈值≥5来甄别杭州湾南岸植被.


(3)海岸带植被景观格局变化分析. 景观指数可浓缩海岸植被演化信息, 对海岸带植被景观结构的组成和空间配置特征进行表征, 研究选取聚集度指数(COHESION)、斑块数(NP)、最大斑块数(LPI)、形状指数(LSI), 借助Fragstats 4.2软件对杭州湾南岸植被格局进行甄别.
(4)杭州湾南岸植被区域范围. 海岸带是连接陆地和海洋边缘的两大生态系统交错带, 包括自然地物(河流、海水、潮上带植被、潮间带盐沼和潮下光滩等)和人工地物(湖泊、盐田、农田、养殖塘和建筑等)[7]. 其中, 湿地植被是海岸带蓝色碳汇的典型载体, 同时由湿地植被转化而来的农田也具有较高的固碳能量和潜力, 可作为重要的碳汇类型补充海岸带蓝碳. 1974年以来, 杭州湾共计围垦面积已经超过851.03km2 [32], 垦区线海向利用方式以海水养殖、淡水种植业和工业为主. 植被在自然湿地与人工湿地区域均有分布, 基于海岸带地物和岸线指标综合特征[7], 以研究起始时间为基准, 借助ArcGIS 10.3软件结合Google Earth影像, 提取1984年垦区堤防线作为岸向界线, 并顾及植被物候和34年来潮汐海水淹没频率(图2), 最终确定研究范围是1984年垦区堤防线至滩涂向海延伸, 南北以沿海乡镇为界形成海向敞开的半闭合区域(图1、图2), 包括潮间上带植被、潮间带盐沼、围垦区农田等地物类型.
2 结果与讨论
2.1 杭州湾南岸植被空间分布特征
2.1.1杭州湾南岸植被2018年空间分布

2.1.2杭州湾南岸植被线年度空间分布特征
海岸线作为重要的海岸地貌标志, 是海岸带开发活动的重要依托空间, 其属性在一定程度上代表了海岸人工化趋势[31]. 杭州湾南岸地处亚热带季风气候区, 雨量充沛, 土壤脱盐过程较快, 为开垦滨海沼泽创造了独特的自然禀赋条件, 该区域潮间上带人类沼泽开发利用已有20多年历史[32], 但由于人类活动和自然演化等多因素交互影响的差异, 各镇邻近区域海岸植被分布和演化过程不均匀. 具体来讲, 人类围垦以后在垦区范围内人工植被如农业种植作物会连片规则分布, 而自然植被如互花米草等则会在潮间带淤泥质光滩上呈斑块状生长发育, 并随时间推移连片分布直到斑秃消亡. 此外, 植被区外缘垂直于海岸的变化部分会出现向海扩张和向岸侵退的周期性[33], 特别是当海岸受侵蚀后退时会在植被和光滩间产生陡坎, 造成盐沼植被减少. 总体来说, 杭州湾南岸人工植被呈面状向海推进, 指示海岸淤长堆积, 而近海潮间带植被呈团状连片或稀疏碎片, 指示海岸侵蚀或兼有堆积特征. 因此, 比较1984至2018年不同镇邻近区域岸段植被线的特征, 可指示杭州湾南岸不同位置海岸的侵蚀和堆积格局(图4).

图2 杭州湾南岸植被范围

图3 2018年杭州湾南岸植被分布
2.2 杭州湾南岸植被变化特征
2.2.1杭州湾南岸植被年际动态
海岸植被利于海岸防护, 一方面波浪和潮汐能量在此降低以至消失使海岸所受侵蚀作用减轻, 海水中悬浮物质进入海岸植被遭拦截后沉积使滩面加高延伸, 在陆海物质流动和能量迁移中起着过滤作用; 另一方面作为一种资源与海岸带其他有利物质一起构成了近海生物食物链环节. 因此, 植被年际变化趋势需受到重视.
对研究时段内杭州湾南岸植被时空变化数据进行分析, 可揭示海岸植被动态及扩散特征. 将各期海岸植被数据与镇级行政区划数据叠加, 得到近34年各镇邻近区域植被面积演化情况(图5). 可以看出, 1984到2018年发生了较大变化, 总体呈现波动上升趋势, 海岸植被面积从1984年的7.26 km2上升到了2018年的468.35km2, 这是由于围垦后农田等人工植被增加, 且有少量林地和人工引种的互花米草等在海岸扩张引起. 依据海岸植被年际变化趋势特征, 将其增长划分为5个阶段. 其中第Ⅰ阶段(1984~1994年), 1994年为1984年的3.78倍; 第Ⅱ阶段(1995~2008年), 2008年为1995年的2.06倍; 第Ⅲ阶段(2009~2012年), 2012年为2008年的1.56倍; 第Ⅳ阶段(2013~2014年)净增长了4.38km2; 第Ⅴ阶段(2015~2018年), 2015年是2018年的1.04倍. 就年变化率而言, 从第I阶段1994年的25.26%下降至第II阶段2008年的7.57%, 逐渐降低到第V阶段2018年的1.1%. 可见, 海岸围垦减少了潮间滩涂植被带的宽度, 同时人工引种的互花米草和围垦区陆地的人类农业种植等又扩大了植被面积.
2.2.2杭州湾南岸植被景观格局连通性态势
景观构成单元斑块面积与数量变化会引起景观格局较大动态, 依据景观格局指数计算结果(图6), 研究期间杭州湾南岸植被景观指数变化态势有所差异, 斑块个数占总百分比数由1984年的55%升至2004年的60%, 到2018年下降至31%, 可见, 杭州湾南岸植被在人类围垦和农业活动影响下斑块出现波动破碎化趋势. 杭州湾南岸植被形状指数在1984~2018年期间出现逐渐减小趋势, 其中在1996年以后大幅减小; 同时聚集度指数在研究时段内出现先减小后增大的趋势, 但变化幅度较小, 说明海岸植被斑块形状越来越趋向规则化. 需要指出的是, 1984到2004年聚集度指数在降低后又逐渐升高, 说明2004年以后农业活动和围垦后沿海滩涂互花米草等植被趋于成片分布, 使其聚集度升高.

