增温和增氮及其交互作用对藏北高寒草甸植物群落结构 与物种多样性的影响
2019-12-25葛怡情王学霞闫玉龙干珠扎布胡国铮王子欣李岩水宏伟杨劼高清竹
葛怡情 ,王学霞,闫玉龙,干珠扎布,胡国铮,王子欣,李岩, 水宏伟,杨劼,高清竹*
1. 内蒙古大学生态与环境学院,内蒙古 呼和浩特 010021;2. 中国农业科学院农业环境与可持续发展研究所,北京 100081; 3. 北京农林科学院,北京 100081;4. 安徽师范大学地理与旅游学院,安徽 芜湖 241000
IPCC 第五次评估报告结果表明,由于温室气体排放量增加导致的气候变暖问题日益严重,尤其是高纬度和高海拔地区更为明显(Pachauri et al.,2014)。随着工业化水平的提高、含氮肥料的大量生产和使用,导致了排向大气中的含氮化合物激增(Liu et al.,2016),中国已经成为继欧洲、美国之后的世界第三大气氮沉降集中区(顾雪峰等,2016)。青藏高原是中国重要的生态安全屏障和战略资源储备基地(杨元合,2018),因其海拔高度平均在4 000 m 以上,生态系统脆弱性显著。目前青藏高原温度升高趋势明显(Gao et al.,2013),自1960 年以来,该区域温度每10 年增加0.2 ℃(Chen et al.,2013),预计到2049 年,温度将升高1.5 ℃,到2099 年时,温度将增加2.9 ℃(Liu et al.,2009)。同时由于南亚地区工业和交通排放的活性氮,加上印度季风的传输,使得青藏高原的氮沉降量逐年增加(Liu et al.,2013;Pei et al.,2009)。
温度升高一方面通过影响光合、呼吸等生理过程来影响植被的生长发育,另一方面通过影响土壤水分含量来改变其生长及生物量的分配,从而改变植被群落特征(Yang et al.,2011;Melillo,2002;Ganjurjav et al.,2016)。在青藏高原东北部增温实验中发现,短期增温使高海拔地区物种多样性急剧下降(Klein et al.,2004),由于高寒地区植物生长高度较低,凋落物分解的缓慢与积累,抵挡了到达植物的光合辐射,从而抑制植物的生长,减少物种多样性。氮素是限制陆地生态系统生产力的重要元素之一(Gross et al.,1998;Tilman,1993),研究表明,不同功能群植物应对外源氮输入的响应也不同(Bai et al.,2010)。氮添加使禾本科植物生物量显著增加,而杂类草和豆科植物的生物量却随着氮添加水平的增加而减少,群落的物种丰富度显著降低(张杰琦等,2010)。交互作用下植被高度、生物量均显著高于对照处理,并表明交互作用有助于植物生长;增温处理下,植物物种多样性增加,土壤湿度降低(郭红玉等,2015)。温度升高在一定程度上能满足植物对热量的需求(Harte et al.,1995;周兴民等,2000),有利于植物的生长发育。而氮添加主要有铵态氮、硝态氮两种形式,研究发现,这两种氮素均能够促进植物生长(周国英等,2004),因此认为其交互作用有利于植物生长(宗宁等,2014)。而在内蒙古荒漠草原的研究中发现,对于植物的盖度,增温呈负效应,增氮呈正效应,两者交互作用相互抵消,所以交互作用对于其盖度无显著影响(李元恒,2014)。
