咖啡种植模式对蚂蚁多样性的影响
2019-12-25于潇雨卢志兴陈又清
于潇雨,卢志兴,李 巧①,燕 迪,陈又清
(1.西南林业大学生物多样性保护学院,云南 昆明 650224; 2.中国林业科学研究院资源昆虫研究所,云南 昆明 650224)
咖啡(Coffeaarabica)是世界四大饮料作物之一,云南省是中国咖啡种植的主要省份,种植面积及产量均占我国咖啡种植总量的98%以上,近年国家着力打造有机咖啡园,推进绿色种植[1]。云南主要种植的小粒咖啡原产于埃塞俄比亚的热带地区,喜荫蔽、温凉、湿润环境[2],但云南的小粒咖啡已被扩展到气候最适宜区以外,例如元阳、墨江、金平[3]。传统的小粒咖啡生长于荫蔽树下[4],按照此模式与遮荫树种混合种植的咖啡被称为荫蔽咖啡(shade coffee)。研究认为,无遮荫的阳光咖啡(sun coffee)即咖啡单作模式,存在土地侵蚀和生物多样性丧失的问题[5-6],且需要花费大量化肥、杀虫剂等经济成本[7]。在大力发展咖啡种植过程中,应考虑保护当地生物多样性,打造有机咖啡种植园。我国咖啡的种植模式较多,其中荫蔽咖啡复合农林内的遮荫树种也有所不同。有的使用本地树种作为遮荫树,如龙眼(Dimocarpuslongan)[8],也有的使用外来经济作物,例如橡胶(Heveabrasiliensis)[9]。外来经济林的种植园被认为有降低和取代现有生态系统的作用[10]。由于咖啡本就是外来树种,再使用外来植物作为遮荫树种,这样的复合农林系统可能会对当地生态系统服务与生物多样性造成威胁[11]。持续的集约化生产将降低农业生态系统中的动物多样性[12],国外研究指出咖啡集约化将会降低节肢动物多样性[13],但我国对咖啡园内生物多样性的研究还较少。
蚂蚁是包括咖啡种植园在内的很多农业生态系统中有效的生物防治类群[14]。蚂蚁作为重要捕食者,在农业系统中可通过直接捕食害虫,或驱赶和惊扰害虫从寄主植物上掉落,间接抑制植食性节肢动物群落发展[15-16]。同时蚂蚁也能通过除去孢子来减少真菌植物病原体,或限制植物与病菌媒介之间的相互作用来控制病害[17-18]。另外,蚂蚁作为一类有效的生物指示物,能敏感地对栖境变化做出响应[19],其多度和群落结构参数可用来反映环境变化与人为干扰程度[20],也能为生物多样性的恢复提供科学的度量标准[21]。
云南省普洱市是重要的咖啡原料生产基地,咖啡种植面积广,阳光咖啡与荫蔽咖啡2种种植模式均有,但关于其生物多样性的研究却较少。荫蔽咖啡种植模式下,使用本地树种钝叶黄檀和外来树种橡胶作为遮荫树种的情况较多,但前人研究较少关注不同遮荫树种对咖啡种植园生物多样性的影响[21-22]。同时,咖啡与橡胶2种外来树种混种是否会对当地生物多样性造成更大的负担,选择乡土还是外来树种作为遮荫树更好,是值得探讨的问题。该研究选取3种不同类型咖啡种植样地,通过调查不同模式下的蚂蚁多样性、群落结构和指示物种,探讨以下问题:(1)咖啡单作对当地生物多样性有无影响?(2)荫蔽咖啡是否有利于当地蚂蚁多样性保护?(3)用本地或外来树种作为咖啡遮荫树种对当地的生物多样性有何影响?
