苦瓜强雌系品种的选育研究
2020-01-08王国莉
王国莉
(惠州学院 生命科学学院,广东 惠州 516007)
苦瓜(Momordica charantia L.)不仅营养丰富[1],还兼具药用[2]和保健功能[3],在我国栽培历史悠久,主要以广东、广西、福建、湖南、江西等省的栽培面积最广[4].随着苦瓜消费市场的需求不断扩大,苦瓜高产、抗性新品种的选育迫在眉睫.但苦瓜的遗传育种研究起步较晚,我国还没有系统地对苦瓜种质资源开展研究工作.虽然广东、湖南及江西的科研人员相继选育出了一批综合性状优良的苦瓜一代杂种,如热研2 号、穗新2 号、湘丰1 号、绿美、川苦6 号等新品种,但是这些品种还远远不能满足市场需要.因此,培育早熟、高产、优质的苦瓜新品种是目前苦瓜生产上急需解决的重要问题.强雌系苦瓜品种选育是苦瓜育种的一个主要方向,强雌系苦瓜在育种上(全雌系为雌花率100%;强雌系则为雌花率在80%以上)可以提高品种的早熟性、丰产性,简化制种程序,降低成本并提高F1 代种子质量.通过与具有其他优良性状的亲本杂交,可间接培育出早熟、丰产、优质等综合性状优良的F1 代杂种[5-6].
关于苦瓜性别分化和性别决定基因的研究方兴未艾.赵志伟等[7]的研究再次证明了苦瓜的全雌性状是由1 个隐性基因控制的,但受一些微效多基因的控制.王日升等[8]以全雌系苦瓜花为材料,获得了全新的Mc-ACS4序列,跟其他ACS序列一样可能与苦瓜性别分化有关.李明珠等[9]的研究发现与苦瓜性别分化有关的特异蛋白质,其中11 kD 的蛋白质可能是雌花分化程序表达中的一种关键蛋白,而30 kD 的蛋白质很可能是雄花程序表达中的一种关键蛋白,外界因素和环境因子均能影响这些蛋白的表达.汪俏梅和曾广文[10]的研究则发现苦瓜的性别分化首先要经过两性期,然后再分别向雌或雄的方向发育.
孔冬梅[11]、杨䶮等[12]和汪俏梅等[13]的研究说明化学试剂在植物的性别分化中发挥着重要的调控作用,例如赤霉素(GA)、细胞分裂素(CTK)、生长素(IAA)、乙烯、脱落酸(ABA)和表油菜素内酯(epi-BL),均对苦瓜的性别分化有着重要的影响.王日升等[14]都得出AgNO3具有促雄效果,可以成功诱导全雌花分化出双性花,并得出三叶期是全雌性苦瓜性别分化的关键时期.汪俏梅等[13]提出环境因素中温度和光周期是主要影响,短日照(8 h)、低温作用于苦瓜苗期,可使植株的发育提早,第一雌花节位降低,雌花数增多.万翔等[16]在黄瓜的研究中指出雌性系开少量雄花或基本不开雄花,致使雌性系自身的育种变得困难,国内主要使用化学调控方法.
随着对苦瓜强雌系育种的研究不断深入,加之种植强雌系苦瓜的经济效益好,因此,强雌系品种在苦瓜生产和育种研究上都有重要的作用,对新型强雌系苦瓜品种的选育显得十分迫切.本研究以常规育种方法为主,开展新型强雌系苦瓜品种的初步选育工作,对尽快培育出苦瓜高产、优质新品种具有重要的理论和现实意义.
1 材料及方法
1.1 材料
试验材料为课题组自有的强雄性苦瓜品种Qx001,强雌性苦瓜品种Mc001和全雌性品种Qc001.