基于前人对于青藏高原草地生态系统的研究,发现温度升高与氮沉降量的增加,使土壤含水量及养分含量发生变化,温度升高,土壤含水量会下降,氮素增加,有时会超过植物群落生产的饱和阈值,进而发挥不了它应有的效果;也有因为氮素的输入,促进了矿化作用,为植被生长提供更多的氮素需求。为了解在未来温度升高、氮沉降增加的趋势下,高寒草甸植物群落结构与多样性的变化趋势,本研究选取西藏自治区那曲市典型高寒草甸进行增温增氮实验,探讨增温和氮沉降对高寒草甸生产力、物种多样性的影响,有助于明确未来增温与氮沉降对高寒草甸生态系统功能与结构的变化。
1 材料与方法
1.1 研究区概况
试验区位于藏北地区那曲市那曲镇(31.441°N,92.017°E),海拔4 500 m 左右,位于唐古拉山脉、念青唐古拉山脉和冈底斯山脉之间。气候属于典型的亚寒带气候。该地区雨热同季,月平均温度在5-9 月高于0 ℃,其中90%以上的降水集中在该时段。样地主要以小嵩草(Kobresia pygmaea)和青藏苔草(Carex moorcroftii)为建群种;早熟禾(Poapratensis)为主要的禾本科植物;主要的杂草包括钉柱委陵菜(Potentilla saundersiana)、紫花棘豆(Oxytropis subfalcata)等。
1.2 实验设计
1.2.1 样地设置
样地内共设4 个处理,对照(CK)、增氮(N,40 kg·hm-2·a-1)、增温(W)、增温+增氮(WN),每个处理4 个重复,共16 个小区,每个样方大小为2 m×2 m,相邻样方之间设置2 m 的缓冲隔离带。
1.2.2 实验处理方法
(1)对照(CK):自然样地,4 个实验小区,大小为2 m×2 m,相邻样方之间设置2 m 的缓冲隔离带。
(2)增温(W):2014 年5 月采用开顶式生长室(open top chamber,OTC)模拟增温。
(3)增氮(N):氮素添加剂量参照中国氮沉降分布的格局(张杰琦等,2010),西藏地区的干湿沉降率为7 kg·hm-2·a-1,所以本试验添加浓度是模拟自然沉降量的6 倍,到2050 年的沉降量约为40 kg·hm-2·a-1。使用NH4NO3模拟大气氮沉降,于2014 年5 月开始,在每年的5、6、7、8 月中旬以溶液的形式给样地喷洒,将26 gNH4NO3溶解在5 L水中,均匀喷洒在样地中,然后再用5 L 清水冲洗残留在叶片上的氮素,此外对照与增温处理喷施同样剂量的水以避免外加的水造成的影响(每年每个小区总用水量40 L,相当于年降雨量的1%(闫玉龙,2019)。
(4)增温+增氮(WN):在增温的基础上,加入增氮处理。
1.3 测定项目与方法
1.3.1 生物量测定
地上植物按其功能群分为禾本科(Gramineae)、莎草科(Cyperaceae)、豆科(Leguminous)、杂类草(Forbs)。采用无损测量法测定每个样方的地上生物量(Yan et al.,2018)。于2017-2018 年每年8 月中旬植被生长季盛期,在每个小区内选取0.5 m×0.5 m 的样方,调查不同植物功能群种类组成、高度和盖度等。在试验样地外选取20 个相同规格的校正样方,调查功能群植物种类组成、高度和盖度等,齐地面收集地上植物,分功能群装入信封中,带回实验室,置于105 ℃烘箱中杀青0.5 h,65 ℃下烘干至恒重,测定干重。建立校正样方中每种功能群植物高度、盖度以及生物量回归方程,利用该方程计算各小区中每种功能群植物的生物量。
1.3.2 土壤温度与土壤含水量测定
试验期间,用EM 50 数据收集系统(Decagon Devices,Inc.,NE,USA)记录样方地下5 cm 土壤温度和土壤含水量数据,每隔30 min 记录1 次数据。
