绿色超级稻品种的农艺与生理性状分析
2019-12-25许阳东朱宽宇章星传王志琴杨建昌
许阳东 朱宽宇 章星传 王志琴 杨建昌
绿色超级稻品种的农艺与生理性状分析
许阳东 朱宽宇 章星传 王志琴 杨建昌*
扬州大学江苏省作物遗传生理国家重点实验室培育点/ 粮食作物现代产业技术协同创新中心, 江苏扬州 225009
探明绿色超级稻的农艺与生理性状, 对于培育和选用绿色超级稻品种有重要意义。本研究以4个绿色超级稻品种为材料, 1个超级稻品种和1个非超级稻品种为对照, 观察了绿色超级稻的农艺与生理表现。结果表明, 与对照品种相比, 绿色超级稻品种具有较高的产量和氮素利用效率。绿色超级稻品种较高的产量得益于总颖花数和结实率的同步提高, 较高的氮素利用率主要在于较高的植株氮素籽粒生产效率(氮素内部利用效率)。绿色超级稻具有较高的茎蘖成穗率和粒叶比, 抽穗期较高的糖花比, 灌浆期较高的作物生长速率、净同化率、根系氧化力和茎中同化物向籽粒的运转率和成熟期较高的收获指数。这些性状与产量及植株氮素籽粒生产效率均呈极显著的正相关。建议将上述性状作为培育和选用绿色超级稻品种的参考指标。
绿色超级稻; 产量; 植株氮素籽粒生产效率; 群体质量; 生理性状
水稻是世界上最主要的粮食作物之一, 为30多亿人口提供了近60%的饮食热量[1-2]。中国是世界上最大的水稻生产国和稻米消费国, 持续提高水稻产量对保障我国乃至世界的粮食安全和人民生活水平具有极其重要的作用[2-3]。长期以来, 我国水稻等主要粮食作物生产以矮秆、抗倒、耐肥品种的培育和应用为基础, 以增加化肥、农药和水资源的用量为手段, 大幅度地提高了单位面积的产量, 使得我国目前的稻谷和小麦等多种农产品产量位居世界第一, 成功地创造了以约占全球9%的耕地, 养活了约占全球22%人口的奇迹[4]。但与此同时也形成了高投入、高产出、高污染、低效益的“三高一低”生产模式, 对社会、经济和环境带来了巨大的压力[4]。针对这一严重问题, 张启发[5-6]提出了绿色超级稻的构思并付诸实践。与目前生产上推广应用的“高投入、高产出”的超级稻相比, 绿色超级稻不但具有高产优质, 而且具备抗多种病虫害、营养高效、抗逆等多种优良性状[6-7]。彭少兵[8]认为, 相对于超级稻的中等分蘖力、大穗、高大株型、高叶面积指数、高干物质生产等株型和生理特征, 绿色超级稻则具有高分蘖力、多穗、中等株高, 高干物质转运效率、高结实率、高收获指数等性状。但绿色超级稻的这些株型和生理性状是否具有普遍性?缺乏论证; 物质转运率、结实率和收获指数等性状是最终收获后的结果, 绿色超级稻在生长期间有哪些重要诊断指标或农艺生理性状?缺乏研究。阐明这些问题, 对于绿色超级稻品种的培育和筛选具有重要指导意义。鉴此, 本研究较为系统地比较分析了绿色超级稻、超级稻及非超级稻水稻品种的农艺与生理性状, 旨为绿色超级稻品种的培育和选用提供依据。
1 材料与方法
1.1 材料与试验设计
试验于2016年和2017年在江苏省里下河地区农业科学研究所夏桥基地进行。试验地前茬作物为小麦, 土壤为沙壤土, 耕作层含有机质21.2 g kg–1、有效氮101 mg kg–1、速效磷21.5 mg kg–1、速效钾68.6 mg kg–1。选用能反映当地平均水稻产量水平, 且已通过农业部认定的超级稻品种扬粳4038和通过江苏省审定的非超级稻品种宁粳1号为对照品种, 以符合绿色超级稻特征的武运粳24、连粳7号、淮稻13和武运粳30作为绿色超级稻品种。武运粳24和连粳7号从超级稻品种中选出, 淮稻13和武运粳30从江苏省审定品种中选出。这4个绿色超级稻品种在2014和2015年的试验中, 在200 kg hm–2施氮量条件下, 武运粳24和连粳7号的产量能较对照品种扬粳4038的氮肥利用率(产量/施氮量)提高5%以上, 淮稻13和武运粳30的氮肥利用率比对照品种宁粳1号提高10%以上, 符合国家“863”计划“绿色超级稻新品种选育(2014AA10A600)”项目组制定的绿色超级稻品种鉴定标准(目前仅在项目组内部试行)。各品种5月17日至18日播种, 6月14日至15日移栽。株、行距为15 cm × 25 cm, 每穴3株苗, 小区面积30 m2, 随机区组排列, 重复3次。小区之间筑埂并用塑料薄膜包裹。各小区全生育期施用尿素(含氮量46%)折合成纯氮200 kg hm–2, 按基肥(移栽前1 d)∶分蘖肥(移栽后7 d)∶穗肥(叶龄余数2.0) = 4∶2∶4施用。在移栽前施过磷酸钙(含P2O513%) 300 kg hm–2。施氯化钾(含K2O 62%) 177 kg hm–2, 按6∶4分基肥和拔节肥(移栽后36~37 d) 2次施用。参照Yang等[9]方法全生育期轻干湿交替灌溉, 严格控制杂草和病虫害。各供试品种于8月25日至27日抽穗, 10月14日至15日收获。
1.2 取样与测定