图4 1984~2018年杭州湾南岸植被线动态

图5 杭州湾南岸植被面积变化特征

图6 1984-2018年杭州湾南岸植被景观格局指数特征
不同性质的海岸类型其植被演化影响因素有所差异(表2). 人类围垦是影响淤泥质河口海岸资源环境演化的干扰因素之一, 一方面使潮间带盐沼植被生境变化导致植被面积减少, 另一方面受人类围垦活动影响, 河口与滨海湿地丧失, 滩涂生境破坏, 陆域环境取代潮滩环境. 在淤泥质杭州湾南岸, 植被因其生物特性对地貌演变过程中的固沙、消能等有重要影响, 反过来地貌环境对植被生长和消亡也至关重要. 另外, 淤泥质海岸潮滩分带性明显, 潮间中下带多为光滩, 而潮上带多为盐沼植被(互花米草等), 伴随着杭州湾南岸围垦造地和海堤防护建设等人类活动, 人工植被扩张速率远高于自然植被演替. 因此, 需要科学合理地规划海岸带防护林体系建设, 以应对海平面上升、滨海洪涝等潜在风险.
3 结论
杭州湾南岸作为连接陆地与海洋生态系统的关键交错带, 是高强度人类活动和全球气候变化双重影响的空间单元, 也是全球重要的湿地资源之一. 该区域潮汐动力、地质地貌以及气候条件为互花米草等植被生长发育提供了优越环境. 近34年来, 杭州湾南岸人类围垦面积较多, 滩涂围垦与潮沟裁弯取直等导致海岸植被带景观向海推移. 人工引种互花米草等在一定程度上弥补了因围垦而减少的潮间带盐沼面积, 同时随着围垦区农业活动加剧, 杭州湾南岸植被面积增幅显著, 进一步促进了杭州湾南岸景观生态系统类型的演替速度.
值得指出的是, 杭州湾南岸滩涂淤积、米草引种以及围垦后人工植被种植等, 导致了杭州湾南岸植被群落结构和景观生态特征发生变化, 同时围垦降低了杭州湾南岸原生植被的生态位, 干扰了海岸带植被景观自然演替序列, 使海岸带湿地整体生态系统服务功能受到冲击. 以互花米草为代表的种群较强的入侵力和杭州湾南岸湿地生态系统的可入侵性, 使人工引种的海岸带植被由最初的担负促淤护滩使命, 转向至外来入侵物种角色. 因此, 需要连续评估监测盐沼植被生态环境效应, 对标国际研究计划“未来地球海岸”的陆海统筹蓝图重绘战略, 将其纳入到滨海湿地整治修复实施和海岸带保护与开发规划中, 稳步落实生态保护红线制度和自然岸线占补平衡机制在大湾区滨海湿地区域的实践, 以服务未来海岸弹性发展.
表2 中国海岸带植被演化影响因素分析
[1] Jevrejeva S, Jackson L P, Riva E M, et al. Coastal sea level rise with warming above 2℃[J]. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113:13342-13347.
[2] 胥为, 周云轩, 沈芳, 等. 基于Sentinel-1A雷达影像的崇明东滩芦苇盐沼植被识别提取[J]. 吉林大学学报(地球科学版), 2018, 48(4):1192-1200.
[3] Li J L, Yang L, Pu R L, et al. A review on anthropogenic geomorphology[J]. Journal of Geographical Sciences, 2017, 27(1):109-128.
[4] 李加林, 刘永超. 人工地貌学学科体系框架构建初探[J]. 地理研究, 2016, 35(12):2203-2215.
[5] 刘永超, 李加林, 袁麒翔, 等. 人类活动对象山港潮汐汊道及沿岸生态系统演化的影响[J]. 宁波大学学报(理工版), 2015, 28(4):120-123.
[6] 刘永超, 李加林, 袁麒翔, 等. 人类活动对港湾岸线及景观变迁影响的比较研究—–以中国象山港与美国坦帕湾为例[J]. 地理学报, 2016, 71(1):86-103.
[7] 李飞, 曹可, 赵建华, 等. 典型海岸线指标识别与特征研究—–以江苏中部海岸为例[J]. 地理科学, 2018, 38(6):963-971.
[8] 李加林, 田鹏, 邵姝遥, 等. 中国东海区大陆岸线变迁及其开发利用强度分析[J]. 自然资源学报, 2019(9): 1886-1901.
[9] 唐剑武, 叶属峰, 陈雪初, 等. 海岸带蓝碳的科学概念、研究方法以及在生态恢复中的应用[J]. 中国科学: 地球科学, 2018, 48(6):661-670.
[10] 王秀君, 章海波, 韩广轩. 中国海岸带及近海碳循环与蓝碳潜力[J]. 中国科学院院刊, 2016, 31(10):1218-1225.
[11] 冯佰香, 李加林, 龚虹波, 等. 基于恢复能力与影响周期的围海养殖工程生态损害特征及补偿标准—–以象山县水湖涂名优水产养殖区为例[J]. 自然资源学报, 2019, 34(4):745-758.
[12] Sun C, Fagherazzi S, Liu Y X. Classification mapping of salt marsh vegetation by flexible monthly NDVI time- series using Landsat imagery[J]. Estuarine Coastal and Shelf Science, 2018, 213:61-80.
[13] Webb E L, Friess D A, Krauss K W, et al. A global standard for monitoring coastal wetland vulnerability to accelerated sea-level rise[J]. Nature Climate Change, 2013, 3(5):458-465.
[14] Liu Y C, Liu Y X, Li J L, et al. Evolution of landscape ecological risk at the optimal scale: A case study of the open coastal wetlands in Jiangsu, China[J/OL]. International Journal of Environmental Research and Public Health, 2018, 15(8):1691 [2018-08-08]. https://doi. org/10.3390/ijerph15081691.
[15] Traganos D, Aggarwal B, Poursanidis D, et al. Towards global-scale seagrass mapping and monitoring using sentinel-2 on Google Earth Engine: The case study of the Aegean and Ionian seas[J/OL]. Remote Sensing, 2018, 10(8):1227 [2019-03-22].https://doi.org/10.3390/rs1008 1227.