1.3.3 数据计算
在测定群落高度和盖度之后,用某种植物高度与群落中所有植物高度之和的比值代表该种植物的相对高度,用某种植物分盖度与群落中所有植物分盖度之和的比值代表该物种相对盖度。用每种植物相对高度和相对盖度计算其重要值(Importance value,IV),并通过IV 计算物种多样性(Yan et al.,2018)。物种多样性采用Shannon-Weiner 指数(H)、Simpson 指数(D)和E. Pielou 均匀度指数(J)表示(干珠扎布等,2015),计算公式如下:
Shannon-Weiner 指数(H):

Simpson 指数(D):

E. Pielou 均匀度指数(J):
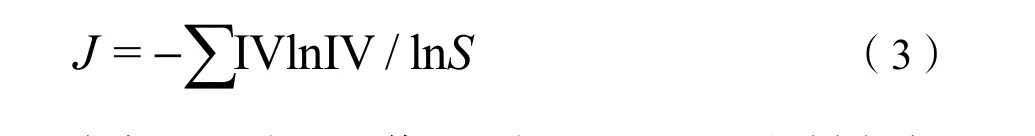
式中,IV 为重要值;S 为0.5 cm×0.5 的样方内出现的物种数。
1.4 数据分析
高寒草甸地上生物量、功能群生物量、物种丰富度、物种多样性指数在增温、增氮处理间的差异采用三因素方差分析。采用回归分析方法对生物量、物种丰富度和土壤温度、土壤含水量进行相关性分析。统计分析均于SPSS 24.0(SPSS.Chicago,USA)中实现,统计图形在Excel 2010 中绘制。
2 结果与分析
2.1 增温增氮对土壤温度和土壤含水量的影响
土壤地下5 cm 温度与含水量有明显的年际变化,W、WN 处理下,土壤温度均显著增加,土壤含水量均显著下降(P<0.05)。2017 年,W、WN处理土壤温度分别增加0.46 ℃和0.53 ℃,2018 年W、WN 处理土壤温度分别增加0.81 ℃和0.88 ℃;2017 年,W、WN 处理土壤含水量分别下降7.02%和6.74%,2018 年,W、WN 处理土壤含水量分别下降6.90%和5.97%(表1)。
2.2 增温增氮对各功能群地上生物量的影响
高寒草甸生物量存在显著的年际变化(图1),CK 样地从2017 年的40.76 g·m-2上升至2018 年的65.56 g·m-2。相比于2017 年,2018 年N 处理下生物量增加25.3%,总生物量升高17.9%。在不同处理间,N、W、WN 处理均显著高于CK(P<0.05)。除总生物量呈年际变化外,各功能群的生物量也有显著的年际变化。2017 年W 处理下莎草科、禾本科、杂类草的生物量均显著增加,2018 年莎草科生物量在N 处理下生物量显著增加,豆科的生物量在2017 年和2018 年的WN 处理下均显著增加,分别增加了15.06 g·m-2和50.54 g·m-2。相关分析表明,豆科生物量与土壤含水量呈显著负相关(P=0.043,R2=0.1497),即土壤含水量下降,高寒草甸豆科生物量增加。

图1 增温增氮对各功能群生物量及总生物量的影响 Fig. 1 Effects of warming and nitrogen addition on biomass and total biomass of functional groups
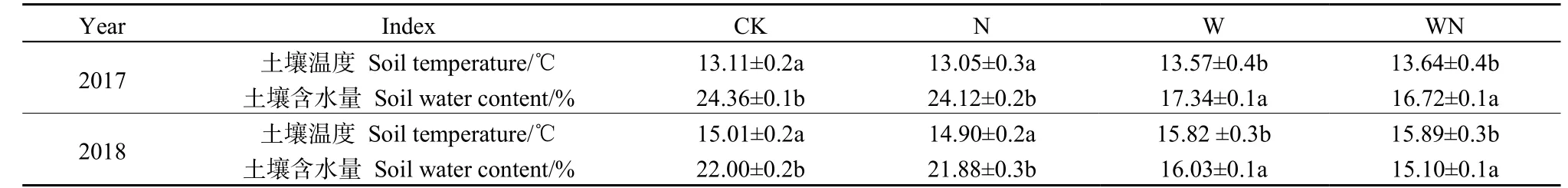
表1 增温增氮对土壤温度和土壤含水量的影响 Table 1 Effects of warming and nitrogen addition on soil temperature and soil water content