1.2.1 茎蘖动态、干物质重、非结构性碳水化合物(NSC)和植株含氮量 移栽后各小区定点20穴, 每7 d观察一次茎蘖数, 直至茎蘖数稳定。分别于分蘖期(移栽后15 d)、拔节期(茎基部第1节间开始伸长)、抽穗期(50%的植株抽穗)和成熟期(收获前1 d), 各小区从离边行的第3行开始, 取生长均匀并有代表性的稻株5穴, 用叶面积仪(LI-3050C, Li-Cor, Licin, NE, USA)测定绿色叶片的叶面积, 将叶片(绿叶+黄叶)、茎+鞘、穗(抽穗期和成熟期)分开并烘干至恒重后测定植株干重。在抽穗期和成熟期取样测定的植株样品中, 取出部分样品经粉碎后参照Pucher等[10]的方法测定茎与鞘中的NSC (淀粉和可溶性糖)含量。用凯氏定氮法测定成熟期植株样品中的含氮量[11]。
1.2.2 根干重和根系氧化力 分别于抽穗期, 灌浆前期(抽穗后10 d)、灌浆中期(抽穗后25 d)和灌浆后期(抽穗后38 d), 各小区取代表性植株3穴(每穴以稻株基部为中心, 挖取25 cm × 15 cm × 20 cm的土块), 装于70目的筛网袋中, 先用流水再用农用压缩喷雾器将根冲洗干净, 称取根鲜重, 取部分根按Zhang等方法[12]测定根系氧化力, 其余根烘干后测定根干重。
1.2.3 考种计产 于成熟期各小区取30穴考察实际穗数, 取10穴测定每穗颖花数、结实率与千粒重, 结实率采用水漂法测定, 将稻谷置于密度为1.1×103kg m–3的生理盐水中, 沉入水底则是饱粒, 其余为空粒或瘪粒。每小区实收5 m2稻株脱粒计产。
1.3 数据分析与计算
用SAS/STAT统计软件(Version 9.2, SAS Institute, Cray, NC, USA)进行方差分析、显著性检验和相关等统计分析。常见生理指标按下式计算:
茎蘖成穗率(%) = 成熟期有效穗数(个 m–2)/拔节期最高茎蘖数(个 m–2) × 100 (1)
糖花比(mg spikelet–1) = 抽穗期茎与鞘中NSC量(mg m–2)/颖花数(spikelets m–2) (2)
NSC转运率(%) = [抽穗期茎与鞘中NSC积累量(g m–2)-成熟期茎与鞘中NSC积累量(g m–2)]/抽穗期茎与鞘中NSC积累量(g m–2) × 100 (3)
颖花粒叶比(spikelets per leaf area, cm2) = 颖花数(spikelets m–2)/抽穗期叶面积(cm2m–2) (4)
实粒粒叶比(filled grains per leaf area, cm2) = 实粒数(grains m–2)/抽穗期叶面积(cm2m–2) (5)
粒重粒叶比[grain weight (mg) per leaf area (cm2)] = 总粒重(mg m–2)/抽穗期叶面积(cm2m–2) (6)
绿叶面积持续期(m2m–2d) = (1+2)/2 × (2-1) (7)
作物生长速率(g m–2d–1) = (2-1)/(2-1) (8)
净同化速率(g m–2d–1) = (2-1)/ [(2-1) × (2-1)] (9)
在式(7)、(8)或(9)中,1和2分别为第1次和第2次测定的绿叶面积(m2m–2),1和2分别为第1次和第2次测定的时间(d)),1和2分别为第1次和第2次测定的地上部植株干重(g m–2)。
植株氮素籽粒生产效率(internal nitrogen use efficiency, kg kg–1N, IEN) = 产量(kg hm–2)/成熟期植株吸氮量(kg hm–2) (10)
每百kg籽粒需氮量(kg per 100 kg) = 成熟期植株吸氮量(kg hm–2)/产量(kg hm–2) ×100 (11)
氮肥偏生产力(kg kg–1N) = 产量(kg hm–2)/施氮量(kg hm–2) (12)
收获指数= 产量(干重, kg m–2)/成熟期地上部植株干重(kg m–2) ×100 (13)
2 结果与分析
2.1 产量和氮肥利用率
4个绿色超级稻品种的产量较两个对照品种的产量显著增加(<0.05), 平均较超级稻品种(扬粳4038, CK1)增产9.56%, 平均较非超级稻品种(宁粳1号, CK2)增产12.03% (表1)。在4个超级稻品种间以及在2个对照品种间的产量差异不显著。从产量构成因素分析, 绿色超级稻品种增产的原因主要是总颖花数和结实率的同步提高, 而总颖花数的提高在于单位面积穗数或每穗颖花数的增加, 千粒重在品种间差异较小(表1)。

表1 绿色超级稻品种的产量及其构成因素
同栏内不同字母者表示在= 0.05水平上差异显著;*,**分别表示在= 0.05和= 0.01水平上差异显著; NS: 在= 0.05水平上差异不显著。
Values in the same column followed by different letters are significant at= 0.05.*,**-values are significant at= 0.05 and= 0.01, respectively. NS means not significant at= 0.05. YJ-4: Yangjing 4038; WYJ-24: Wuyunjing 24; LJ-7: Lianjing 7; NJ-1: Ningjing 1; HD-13: Huaidao 13: WYJ-30: Wuyunjing 30.