[16] Schmidt K S, Skidmore A K. Spectral discrimination of vegetation types in a coastal wetland[J]. Remote Sensing of Environment, 2003, 85(1):92-108.
[17] Gedan K B, Kirwan M L, Wolanski E, et al. The present and future role of coastal wetland vegetation in protecting shorelines: Answering recent challenges to the paradigm [J]. Climatic Change, 2011, 106(1):7-29.
[18] 刘明月. 中国滨海湿地互花米草入侵遥感监测及变化分析[D]. 长春: 中国科学院东北地理与农业生态研究所, 2018.
[19] Sun C, Liu Y X, Zhao S S, et al. Classification mapping and species identification of salt marshes based on a short-time interval NDVI time-series from HJ-1 optical imagery[J]. International Journal of Applied Earth Observation and Geoinformation, 2016, 45:27-41.
[20] Goodchild M F, Guo H, Annoni A, et al. Next-generation digital earth[J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(28): 11088-11094.
[21] Gorelick N, Hancher M, Dixon M, et al. Google earth engine: Planetary-scale geospatial analysis for everyone [J]. Remote Sensing of Environment, 2017, 202:18-27.
[22] Hansen M C, Potapov P V, Moore R, et al. High- resolution global maps of 21st-century forest cover change[J]. Science, 2013, 342(6160):850-853.
[23] Pekel J F, Cottam A, Gorelick N, et al. High-resolution mapping of global surface water and its long-term changes[J]. Nature, 2016, 540:418-422.
[24] Dong J W, Xiao X M, Menarguez M A, et al. Mapping paddy rice planting area in northeastern Asia with Landsat 8 images, phenology-based algorithm and Google Earth Engine[J]. Remote Sensing of Environment, 2016, 185:142-154.
[25] Murray N J, Phinn S R, DeWitt M, et al. The global distribution and trajectory of tidal flats[J]. Nature, 2019, 565:222-225.
[26] 孟梦, 田海峰, 邬明权, 等. 基于Google Earth Engine平台的湿地景观空间格局演变分析: 以白洋淀为例[J]. 云南大学学报(自然科学版), 2019, 41(2):416-424.
[27] Pettorelli N, Vik J O, Mysterud A, et al. Using the satellite-derived NDVI to assess ecological responses to environmental change[J]. Trends in Ecology & Evolution, 2005, 20(9):503-510.
[28] Huete A, Didan K, Miura T, et al. Overview of the radiometric and biophysical performance of the MODIS vegetation indices[J]. Remote Sensing of Environment, 2002, 83(1/2):195-213.
[29] Xiao X, Braswell B, Zhang Q, et al. Sensitivity of vegetation indices to atmospheric aerosols: Continental- scale observations in Northern Asia[J]. Remote Sensing of Environment, 2003, 84(3):385-392.
[30] Xiao X M, Biradar C M, zarnecki C, et al. A simple algorithm for large-scale mapping of evergreen forests in tropical America, Africa and Asia[J]. Remote Sensing, 2009,1(3):355-374.
[31] Wang X, Xiao X M, Zou Z H, et al. Tracking annual changes of coastal tidal flats in China during 1986-2016 through analyses of Landsat images with Google Earth Engine[J/OL]. Remote Sensing of Environment [2018- 12-15]. https://doi.org/10.1016/j.rse.2018.11.030.