1 材料与方法
1.1 研究地概况
研究地位于云南省普洱市墨江县雅邑乡座细村(23°13′56″~23°15′51″ N,101°43′14″~101°45′05″ E),海拔为850~1 300 m。该地区属于西南山地季风气候区,年均温21.5 ℃,年均降水量1 450 mm,主要栽培茶叶、咖啡、核桃、玉米等经济作物,另外还使用钝叶黄檀等寄主植物通过放养紫胶虫来生产紫胶。
1.2 样地设置
选择当地4种不同农林生态系统作为4种类型种植样地,分别为钝叶黄檀-玉米混农林(DY)、咖啡纯林(K)、钝叶黄檀-咖啡混农林(DK)和橡胶-咖啡混农林(XK)。其中钝叶黄檀为本地遮荫树种,橡胶为外来遮荫树种。钝叶黄檀-玉米混农林为当地常见且面积最大的土地利用类型,研究选取曾放养过紫胶虫的DY样地作为对照样地。其余样地信息详见表1。
每种类型种植模式下各选择2个调查小区,每个小区面积至少0.5 hm2,小区间距1 km以上,小区上设置样带,为保证样带间的独立性,间距至少50 m。所选小区均在南坡,土壤和坡度条件基本一致。各种植模式样地周围均种有经济作物,无空置撂荒地。另外,为减少边缘效应对实验结果的影响,各种植模式样地均距离道路及其他人为扰动生境50 m以上[23]。
1.3 调查方法
由于云南当地咖啡生产的关键期为每年8—9月,正值当地雨季,适合蚂蚁繁殖。故选择云南雨季开展调查,2次调查时间分别为2015年9月与2016年8月,在晴天使用陷阱法对树冠层和地表层蚂蚁开展调查,每次调查持续10~15 d,遇到突发下雨情况重新选择晴天开展调查。参考国际上认可的蚂蚁抽样标准[24-25],在4种类型样地内各设置2个小区,各样地内设置2条样带,每条样带上设置20个地表陷阱。由于当地蚂蚁的活动范围在10 m以内(前期预调查结果),为保证每个陷阱的独立性,陷阱间距10 m,共设置地表陷阱320个。在埋置地表陷阱杯(直径为6.5 cm,高度为7 cm)时,先将陷阱杯杯口向上放置于预先挖的凹坑内,保证杯口与地面齐平并固定,然后使用周围细土填平地表与杯口的缝隙,利于陷阱杯口与地面齐平,并平整过渡,有利于提高蚂蚁的掉落率,同时将周围的生境恢复到与放置陷阱前基本一致。将φ=50%的50 mL乙二醇溶液倒入陷阱杯作为保存液,体积为杯容量的1/3。同时,在样带上或者样带附近选择20株乔木,相邻乔木间距至少10 m,将树栖陷阱固定在树干上,离地约1.5 m。同样使用φ=50%的乙二醇作为陷阱保存液,使用蜂蜜和金枪鱼混合物作为诱饵;为防止蚂蚁接触或食用诱饵,用支架将诱饵立于陷阱瓶中央(高度约为杯子高度的3/4处)。待陷阱放置72 h后,收集陷阱中蚂蚁标本,并转移至装有φ=75%乙醇溶液的离心管中保存[26]。实验室主要根据文献[27]进行标本鉴定并统计数量,对于无法鉴定到种的标本以形态种对待[28]。另外,需要说明的是,由于咖啡纯林样地内无乔木,未进行树栖蚂蚁调查,只设置240个树栖陷阱。
表1 各样地信息
Table 1 Characteristics of the four sites

样地代码样地类型作物 密度/(株·hm-2) 人为干扰情况 灌草覆盖度/% DY钝叶黄檀-玉米混农林(对照)钝叶黄檀30~45人工除草和施肥2次·季-1,旱季撂荒40 玉米9.6×103 K咖啡纯林(阳光咖啡)咖啡3×104定期进行人工除草和害虫的化学防治15~20 DK钝叶黄檀-咖啡混农林(荫蔽咖啡)钝叶黄檀30~45定期进行人工除草和害虫的化学防治15~20 咖啡2×104XK橡胶-咖啡混农林(荫蔽咖啡)橡胶2.1×103定期进行人工除草和害虫的化学防治15~20 咖啡2×104