与产量结果十分类似, 绿色超级稻品种的植株氮素籽粒生产效率和氮肥偏生产力显著高于对照品种(表2)。绿色超级稻品种植株氮素籽粒生产效率的提高主要在于每千克籽粒需氮量的减少, 成熟期植株总吸氮量在绿色超级稻品种与对照品种间无显著差异(表2), 表明绿色超级稻品种提高了植株吸收单位氮素的生产籽粒的效率。
2.2 群体质量和生理性状
2.2.1 茎蘖成穗率、叶面积和粒叶比 4个绿色超级稻品种的茎蘖成穗率较2个对照品种高出4.5~9.6个百分点, 差异显著(表3)。抽穗期叶面积指数(LAI)在各供试品种间无显著差异, 但粒叶比的3个指标(颖花/叶面积、实粒/叶面积和总粒重/叶面积), 绿色超级稻品种均显著高于对照品种(表3), 说明绿色超级稻的库源比较高。
2.2.2 作物生长速率、叶片净同化速率和绿叶面积持续期 干物质重在品种间的差异因生育期不同而异(表4)。在分蘖期和成熟期, 绿色超级稻品种武运粳24、淮稻13和武运粳30的地上部植株干重显著高于2个对照品种, 绿色超级稻品种连粳7号与2个对照品种差异不显著; 在拔节期和抽穗期, 植株干重在各供试品种间差异均不显著。拔节至抽穗期作物生长速率在各品种间无显著差异, 但分蘖至拔节及抽穗至成熟, 绿色超级稻品种显著高于对照品种(表4), 说明绿色超级稻品种在生长前期和后期具有较高的物质生产优势。
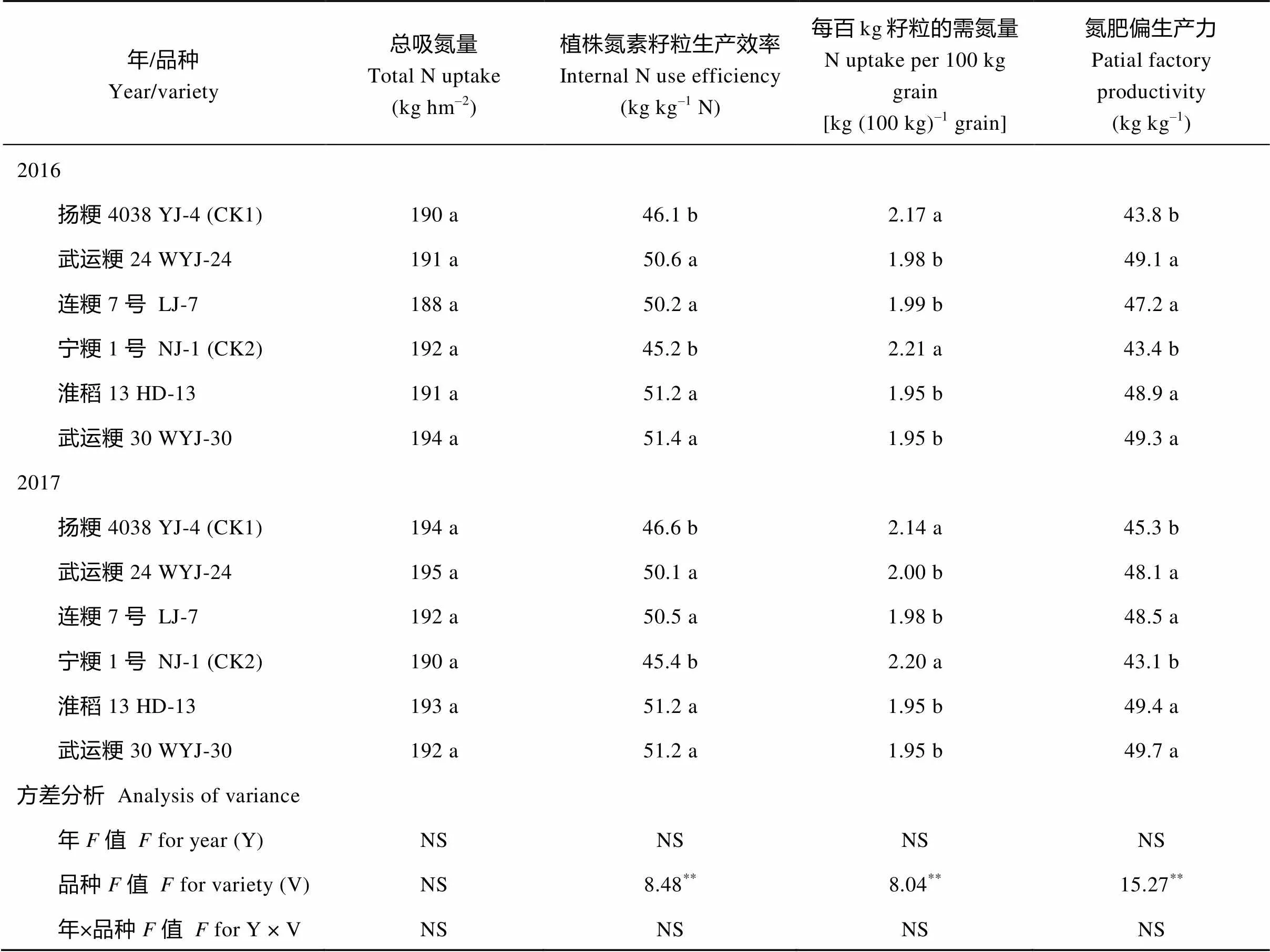
表2 绿色超级稻品种的氮素吸收量和氮利用率
同栏内不同字母者表示在=0.05水平上差异显著;**表示在=0.01水平上差异显著; NS: 在=0.05水平上差异不显著。
Values in the same column followed by different letters are significant at=0.05.**-values are significant at= 0.01. NS means not significant at= 0.05. YJ-4: Yangjing 4038; WYJ-24: Wuyunjing 24; LJ-7: Lianjing 7; NJ-1: Ningjing 1; HD-13: Huaidao 13: WYJ-30: Wuyunjing 30.