[32] 花一明. 杭州湾滩涂围垦及利用动态遥感监测研究[D]. 杭州: 浙江大学, 2016.
[33] 侯西勇, 毋亭, 侯婉, 等. 20世纪40年代初以来中国大陆海岸线变化特征[J]. 中国科学: 地球科学, 2016, 46(8):1065-1075.
[34] 陈吉余, 杨世伦, 张勇, 等. 中国海滨沼泽的初步研究—–纪念竺可桢师诞辰一百周年[J]. 地理科学, 1990(1): 58-68.
[35] 张长宽, 龚政, 陈永平, 等. 潮滩演变研究进展及前沿问题[C]. 第十八届中国海洋(岸)工程学术讨论会, 舟山, 2017.
Dynamic monitoring of vegetation in southern Hangzhou Bay from time series images
LIU Yongchao1,2,3, LI Jialin4, ZHANG Yu5, ZHAO Bingxue1,6, XU Wenxuan1, HE Gaili1, LIU Yongxue1,2,3*
( 1.School of Geography and Ocean Science, Nanjing University, Nanjing 210023, China; 2.Key Laboratory of Coastal Zone Exploitation and Protection of Ministry of Natural Resources, Nanjing 210024, China; 3.Collaborative Innovation Center of South China Sea Studies, Nanjing University, Nanjing 210023, China; 4.Donghai Institute, Ningbo University, Ningbo 315211, China; 5.School of geography, Nanjing Normal University, Nanjing 210023, China; 6.College of geography and planning, Chizhou University, Chizhou 247000, China )
Coastal vegetation is a key element in connecting land and sea ecosystems. Under the background of global warming, sea-level rise (SLR) and increased human activities, mastering the evolution law of coastal vegetation is the basis for supporting the elastic development of the future coastal zones. Taking the south bank of Hangzhou Bay in the West Coast of the Pacific Ocean as the sampled research area, the vegetation dynamics of the Southern Hangzhou Bay (SHB) from 1984 to 2018 were monitored by time-series Landsat remote sensing images. The results show that the vegetation area on the SHB has increased from 7.26km2in 1984 to 468.35km2in 2018, mainly undergoing evolutionary stages from growth and development to rapid expansion then back to slow growth. Under the influence of cofferdam, artificial introduction and sedimentation, the coastal vegetation is distributed perpendicularly to the seawall along the coast with sparse plaques, and its landscape composition and succession characteristics are significant. The study also shows the advantages and potential of Google Earth Engine for continuous monitoring of coastal vegetation. The study reveals that the characteristics of vegetation change on the SHB under the background of human activities are extremely important for maintaining the safety of the coastal wetland ecosystem and the sustainable development of the super bay area.
time series analysis; vegetation dynamics; reclamation; artificial planting; Southern Hangzhou Bay
Q149; X826
A
1001-5132(2020)01-0025-07
2019−09−30.
宁波大学学报(理工版)网址: http://journallg.nbu.edu.cn/
国家自然科学基金(41971378); 江苏省杰出青年基金(BK20160023); NSFC-浙江两化融合联合基金(U1609203); 安徽省高校
自然科学项目(KJ2019A0866).
刘永超(1990-), 男, 甘肃庆阳人, 在读博士研究生, 主要研究方向: 海岸带资源环境演化. E-mail: lycgeo@163.com
刘永学(1976-), 男, 江苏扬州人, 博士/教授, 主要研究方向: 资源环境遥感. E-mail: yongxue@nju.edu.cn
(责任编辑 韩 超)