1.2 研究方法
1.2.1 苦瓜育苗
2014 年春,取3 个品种种子适量,参照王日升等[14]、陈坚和王霞[15]的育种和育苗方法.将种子放入提前预热至55℃的温水中,玻璃棒搅拌10 min后,自然冷却浸泡12 h.浸泡结束后捞出种子,用镊子将种皮捏裂,放入智能人工气候培养箱内,设定温度30℃,湿度80%进行催芽.大概两天种子露白后,点播在穴盘中,29℃进行育苗.
1.2.2 定植和处理
当苦瓜长至3 片真叶时,将所有苦瓜苗移栽到试验田中,当全雌性苦瓜幼苗长至三叶一心期时,喷洒(300 mg·L-1)AgNO3溶液记录时间,一星期后再喷洒一次.
1.2.3 田间管理及杂交
定期对苦瓜进行浇水、除草和施肥.苦瓜开花前一天,进行套袋处理,第二天进行授粉并记录花性和节位.
选择发育正常的苦瓜花进行杂交.雄花可明显看见花药和花丝,雌花则可见其柱头,进行强雌(♂)×全雌(♀)、强雄(♂)×全雌(♀)杂交,强雌植株自交授粉,强雄植株自交授粉.早上9 点前完成授粉,6 点开始隔1小时授粉一次,总共3次,授粉结束后记录亲本的花性和节位,悬挂标签做好记录.
观察杂交后代田间性状表现,选择具有典型性状的12株进行数据统计.
1.2.4 收获第一代杂交种并育苗
当苦瓜成熟、果肉变红或橙红,即可收获苦瓜种子,洗净晾干保存,即为第一代杂交种.
2015 年春,将收获的杂交F1代种子育苗,播种并进行性状观察,筛选强雌系株系自交授粉,获得F1代自交种F2.
1.2.5 选取第二代强雌株的种子进行育苗
2016 年春,F2代育苗播种,再进行性别定向选择,自交后获得强雌系苦瓜育种新品系.
2 结果与分析
2.1 杂交后代F1性别表现
全雌(♀)×强雌(♂)F1代主茎30 节内雌雄花统计结果见表1.由表1 看出,强雌∶非强雌=2∶21,总雌花∶总雄花=180∶358≈1∶2,13 号和23 号表现为强雌株,第一朵花和第一雌花节位分别在5 和7 节位,雌花率为92.0%和83.3%,说明强雌(♂)×全雌(♀)的F1 代雄性比率较高,性别分化比例约为1∶2,推测全雌品种基因为不完全隐性.13号的节位更低,雌性更强,为潜在的强雌系材料.

表1 全雌(♀)×强雌(♂)F1代主茎30节内雌雄花统计
强雄(♂)×全雌(♀)F1代主茎30 节内雌雄花统计结果见表2.由表2 看出,性别表现强雌∶非强雌=1∶4,总雌花∶总雄花=235∶221≈1∶1.强雌株的第一朵花和第一朵雌花节位相同,10 号为第7 节位,16 号为第5 节位,18 号为第6 节位,19 号为第7 节位,雌花率10 号为91.3%,16号为87.5%,18号为85.7%,19号为91.7%.说明强雄(♂)×全雌(♀)杂交后F1代基因型表现,类似Aa×aa 的杂交结果,与孟德尔基因分离相似,10 和19号雌花率高,但第一雌花节位比16和18号高,但19号雌性最强,为潜在的强雌系材料.