表3 绿色超级稻品种茎蘖成穗率(PPST)、叶面积指数(LAI)和粒叶比
同栏内不同字母者表示在= 0.05水平上差异显著;**表示在= 0.01水平上差异显著; NS: 在= 0.05水平上差异不显著。
Values in the same column followed by different letters are significant at= 0.05.**-values are significant at= 0.01. NS means not significant at=0.05. YJ-4: Yangjing 4038; WYJ-24: Wuyunjing 24; LJ-7: Lianjing 7; NJ-1: Ningjing 1; HD-13: Huaidao 13: WYJ-30: Wuyunjing 30.

表4 绿色超级稻品种的地上部分干重和作物生长速率(CGR)
同栏内不同字母者表示在= 0.05水平上差异显著;*,**分别表示在= 0.05和= 0.01水平上差异显著; NS: 在= 0.05水平上差异不显著。
Values in the same column followed by different letters are significant at= 0.05.*,**-values are significant at= 0.05 and= 0.01, respectively. NS means not significant at the=0.05. YJ-4: Yangjing 4038; WYJ-24: Wuyunjing 24; LJ-7: Lianjing 7; NJ-1: Ningjing 1; HD-13: Huaidao 13: WYJ-30: Wuyunjing 30.
与作物生长速率的结果趋势相一致, 拔节至抽穗期的叶片净同化速率在各品种间无显著差异, 但分蘖至拔节及抽穗至成熟这两个阶段的叶片净同化速率, 绿色超级稻品种显著高于对照品种(表5)。各生育阶段的绿叶面积持续期在各品种间均无显著差异(表5)。说明绿色超级稻分蘖至拔节和抽穗至成熟这两个阶段的作物生长速率大, 主要是提高了此两生育时期源的质量(单位叶面积净同化速率)。

表5 绿色超级稻品种绿叶面积持续期和净同化速率
同栏内不同字母者表示在= 0.05水平上差异显著;**表示在= 0.01水平上差异显著; NS: 在= 0.05水平上差异不显著。
Values in the same column followed by different letters are significant at= 0.05.**-values are significant at= 0.01. NS means not significant at= 0.05. T–J: tillering–jointing; J–H: jointing–heading; H–M: heading–maturity; YJ-4: Yangjing 4038; WYJ-24: Wuyunjing 24; LJ-7: Lianjing 7; NJ-1: Ningjing 1; HD-13: Huaidao 13: WYJ-30: Wuyunjing 30.
2.2.3 糖花比、物质转运和收获指数 抽穗期茎与鞘中非结构性碳水化合物(NSC)量, 4个绿色超级稻品种显著高于2个对照品种, 前者较后者平均高出20.8% (表6)。抽穗期的糖花比(NSC量/颖花数), 超级稻品种显著高于对照品种(表6)。在成熟期, 茎与鞘中的NSC在超级稻品种与对照品种间差异较小, 因而抽穗至成熟期茎与鞘中NSC的转运率, 超级稻品种显著高于对照品种(表6)。与NSC的转运率结果趋势一致, 绿色超级稻品种较对照品种显著提高了收获指数(表6)。说明绿色超级稻具有较高的物质生产效率。