1.4 分析方法
(1)物种组成及多度:将2次调查数据合并,对树冠层和地表层蚂蚁分别进行数据分析。为了防止部分种类蚂蚁样本量过大而被大量计数,使用国际上常用的6级计分制对蚂蚁多度数据进行转换(1分=1头;2分=2~5头;3分=6~10头;4分=11~20头;5分=21~50头;≥50头为6分)[29]。使用EstimateS 9.1.0软件计算ACE估计值(abundance-base coverage estimator)[30]。
(2)抽样充分性:抽样充分性是多样性调查的基础,只有当抽样充分时数据才具有代表性[31]。使用R语言软件中iNEXT软件工具包绘制基于个体数的物种稀疏及预测曲线[32]。
(3)多样性:将样带作为重复,1种类型样带有2条样带,2次数据则有4个重复。根据各类型样地的蚂蚁物种丰富度、多度及ACE(abundance-base coverage estimator)估计值[33-34],使用PASW Statistics 18.0软件,经方差齐性检验和正态分布检验后,采用单因素方差分析对4种类型样地的数据进行多重比较(LSD)。
(4)群落结构:使用PRIMER 7软件中的非度量多维尺度(non-metric multi-dimensional scaling,NMDS)工具分析4种类型样地树冠层和地表层蚂蚁群落结构差异(基于Bray-Curtis半度量距离矩阵)[35],同时计算群落结构差异显著性。使用层次聚类(hierarchical cluster analysis)对4种类型样地蚂蚁群落中40%相似水平的样地以圆圈的方式叠加到NMDS结果图中[36]。
(5)指示物种:使用R语言中的Labdsv软件工具包计算各类型样地的物种指示值(IndVal值),其值为Aij×Bij,Aij表示物种i在样地j中的特异性,Bij表示物种i在样地j中的保真度。以IndVal值≥0.7作为确定指示物种的标准[37]。
2 结果与分析
2.1 物种丰富度及多度
共采集到蚂蚁标本6 180头,隶属于6亚科35属71种。咖啡纯林(K)内共采集到标本1 089头,隶属于3亚科17属38种;钝叶黄檀-咖啡混农林(DK)内共采集到标本2 301头,隶属于6亚科23属48种;钝叶黄檀-玉米混农林(DY)内共采集到标本1 747头,隶属于6亚科20属42种;橡胶-咖啡混农林(XK)内共采集到标本1 043头,隶属于3亚科17属37种(表2)。
表2 各类型样地蚂蚁名录及多度
Table 2 Ant species and their abundance in the four types of habitats
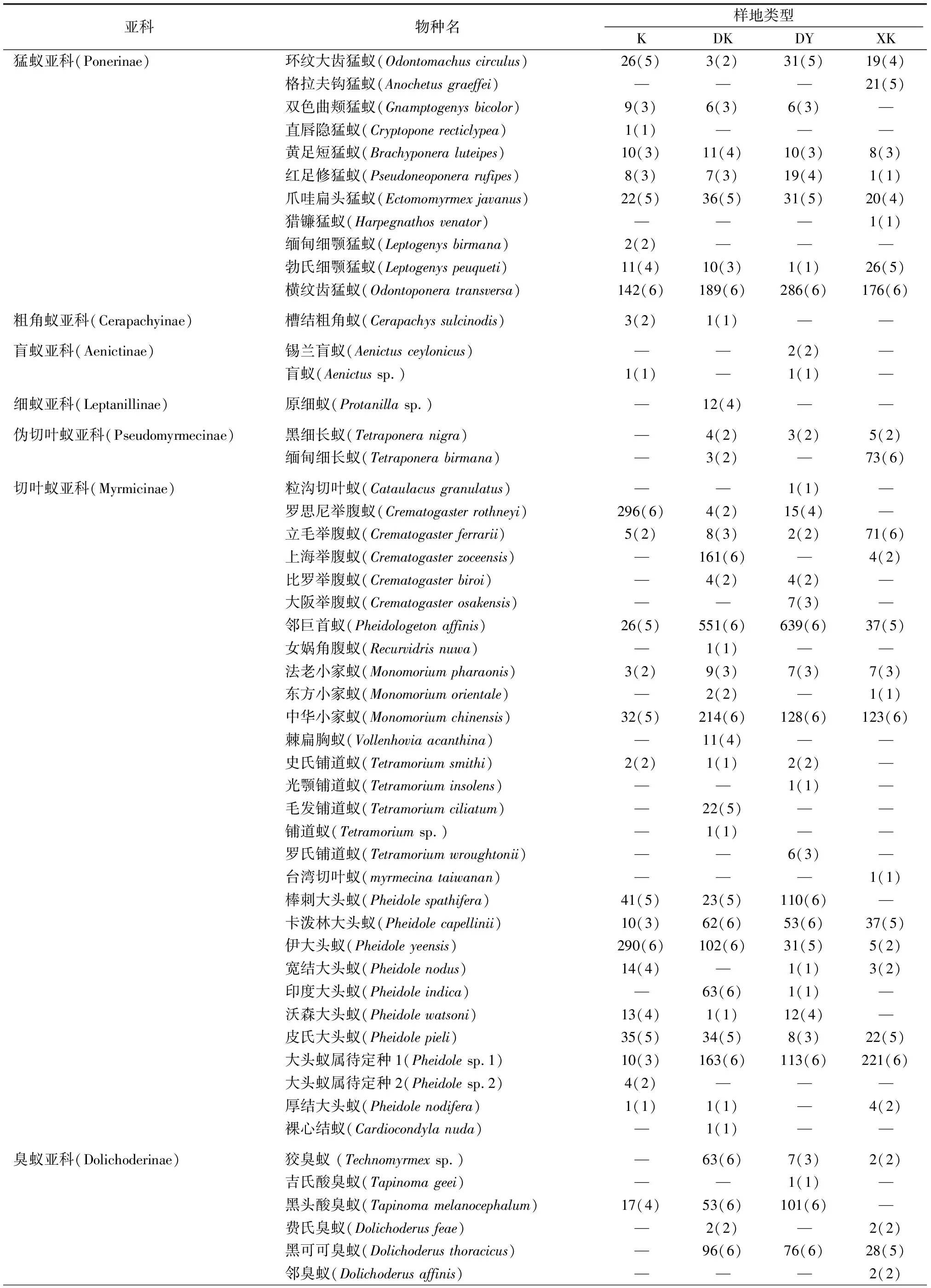
亚科物种名样地类型 KDKDYXK猛蚁亚科(Ponerinae)环纹大齿猛蚁(Odontomachus circulus)26(5)3(2)31(5)19(4) 格拉夫钩猛蚁(Anochetus graeffei)———21(5) 双色曲颊猛蚁(Gnamptogenys bicolor)9(3)6(3)6(3)— 直唇隐猛蚁(Cryptopone recticlypea)1(1)——— 黄足短猛蚁(Brachyponera luteipes)10(3)11(4)10(3)8(3) 红足修猛蚁(Pseudoneoponera