对不同处理条件下藏北高寒草甸生物量进行显著性分析(表2),发现生物量在年际间、增温、增氮、增温与增氮的交互作用均有显著性差异(P<0.05);而年与增温、年与增氮、年与增温增氮间均无明显的交互影响(P>0.05)。

表2 不同处理下高寒草甸生物量年际变化显著性 Table 2 Significance of internal variation of alpine meadow biomass under different treatments
2.3 增温增氮对群落高度和盖度的影响
各处理对藏北高寒草甸各功能群的高度和盖度具有明显的影响(图2),2017 年和2018 年在W 处理下禾本科高度均显著增加,增幅分别为55%和54%。W、N 处理下杂类草的高度显著增加,2017年W、N 处理高度增幅为112%和106%;2018 年增幅为101%和131%。2018 年WN 处理下豆科高度显著增加,增幅分达270%。2017 年和2018 年莎草科植物盖度在N 处理下均显著增加,增幅分别达到148%和138%;豆科植物盖度在WN 处理下显著增加,增幅分别达366%和617%。2017 年杂类草的盖度在N 处理下盖度显著增加,增幅为61.4%(P<0.05)。
2.4 增温增氮对物种多样性的影响
藏北高寒草甸物种多样性对不同处理的反应不同。 2017 年各处理对 Simpson 指数、Shannon-weiner 指数、E. pielou 均匀度指数均无显著影响。2018 年WN 处理下Simpson 指数显著降低(P<0.05)。2018 年N 处理使莎草科植物物种丰富度显著增加(P<0.05),WN 处理下莎草科植物物种丰富度显著减少,减少66%(表3)。相关分析表明,Shannon- weiner 指数(P=0.025)、Simpson指数(P=0.040)、E. pielou 均匀度指数(P=0.020)均与莎草科盖度呈显著负相关(表4)。
3 讨论
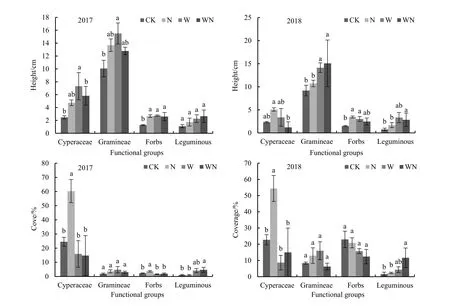
图2 模拟增温增氮对群落高度和盖度的影响 Fig. 2 Effects of warming and nitrogen addition on height and cover

表3 增温增氮对物种多样性的影响 Table 3 Effects of warming and nitrogen addition on species diversity
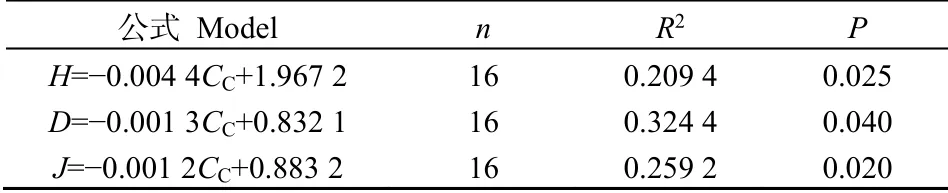
表4 物种多样性指数与莎草科盖度相关性分析 Table 4 Analysis of correlation between species diversity and cyperaceous cover
由于青藏高原处于长期低温环境,土壤有机质矿化速率较慢,使植物和微生物都长期处于贫氮的土壤环境中。有研究认为,大尺度的氮素输入会使生物量显著增加(Lebauer et al.,2008),在植物群落中,有些物种对氮素有明显的偏好,土壤中氮素含量的增加,会明显增加其优势度,加强其对其他物种的竞争力,从而导致群落特征发生变化。草地的物种组成也会使植物的生物量发生改变,由于对有效资源的竞争排斥,尤其是光照、空间、土壤资源的竞争而引起的物种组成的变化,使其生物量发生变化。不同功能群植物的群落特征对氮沉降的反应也不尽相同。