2.2.4 根重、根冠比与根系活力 抽穗期的根干重和根冠比在各品种间均无显著差异(图1-A~D), 灌浆前期根系氧化力在品种的差异也不显著(图1-E, F)。但在灌浆中期和后期的根系氧化力, 4个超级稻品种显著高于对照品种(图1-E, F)。相关分析表明, 灌浆期的平均根系氧化力与灌浆期作物生长速率和净同化率以及成熟期的结实率均呈极显著的正相关(图2-A~C), 说明灌浆期根系活力高是绿色超级稻灌浆结实期物质生产能力强和结实率高的一个重要原因。
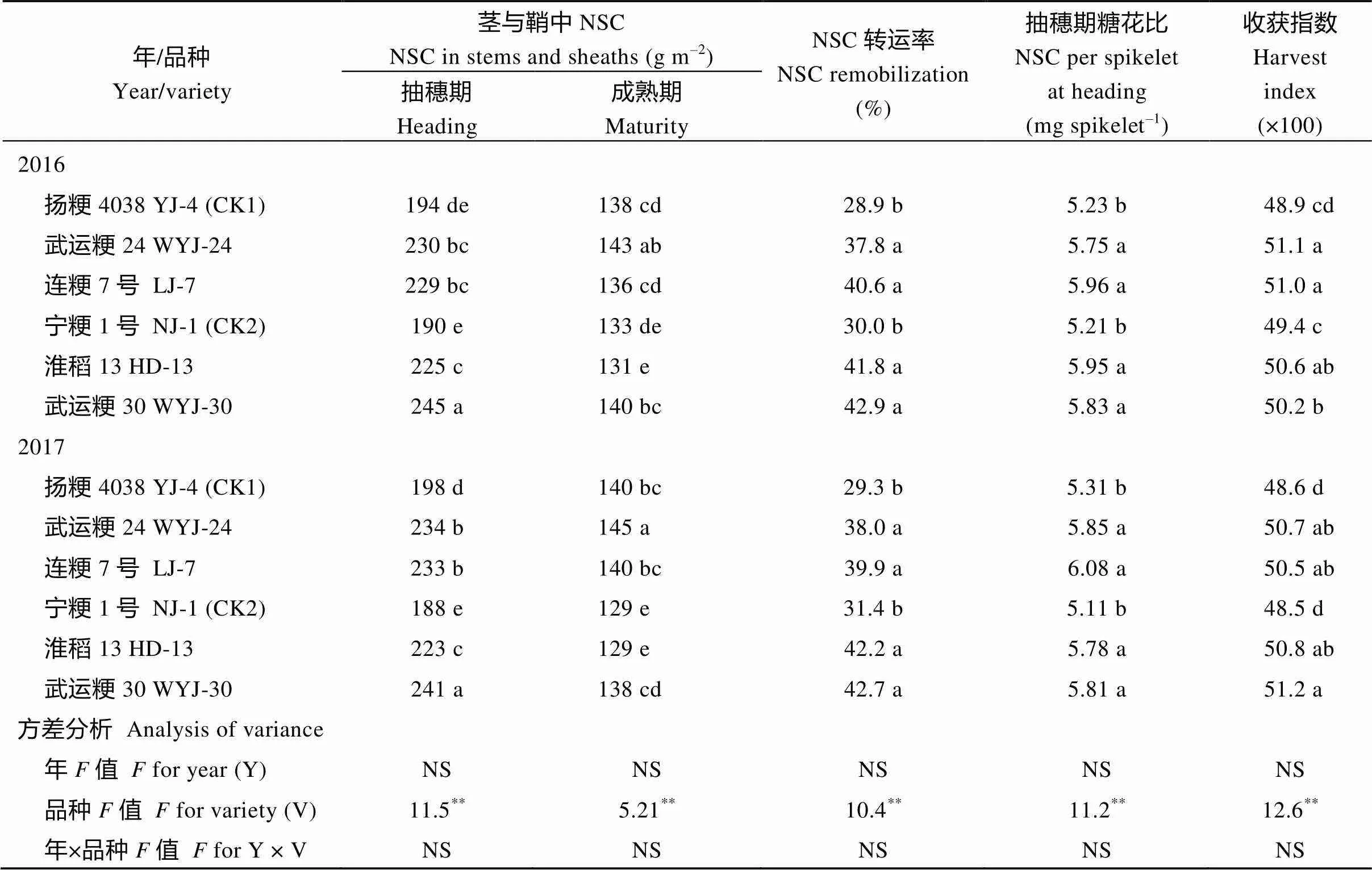
表6 绿色超级稻茎中非结构性碳水化合物(NSC)转运率、糖花比和收获指数
同栏内不同字母者表示在= 0.05水平上差异显著;**表示在= 0.01水平上差异显著; NS: 在= 0.05水平上差异不显著。
Values in the same column followed by different letters are significant at= 0.05.**-values are significant at= 0.01. NS means not significant at= 0.05. YJ-4: Yangjing 4038; WYJ-24: Wuyunjing 24; LJ-7: Lianjing 7; NJ-1: Ningjing 1; HD-13: Huaidao 13: WYJ-30: Wuyunjing 30.
(图1)
不同字母者表示在= 0.05水平上差异显著, NS表示在0.05水平上差异不显著, 同一测定时期、不同品种间比较。YJ-4: 扬粳4038; WYJ-24: 武运粳24; LJ-7: 连粳7号; NJ-1: 宁粳1号; HD-13: 淮稻13; WYJ-30: 武运粳30; EGF: 灌浆早期; MGF: 灌浆中期; LGF: 灌浆后期。
Values in the figure are expressed as means±standard error (= 3). Different letters on the column indicate statistical significance at= 0.05 within the same measurement date. NS means not significant at= 0.05. YJ-4: Yangjing 4038; WYJ-24: Wuyunjing 24; LJ-7: Lianjing 7; NJ-1: Ningjing 1; HD-13: Huaidao 13: WYJ-30: Wuyunjing 30; EGF: early grain filling; MGF: mid grain filling; LGF: late grain filling.

图2 水稻灌浆期根系氧化力(ROA)与灌浆期作物生长速率(A)、灌浆期净同化率(B)和结实率(C)的关系
图中数据来自表1、表4、表5和图1;**表示在= 0.01水平上显著(= 12)
Values are calculated from the data from Tables 1, 4, and 5 and Figure 1;**Significant at= 0.01 (= 12).