rufipes)8(3)7(3)19(4)1(1) 爪哇扁头猛蚁(Ectomomyrmex javanus)22(5)36(5)31(5)20(4) 猎镰猛蚁(Harpegnathos venator)———1(1) 缅甸细颚猛蚁(Leptogenys birmana)2(2)——— 勃氏细颚猛蚁(Leptogenys peuqueti)11(4)10(3)1(1)26(5) 横纹齿猛蚁(Odontoponera transversa)142(6)189(6)286(6)176(6) 粗角蚁亚科(Cerapachyinae)槽结粗角蚁(Cerapachys sulcinodis)3(2)1(1)—— 盲蚁亚科(Aenictinae)锡兰盲蚁(Aenictus ceylonicus)——2(2)— 盲蚁(Aenictus sp.)1(1)—1(1)— 细蚁亚科(Leptanillinae)原细蚁(Protanilla sp.)—12(4)—— 伪切叶蚁亚科(Pseudomyrmecinae)黑细长蚁(Tetraponera nigra)—4(2)3(2)5(2) 缅甸细长蚁(Tetraponera birmana)—3(2)—73(6) 切叶蚁亚科(Myrmicinae)粒沟切叶蚁(Cataulacus granulatus)——1(1)— 罗思尼举腹蚁(Crematogaster rothneyi)296(6)4(2)15(4)— 立毛举腹蚁(Crematogaster ferrarii)5(2)8(3)2(2)71(6) 上海举腹蚁(Crematogaster zoceensis)—161(6)—4(2) 比罗举腹蚁(Crematogaster biroi)—4(2)4(2)— 大阪举腹蚁(Crematogaster osakensis)——7(3)— 邻巨首蚁(Pheidologeton affinis)26(5)551(6)639(6)37(5) 女娲角腹蚁(Recurvidris nuwa)—1(1)—— 法老小家蚁(Monomorium pharaonis)3(2)9(3)7(3)7(3) 东方小家蚁(Monomorium orientale)—2(2)—1(1) 中华小家蚁(Monomorium chinensis)32(5)214(6)128(6)123(6) 棘扁胸蚁(Vollenhovia acanthina)—11(4)—— 史氏铺道蚁(Tetramorium smithi)2(2)1(1)2(2)— 光颚铺道蚁(Tetramorium insolens)——1(1)— 毛发铺道蚁(Tetramorium ciliatum)—22(5)—— 铺道蚁(Tetramorium sp.)—1(1)—— 罗氏铺道蚁(Tetramorium wroughtonii)——6(3)— 台湾切叶蚁(myrmecina taiwanan)———1(1) 棒刺大头蚁(Pheidole spathifera)41(5)23(5)110(6)— 卡泼林大头蚁(Pheidole capellinii)10(3)62(6)53(6)37(5) 伊大头蚁(Pheidole yeensis)290(6)102(6)31(5)5(2) 宽结大头蚁(Pheidole nodus)14(4)—1(1)3(2) 印度大头蚁(Pheidole indica)—63(6)1(1)— 沃森大头蚁(Pheidole watsoni)13(4)1(1)12(4)— 