本研究发现,氮沉降使总地上生物量、莎草科植物的盖度和地上生物量显著增加,这与许庆民等(2017)和庞晓瑜等(2006)研究结果一致,而沈振西等(2002)研究发现在未来氮沉降的情况下,莎草科植物生物量将会保持较稳定的状态。在交互作用中因增温导致土壤水分含量下降,对莎草科植物的盖度、高度和生物量均无显著影响,但莎草科物种丰富度显著减少66%(P<0.05),而氮沉降使莎草科植物物种丰富度显著增加。相关性分析表明,莎草科植物物种丰富度与土壤含水量呈显著正相关(王长庭等,2017),由此可知,土壤含水量是控制莎草科植物物种丰富度关键因素。研究发现增氮处理使杂草的高度显著增加,对其他群落特征无显著影响,由于氮沉降解除了氮素的营养限制,地上生物量显著提高,同时由于氮素的输入导致资源供应的不对称,改变地上地下的竞争,引起植物组成的变化(Wu et al.,2011)。
温度升高会显著增加土壤的温度及改变其他控制植物生长和微生物活性的因素。温度升高使群落总生物量和禾本科生物量显著增加,对其他功能群生物量无显著影响,低温是限制高寒草甸植物生长的关键因素,温度的升高能够促进高寒草甸植物的生长和生物量的积累以及群落物种多样性的增加(姜炎彬等,2017)。藏北高原的年平均温度仅为-1.16 ℃。植物所处的环境远远低于其生长的最适温度。本研究发现,增温能够显著增加禾草和杂草的高度,而对于其他功能群的高度无显著影响,同时能够增加莎草科植物盖度,相关分析表明,莎草科盖度与土壤含水量呈显著正相关,即在藏北高寒草甸,土壤含水量增加,莎草科盖度增加,增温改变了OTC 内的微气候,空气温度、土壤温度、土壤水分、微生物活性都发生了相应的变化,所以植物的生长也会变化。群落的高度与盖度反映的是群落对于空间资源的利用,特别是光资源的竞争,Alward(1999)研究结果也表明,在全球变暖的大背景下,总有一些物种对于温度变化较不敏感,从而引起种间变化,改变植物种组成。增温与增氮处理,对豆科植物均无显著影响,而在交互作用中,豆科植物的高度、盖度和生物量都显著增加,且相关分析表明,豆科生物量与土壤含水量呈显著负相关,即在高寒草甸,土壤含水量下降,豆科生物量增加。
物种多样性能够促进生态系统的稳定性,使整个草地生态系统稳定健康发展,本研究发现在未来增温、氮沉降增加的趋势下,物种多样性呈降低趋势。由于OTC 内温度升高,空气流动性减弱,植物的蒸腾作用增强,植物的生理过程发生改变,同时由于温度的升高,土壤氮矿化速率增加,加上氮沉降的增加,导致氮素可利用性增加,由于不同的植物对于资源竞争力的不同,植物本身应对环境变化的生理过程不同,从而对各物种的生长状况的影响也不尽相同。
4 结论
增氮处理使地上总生物量显著增加,显著提高杂类草的高度和莎草科植物的盖度以及莎草科植物的物种丰富度;增温能够显著提高禾草和杂草的高度;增温增氮处理显著提高豆科植物的高度、盖度和地上生物量、降低莎草科植物的物种丰富度,并使Simpson 多样性指数显著下降7.4%。综合分析,高寒草甸在未来增温、氮沉降增加趋势下,总生物量呈增加趋势,其中豆科植物生物量显著增加,物种多样性降低,莎草科物种丰富度显著下降。由此说明,氮沉降的增加解除了氮素的营养限制,温度的升高解除了植物生长的温度限制,这两者共同促进高寒草甸植物的生长和生物量的积累。温度升高、氮沉降增加,使植物生长可利用资源增加,由于植物对水分、氮素和光的竞争,以及对环境变化的敏感度不同,导致物种多样性降低。