2.3 植株生理性状与产量及氮肥利用率的关系
表7为测定的主要生理性状与产量及氮肥利用率的相关分析。由表可知, 茎蘖成穗率、抽穗期糖花比、粒叶比、灌浆期作物生长速率、灌浆期净同化率、灌浆期根系氧化力、茎中同化物转运率及收获指数与产量、植株氮素籽粒生产效率均呈极显著的正相关(= 0.85**~0.99**) (表7)。说明改善上述性状, 有利于提高产量和氮肥利用效率。
3 讨论
彭少兵[8]指出, 绿色超级稻品种应具有高物质转运效率, 高结实率和高收获指数等生理性状。本研究结果表明, 绿色超级稻品种抽穗至成熟期茎鞘中NSC的转运率、结实率和收获指数均显著高于对照品种(超级稻品种或非超级稻品种), 证实了彭少兵的观点。但本研究还发现, 绿色超级稻品种具有较高的茎蘖成穗率和较高的粒叶比, 抽穗期较高的糖花比, 灌浆期较高的作物生长速率、净同化率和根系氧化力, 上述性状与产量及植株氮素籽粒生产效率均成极显著的正相关, 说明高茎蘖成穗率, 高粒叶比, 高糖花比, 灌浆期高作物生长速率、高净同化率和高根系氧化力也是绿色超级稻的重要生理性状, 建议将这些性状作为培育和选用绿色超级稻品种的参考指标。
为什么茎蘖成穗率、糖花比、粒叶比等性状与产量及氮肥利用率密切相关, 其生理机制是什么? 本研究虽未对此问题进行深入探讨, 但根据以往的研究结果可以总结出以下几点: 第一, 茎蘖成穗率高, 表明无效分蘖就少, 用于无效茎、蘖、叶生长的水分养分消耗就少[13-14]。无效分蘖的减少不仅有利于改善群体通风透光条件, 而且有利于改善冠层结构, 进而有利于抽穗后物质生产和积累[14-15]。第二, 抽穗期糖花比高, 表明抽穗前茎与鞘中同花物(NSC)累积量大, 每朵颖花获得的NSC多, 不仅有利于抽穗前花粉粒的充实完成, 而且可以增加抽穗至成熟期茎中同花物向籽粒的运转量, 促进花后胚乳细胞的发育和籽粒的充实[16-17]。第三, 粒叶比高, 表明库容相对较大, 叶源相对较小。Yang等[18]研究表明, 水稻籽粒库对叶片光合和同化物转运有促进作用。在群体颖花量或叶面积指数接近条件下, 凡粒叶比越高的群体, 叶片的光合速率就越高[19-20]。不仅如此, 水稻颖花等经济器官并不是被动接纳产量物质的器官, 它具有主动向光合生产系统“提取”光合常产物的能力[20-21], 因此, 粒叶比高, 抽穗至成熟期茎中同花物向籽粒的运转速度就越快, 运转量就越大[19-23]。我们认为, 高茎蘖成穗率、高糖花比和高粒叶比是绿色超级稻品种高物质转运效率, 高结实率和高收获指数的重要生理基础。

表7 水稻部分农艺生理性状与产量及植株氮素籽粒生产效率(IEN)的相关系数(r)
表中数据来自表1至表6和图1; IEN:植株氮素籽粒生产效率;**表示在= 0.01水平上显著(= 12)。
Values are calculated from the data of Tables 1 to 6 and Figure 1; IEN: internal nitrogen use efficiency;**Significant at= 0.01 (= 12).
一般认为, 水稻的颖花数与结实率呈负相关[24-25]。但本研究表明, 与对照品种相比, 绿色超级稻品种不仅总颖花量较多, 而且结实率也较高。分析两者协同提高的原因, 除了高的糖花比有利于籽粒灌浆充实外, 绿色超级稻灌浆期具有较高的根系氧化力和较高作物生长速率也是重要因素。根系氧化力是根系活性的一个重要指标[26-27]。抽穗至成熟期特别是灌浆中后期根系活性强有利于提高地上部叶片净光合速率和延长叶片光合作用时间, 提高花后干物质生产[28-29]。另一方面, 较高的地上部光合生产能力又为根系提供较多的光合同化物, 保证了根系生长和活力。两者协同的作用确保了绿色超级稻实现总颖花量和结实率的同步提高。
扩大产量库容(增加总粒数或总颖花数)是实现水稻高产的主要途径[30-31]。扩大库容可以通过: 1) 增加穗数; 2) 增加每穗粒数; 3) 两者兼之。彭少兵[8]认为,相对于超级稻的大穗, 绿色超级稻则具有多穗。本研究结果表明, 绿色超级稻品种的总颖花量高, 或是每穗颖花数多, 或是单位地面积的穗数多。说明水稻增产途径因品种或环境不同而异, 增加每穗颖花数或增加穗数均是绿色超级稻丰产的重要途径。
本研究观察到, 超级稻扬粳4038在施氮量为200 kg hm–2条件下的产量仅为8.9 t hm–2, 低于其他超级稻品种。Ju等[32]也观察到, 当施氮量为200 kg hm–2时, 扬粳4038的产量仅为8.52 t hm–2, 当施氮量为300 kg hm–2时, 该品种的产量为9.88 t hm–2。说明超级稻扬粳4038适合于高施氮量( >200 kg hm–2), 在较低施氮量( ≤ 200 kg hm–2)下没有产量优势, 同时也说明该品种对氮素响应较为钝感。
本研究发现, 绿色超级稻的氮肥利用率高, 不是它的吸收量高, 而是吸收单位氮素生产籽粒的效率高(植株氮素籽粒生产效率高)或生产每千克籽粒的需氮量少。为什么绿色超级稻品种吸收单位氮素生产籽粒的效率高?从本研究的结果分析, 较高的茎蘖成穗率, 较高的物质转运效率, 较高的结实率和较高收获指数均是植株氮素籽粒生产效率高的重要原因[32-33]。有关绿色超级稻品种植株氮素籽粒生产效率高的机制, 有待深入研究。
4 结论
与超级稻品种或非超级稻品种相比, 绿色超级稻品种具有较高的产量和氮素利用效率。绿色超级稻品种较高的产量得益于总颖花数和结实率的同步提高, 较高的氮素利用率主要在于较高的植株氮素籽粒生产效率。较高的茎蘖成穗率和粒叶比, 抽穗期较高的糖花比, 灌浆期较高的作物生长速率、净同化率、根系氧化力和茎中同化物向籽粒的运转率以及成熟期较高的收获指数是绿色超级稻品种高产与氮高效利用的重要生理基础, 上述性状可作为培育和选用绿色超级稻品种的参考指标。
[1] FAOSTAT. FAO Statistical Databases, Food and Agriculture Organization (FAO) of the United Nations, Rome, 2016. http://www.fao.org/.