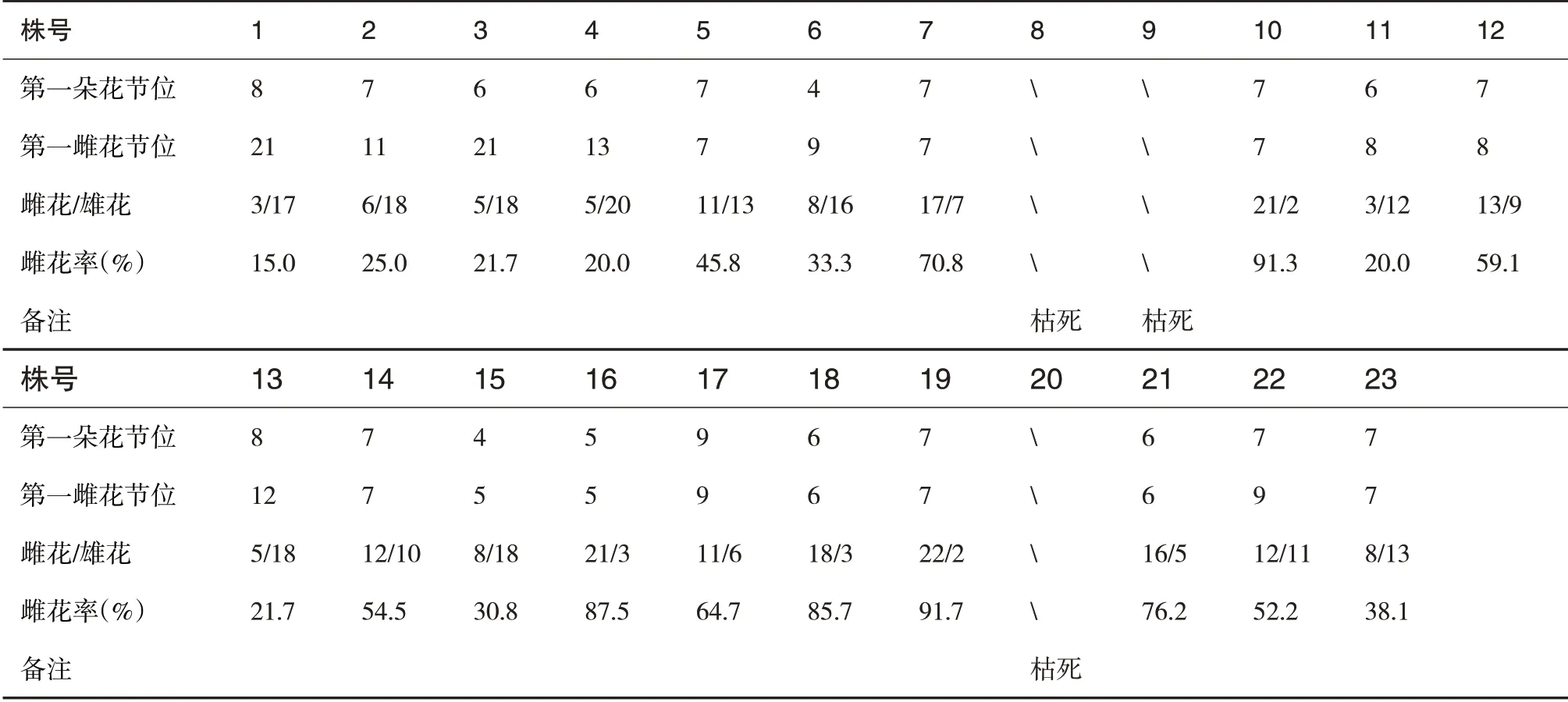
表2 强雄(♂)×全雌(♀)F1代主茎30节内雌雄花统计
强雌品种自交F1代主茎30 节内雌雄花统计结果见表3.由表3 看出,强雌∶非强雌=0∶22,总雌花∶总雄花=96∶469≈1∶5,无强雌株后代.结果表明,强雌自交后代雌性很弱,普遍第一朵花节位降低,第一雌花节位升高,说明母本可能为基因型不稳定的强雌系种子.无潜在强雌系材料.

表3 强雌自交F1代主茎30节内雌雄花统计
强雄品种自交F1代主茎30 节内雌雄花统计结果见表4.由表4 看出,强雌∶非强雌=1∶20,总雌花∶总雄花=51∶417≈1∶8,只有17号为强雌株,第一朵花和第一雌花节位为第7 节位,雌花率为91.3%,说明强雄自交后代雄性极高.出现强雌株,可能是基因突变或者是雌性基因由隐性基因控制,基因分离比例约为1∶8,接近1∶9,与多基因控制同一性状相似,17 号雌性极高,为潜在强雌系材料.