皮氏大头蚁(Pheidole pieli)35(5)34(5)8(3)22(5) 大头蚁属待定种1(Pheidole sp.1)10(3)163(6)113(6)221(6) 大头蚁属待定种2(Pheidole sp.2)4(2)——— 厚结大头蚁(Pheidole nodifera)1(1)1(1)—4(2) 裸心结蚁(Cardiocondyla nuda)—1(1)—— 臭蚁亚科(Dolichoderinae)狡臭蚁 (Technomyrmex sp.)—63(6)7(3)2(2) 吉氏酸臭蚁(Tapinoma geei)——1(1)— 黑头酸臭蚁(Tapinoma melanocephalum)17(4)53(6)101(6)— 费氏臭蚁(Dolichoderus feae)—2(2)—2(2) 黑可可臭蚁(Dolichoderus thoracicus)—96(6)76(6)28(5) 邻臭蚁(Dolichoderus affinis)———2(2)
续表2 Table 2 (Continued)

亚科物种名 样地类型 KDKDYXK蚁亚科(Formicinae)开普刺结蚁(Lepisiota capensis)16(4)9(3)3(2)— 暗淡刺结蚁(Lepisiota opaca)1(1)1(1)1(1)1(1) 网纹刺结蚁(Lepisiota reticulate)2(2)——— 罗思尼斜结蚁(Plagiolepis rothneyi)—3(2)—2(2) 长足光结蚁(Anoplolepis gracilipes)7(3)147(6)10(3)2(2) 宾氏长齿蚁(Myrmoteras binghamii)———1(1) 普通拟毛蚁(Pseudolasius familiaris)2(2)41(5)—— 拟毛蚁 (Pseudolasius sp.)—1(1)—— 长角立毛蚁(Paratrechina longicornis)1(1)——7(3) 黄足立毛蚁(Paratrechina flavipes)3(2)——— 大眼平结蚁(Prenolepis magnocula)——1(1)— 黄猄蚁(Oecophylla smaragdina)—1(1)1(1)10(3) 邻居多刺蚁(Polyrhachis proxima)1(1)—2(2)— 伊劳多刺蚁(Polyrhachis illaudata)—3(2)—— 毛钳弓背蚁(Camponotus lasiselene)—3(2)4(2)4(2) 巴瑞弓背蚁(Camponotus parius)14(4)135(6)2(2)50(5) 平和弓背蚁(Camponotus mitis)7(3)22(5)7(3)45(5) 红头弓背蚁(Camponotus singularis)1(1)——— 蚁亚科待定种———1(1)
K为咖啡纯林;DK为钝叶黄檀-咖啡混农林;DY为钝叶黄檀-玉米混农林;XK为橡胶-咖啡混农林。括号内数值为六级计分制转化后的多度。“—”表示未采集到该品种蚂蚁。
2.2 抽样充分性
4种类型样地的树冠层和地表层蚂蚁物种稀疏及预测曲线结果见图1~2。结果显示,虚线部分均趋于平缓,说明各类型样地抽样充分。

K—咖啡纯林;DK—钝叶黄檀-咖啡混农林;DY—钝叶黄檀-玉米地;XK—橡胶-咖啡混农林。实线为实际物种数和多度,虚线为估计值,阴影部分为累积曲线的置信区间。q=0代表使用全部物种。
2.3 蚂蚁多样性比较