[2] 余四斌, 熊银, 肖景华, 罗利军, 张启发. 杂交稻与绿色超级稻. 科学通报, 2016, 61: 3797−3803. Yu S B, Xiong Y, Xiao J H, Luo L J, Zhang Q F. Hybrid rice and green super rice., 2016, 61: 3797−3803 (in Chinese with English abstract).
[3] Peng S B, Tang Q Y, Zou Y B. Current status and challenges of rice production in China.,2009, 12: 3−8.
[4] 张启发. 资源节约型、环境友好型农业生产体系的理论与实践. 北京: 科学出版社, 2015. pp 1−15. Zhang Q F. Theory and Practice of Resource-saving and Environment-friendly Agricultural Production System. Beijing: Science Press, 2015. pp 1−15 (in Chinese).
[5] Zhang Q F. Strategies for developing green super rice., 2007, 104: 16402–16409.
[6] 张启发. 绿色超级稻的构想与实践. 北京: 科学出版社, 2010. pp 1−32. Zhang Q F. Strategies and Practice for Developing Green Super Rice. Beijing: Science Press, 2010. pp 1−32 (in Chinese).
[7] Wang F, Peng S B. Yield potential and nitrogen use efficiency of China’s super rice., 2017, 16: 1000−1008.
[8] 彭少兵. 对转型时期水稻生产的战略思考. 中国科学: 生命科学, 2014, 44: 845−850. Peng S B. Reflection on China’s rice production strategies during the transition period., 2014, 44: 845–850 (in Chinese with English abstract).
[9] Yang J C, Liu K, Wang Z Q, Du Y, Zhang J H. Water-saving and high-yielding irrigation for lowland rice by controlling limiting values of soil water potential., 2007, 49: 1445−1454.
[10] Pucher G W, Leavenworth C S, Vickery H B. Determination of starch in plant tissues., 1948, 20: 850−853.
[11] Yoshida S, Forno D, Cock J, Gomez K. Laboratory Manual for Physiological Studies of Rice. The Philippines: International Rice Research Institute, 1976. pp 14−20.
[12] Zhang H, Xue Y G, Wang Z Q, Yang J C, Zhang J H. An alternate wetting and moderate soil drying regime improves root and shoot growth in rice., 2009,49: 2246−2260.
[13] Yang J C, Zhang J H. Crop management techniques to enhance harvest index in rice., 2010, 61: 3177−3189.
[14] Wang Z Q, Zhang W Y, Beebout S S, Zhang H, Liu L J, Yang J C, Zhang J H. Grain yield, water and nitrogen use efficiencies of rice as influenced by irrigation regimes and their interaction with nitrogen rates., 2016, 193: 54−69.
[15] 张自常, 徐云姬, 褚光, 王志琴, 王学明, 刘立军, 杨建昌. 不同灌溉方式下的水稻群体质量. 作物学报, 2011, 37: 2011−2019. Zhang Z C, Xu Y J, Chu G, Wang Z Q, Wang X M, Liu L J, Yang J C. Population quality of rice under different irrigation regimes., 2011, 37: 2011−2019 (in Chinese with English abstract).
[16] Fu J, Huang Z H, Wang Z Q, Yang J C, Zhang J H. Pre-anthesis non-structural carbohydrate reserve in the stem enhances the sink strength of inferior spikelets during grain filling of rice., 2011, 123: 170−182.
[17] Yang J C,Peng S B, Zhang Z J, Wang Z Q, Visperas R M, Zhu Q S. Grain and dry matter yields and partitioning of assimilates inhybrid rice., 2002, 42: 766−772.