表4 强雄品种自交F1代主茎30节内雌雄花统计
2.2 杂交后代F2性别表现
强雌品种自交F2代主茎30 节内雌雄花统计结果见表5.由表5 看出,筛选出的强雌F118 号(表2)的自交后代全为强雌系,总雌花∶总雄花=37∶3≈12∶1,但1和2号第一朵花和第一雌花节位都较高,分别为第8和第10 节位,雌花率分别为90.0%和95.0%,说明强雄(♂)×全雌(♀)的F2代基因型稳定,雌性强,基因分离比例为12∶1,与多基因控制同一性状型相似.2号虽然第一雌花节位较高,但雌花率高,可以作为研究所要的强雌系新品种.

表5 强雌自交F2代主茎30节内雌雄花统计
3 结论与讨论
目前研究瓜类的性状遗传多在质量性状上研究,不能区分多基因效应.赵志伟等[7]研究苦瓜强雌性状遗传分析表明,全雌性状由一个隐性单基因控制,强雌性状由一对不完全显性基因控制.邹晓艳[16]首次将主基因-多基因混合遗传模型用于瓜类的花性型研究,结果表明黄瓜的全雌性性状和强雌性性状均是由1 对不完全显性主基因控制,而且存在微效多基因的调控.闫立英等[17]应用该法发现雌雄同株黄瓜单性结实性表现为不完全显性遗传,受1 对加性主基因+加性-显性多基因控制.刘莉等[18]利用该模型对西瓜强雌性的6 世代群体的雌花节率性状进行联合分析,发现西瓜强雌性性状遗传受2 对主基因的加性-显性-上位性模型控制,主基因表现为隐性.
表1 和表2 中,与全雌品种杂交后,F1中的强雌后代数量明显增多,其中强雄(♂)×全雌(♀)的强雌后代最多,与预期强雌(♂)×全雌(♀)后代最多不一样,而强雌自交F1代没有出现强雌株,强雄自交出现1 株强雌株,说明控制强雌性状的基因不完全显性,存在其他控制基因,与赵志伟等[7]、邹晓艳[16]、闫立英等[17]和刘莉等[18]的研究结论相近,即苦瓜基因遗传与加性主基因+加性-显性多基因控制系统相似.
由表1-5 看出,强雌系植株第一雌花节位全为第一朵花节位,并且节位低,说明全部潜在强雌株均为早熟丰产型苦瓜.表5 中总雌花∶总雄花=37∶3≈12∶1,证明研究所得强雌株中控制强雌性状的基因分离达到预期结果,雌花率极高,说明了利用雌性强的苦瓜品种杂交,可获得雌性很强的新品种,并且能加快选育新型的强雌系品种.
王日升等[14]利用AgNO3进行化学调控成功诱导出双性花并获得具有发芽力的种子.本研究也利用AgNO3进行了全雌系的留种实验,发现喷洒2 次300 mg·L-1AgNO3深液后,成功诱导出双性花,并进行同性姊妹株之间的授粉,早上9 点前完成授粉,隔1 小时授粉一次,总共3次,苦瓜子房能正常发育并获得全雌性种子,证明花粉可育.但双性花节位在主茎14 节左右变为雌花,而增加AgNO3溶液的喷洒次数可提高双性花的节位.结果表明化学调控可用于保存珍贵的实验材料.
由于大田育种会受到多方面因素的影响,汪俏梅等[13]研究指出温度和光周期会影响苦瓜的性别分化,研究中因为种植季节环境的变化有可能会对试验结果造成影响,加之因为意外损失果实,也导致试验结果有失偏颇.