由表3可知,地表蚂蚁物种丰富度〔F(3,12)=3.74,P=0.04〕、多度〔F(3,12)=6.36,P=0.01〕及ACE估计值〔F(3,12)=5.08,P=0.02〕均存在显著差异,DK样地物种丰富度、多度和ACE估计值最高,XK样地最低。树栖蚂蚁物种丰富度〔F(2,9)=1.15,P=0.36〕、多度〔F(2,9)=0.35,P=0.71〕及ACE估计值〔F(2,9)=0.69,P=0.53〕无显著差异。

DK—钝叶黄檀-咖啡混农林;DY—钝叶黄檀-玉米地;XK—橡胶-咖啡混农林。实线为实际物种数和多度,虚线为估计值,阴影部分为累积曲线的置信区间。q=0代表使用全部物种。
2.4 群落结构相似性
地表层和树冠层蚂蚁群落结构相似性分析结果(图3~4)显示,4种类型样地蚂蚁群落结构均存在显著差异(地表ANOSIM GlobalR=0.319,P=0.001;树栖ANOSIM GlobalR=0.451,P=0.004)。在地表层蚂蚁群落中,DK样地与DY样地相似度高,与XK样地相似程度低,与K样地也具有一定相似性;树栖层DK样地与DY样地的树栖蚂蚁群落结构相似度高,与XK样地几乎不相似。
表3 地表层和树冠层蚂蚁多样性比较
Table 3 Species richness, abundance and ACE of ground-foraging and canopy-foraging ants

样地类型地表层树冠层多度物种丰富度ACE值多度物种丰富度ACE值 K104.75±9.34a18.25±1.25b22.27±2.39bc DK109.00±15.69a25.50±1.85a32.41±3.46a64.00±10.97a13.75±2.29a17.31±2.93a XK56.25±11.21b15.25±1.55b17.69±1.15c50.75±10.04a12.25±0.48a15.05±2.04a DY89.00±12.43ab19.00±2.08b26.23±3.41ab45.75±3.35a12.00±1.47a13.46±1.90a
K为咖啡纯林;DK为钝叶黄檀-咖啡混农林;DY为钝叶黄檀-玉米混农林;XK为橡胶-咖啡混农林。同一列数据后英文小写字母不同表示不同样地类型间某指标差异显著(P<0.05)。

K—咖啡纯林;DK—钝叶黄檀-咖啡混农林;DY—钝叶黄檀-玉米地;XK—橡胶-咖啡混农林。

DK—钝叶黄檀-咖啡混农林;DY—钝叶黄檀-玉米地;XK—橡胶-咖啡混农林。
2.5 指示物种
将4种类型样地蚂蚁物种指示值≥0.7的物种作为指示物种(表4),其中地表层K样地的蚂蚁指示物种为开普刺结蚁,DK样地为毛发铺道蚁,DY样地为罗氏铺道蚁;树冠层中XK样地的蚂蚁指示物种为立毛举腹蚁。
3 讨论
笔者研究发现,阳光咖啡地表蚂蚁物种丰富度、ACE值显著低于钝叶黄檀-咖啡样地,橡胶咖啡地表蚂蚁多度最低。另外,不同种植模式对蚂蚁群落结构也产生一定的影响。阳光咖啡与钝叶黄檀-咖啡及钝叶黄檀-玉米样地中地表蚂蚁群落有一定的相似性,但橡胶-咖啡地表蚂蚁群落与上述样地的差异性较大。树冠层钝叶黄檀-咖啡样地与钝叶黄檀-玉米样地的树栖蚂蚁群落结构相似度高,与橡胶-咖啡样地几乎不相似。4种类型样地各有1种指示物种。综上,阳光咖啡对蚂蚁多样性的保护较弱于隐蔽咖啡。2种荫蔽咖啡种植模式比较而言,选择乡土树种有利于蚂蚁群落多样性的保护。
表4 各类型样地指示物种
Table 4 Indicator species in the four types of habtiats