[18] Yang J C, Zhang J H, Wang Z Q, Zhu Q S. Hormones in the grains in relation to sink strength and postanthesis development of spikelets in rice., 2003, 41: 185−195.
[19] 凌启鸿, 杨建昌. 水稻群体“粒叶比”与高产栽培途径研究. 中国农业科学, 1986, 19(3): 1−8. Ling Q H, Yang J C. Studies on “grain-leaf ratio” of population and cultural approaches of high yield in rice plants., 1986, 19(3): 1−8 (in Chinese with English abstract).
[20] 杨建昌. 水稻粒叶比与产量的关系. 江苏农学院学报, 1993, 14(专刊): 11−14. Yang J C. Relationship between grain-leaf ratio and grain yield of rice., 1993, 14(Special issue): 11−14 (in Chinese with English abstract).
[21] 凌启鸿. 作物群体质量. 上海: 上海科学技术出版社, 2000. pp 42−216. Ling Q H. Crop population Quality. Shanghai: Shanghai Scientific & Technical Publishers, 2000. pp 42−216 (in Chinese).
[22] Yang J C. Approaches to achieve high yield and high resource use efficiency in rice., 2015, 2: 115−123.
[23] Li H W, Liu L J, Wang Z Q, Yang J C, Zhang J H. Agronomic and physiological performance of high-yielding wheat and rice in the lower reaches of Yangtze River of China., 2012, 133: 119−129.
[24] Kato T, Takeda K. Associations among characters related to yield sink capacity in space-planted rice., 1996, 36: 1135−1139.
[25] Fageria N K.Yield physiology of rice., 2007, 30: 843−879.
[26] Yang J C, Zhang H, Zhang J H. Root morphology and physiology in relation to the yield formation of rice., 2012, 11: 920−926.
[27] Chu G, Chen T T, Wang Z Q, Yang J C, Zhang J H. Morphological and physiological traits of roots and their relationships with water productivity in water-saving and drought-resistant rice., 2014, 162:108−119.
[28] Osaki M, Shinano T, Matsumoto M, Zheng T, Tadano T. A root-shoot interaction hypothesis for high productivity of field crops., 1997, 37: 445−454.
[29] Samejima H, Kondo M, Ito O, Nozoe T, Shinano T, Osaki M. Root-shoot interaction as a limiting factor of biomass productivity in new tropical rice lines., 2004, 50: 545−554.
[30] Yang J C, Zhang J H. Grain-filling problem in “super” rice., 2010, 61: 1−5.
[31] Zhang Z J, Chu G, Liu L J, Wang Z Q, Wang X M, Zhang H, Yang J C, Zhang J H. Mid-season nitrogen application strategies for rice varieties differing in panicle size., 2013, 150: 9−18.
[32] Ju C X, Buresh R J, Wang Z Q, Zhang H, Liu L J, Yang J C, Zhang J H. Root and shoot traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates application., 2015, 175: 47−59.
[33] Chu G, Wang Z Q, Zhang H, Yang J C, Zhang J H. Agronomic and physiological performance of rice under integrative crop management., 2016, 108: 117−128.
Analysis in agronomic and physiological traits of green super rice
XU Yang-Dong, ZHU Kuan-Yu, ZHANG Xing-Chuan, WANG Zhi-Qin, and YANG Jian-Chang*
Jiangsu Key Laboratory of Crop Genetics and Physiology / Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou 225009, Jiangsu, China
Understanding agronomic and physiological characteristics of green super rice (GSR) is essential to make strategies for breeding GSR varieties and crop management. This study aimed to identify major agronomic and physiological traits associated with high grain yield and high nitrogen use efficiency (NUE) of rice. Four GSR varieties and two control varieties (one super rice variety and one non-super rice inbred) were grown in the paddy field. The results showed that the GSR varieties produced higher grain yield and higher NUE than control varieties. Synchronous increases in both total spikelet number and filled-grain percentage contributed to higher grain yield, and an increase in grain yield production per unit absorbed N (internal NUE) resulted in higher NUE for GSR. Compared with control varieties, GSR varieties exhibited greater percentage of productive tillers and the ratio of spikelet number to leaf area, higher ratio of sugar amount to spikelet number at the heading time, higher crop growth rate, net assimilate rate, and root oxidation activity and more remobilization of assimilates from stems and sheaths to grains during the grain filling period, and higher harvest index at maturity. These traits were positively and very significantly correlated with both grain yield and internal NUE, and could be used as indexes for breeding and selecting GSR varieties.
green super rice (GSR); grain yield; internal nitrogen use efficiency; population quality; physiological trait
2018-07-08;
2018-10-08;
2018-11-08.
10.3724/SP.J.1006.2019.82036
通信作者(Corresponding author): 杨建昌, E-mail: jcyang@yzu.edu.cn
E-mail: 1376959876@qq.com
本研究由国家高技术研究发展计划项目(863计划)(2014AA10A600), 国家自然科学基金项目(31461143015), 江苏高校优势学科建设工程项目(PAPD)和扬州大学高端人才支持计划项目(2015-1)资助。
This study was supported by the National High Technology Research and Development Program of China (2014AA10A600), the National Natural Science Foundation of China (31461143015), the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the Top Talent Supporting Program of Yangzhou University (2015-01).
URL:http://kns.cnki.net/kcms/detail/11.1809.S.20181105.0941.006.html