采集位置样地类型指示物种指示值P值 地表层K开普刺结蚁0.730.01 DK毛发铺道蚁0.750.03 DY罗氏铺道蚁0.750.03 树冠层XK立毛举腹蚁0.950.01
K为咖啡纯林;DK为钝叶黄檀-咖啡混农林;DY为钝叶黄檀-玉米混农林;XK为橡胶-咖啡混农林。P值是在1 000 次重复基础上计算得到的。表中仅列出具有统计学差异的指示物种。
国外对遮荫系统的研究显示,不同遮荫树种对当地生物多样性的影响存在差异[32]。阳光咖啡和荫蔽咖啡是云南咖啡种植的2种主要模式,但不同模式下的蚂蚁多样性及具体树种选择对蚂蚁多样性的影响研究鲜见报道。在前人的研究中,阳光咖啡和荫蔽咖啡对当地蚂蚁多样性的影响不一致[38]。阳光咖啡的高度集约化管理会导致蚂蚁物种丰富度和多度降低,而荫蔽咖啡则表现出与原有生境类似甚至更高的蚂蚁物种丰富度[6,13]。在该研究中,阳光咖啡地表蚂蚁物种丰富度、ACE值显著低于钝叶黄檀-咖啡样地,橡胶-咖啡地表蚂蚁多度最低。说明阳光咖啡对蚂蚁多样性具有一定程度的负面影响,而荫蔽咖啡对于蚂蚁多样性有保护价值,但与具体树种选择有关,这与前人研究结论一致[6,13]。
蚂蚁的群落结构也能作为指示生境变化响应的因子[39]。荫蔽咖啡复合系统的植物群落结构对蚂蚁多样性的影响存在差异[10]。对于遮荫种植模式的研究显示,不同的遮荫树种会提供不同栖境,从而吸引不同的鸟类物种栖息[40]。该研究与上述研究结果类似,在荫蔽咖啡复合系统中,无论是地表还是树栖蚂蚁群落结构,都显示出相似度低或完全不相似的结果,说明荫蔽咖啡的树种使环境发生了变化,栖息地的变化导致了蚂蚁群落的差异。并且,保护乡土树种有利于当地物种的保护[41]。钝叶黄檀-咖啡混农林与钝叶黄檀-玉米地的地表和树栖蚂蚁群落结构相似度都较高。综上所述,选择乡土树种钝叶黄檀-咖啡混农林这种咖啡种植模式,可以在保护本土蚂蚁的同时,提高蚂蚁多样性。
蚂蚁作为指示物种能较好地反映环境特征,在农业系统中能敏感地对生境状态进行响应[19,42]。毛发铺道蚁依赖较好质量的栖境生存[43],在卢志兴等[44]的研究中,毛发铺道蚁只出现在生境最佳的样地里,是天然次生林的指示物种。笔者研究中,毛发铺道蚁是钝叶黄檀-咖啡混农林的指示物种,说明钝叶黄檀-咖啡林的生境质量较好。罗氏铺道蚁与半翅目昆虫间存在明显的互利关系[45],钝叶黄檀上曾放养过紫胶虫,钝叶黄檀-玉米混农林中的指示物种为罗氏铺道蚁,这证明紫胶虫与蚂蚁的互利关系造成的影响具有一定的时空效应,与前人研究一致[25]。举腹蚁属是常见于咖啡树上的关键捕食者[42],其不仅捕食一般的食草性昆虫,还会捕食危害咖啡浆果的蛀虫[46-47]。橡胶-咖啡混农林的指示物种为立毛举腹蚁,验证了橡胶-咖啡混农林中害虫多的现象。开普刺结蚁喜食咖啡上的介壳虫蜜露,会搬运蚧虫[41,48],咖啡纯林中的指示物种为开普刺结蚁,验证了林内咖啡介壳虫危害严重的现象。
综上所述,咖啡单作对当地蚂蚁多样性有一定的负面影响,荫蔽咖啡有利于生物多样性的保护,但其保护成效与遮荫树种的选择有关。由当地树种钝叶黄檀作为遮荫树种的钝叶黄檀-咖啡混农林有利于当地的蚂蚁多样性保护,且有较高的生态效益,是咖啡有机种植模式的较好选择。另外,笔者研究与传统的生物多样性研究相比存在一定的差异性,特别是引入了物种累计曲线、ACE值及指示物种,这些尝试有利于与国外的同类研究进行比较,应该是有益的尝试。
致谢:感谢赵婧文和付兴飞在野外调查和内业工作上给予的帮助与支持。
