黄土丘陵区草地根系生物量对踩踏干扰生物结皮的响应
2019-12-20赵允格许明祥徐安凯
任 伟,赵允格,许明祥,徐安凯
(1. 中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨陵 712100; 2. 吉林省农业科学院, 吉林 长春 130124; 3. 中国科学院大学, 北京 100049)
生物土壤结皮(简称生物结皮),是由苔藓、地衣、蓝藻、细菌或真菌等形成的十分复杂的复合体,约占陆地表面的35%~70%,常与高等维管束植物呈镶嵌式分布,相互影响[1]。一方面,生物结皮改善了地表微环境,提高了0~2 cm土壤有机碳、全氮、碱解氮及速效磷等养分含量,细化土壤质地,显著改善土壤理化性质,促进高等植物生长发育及群落演替[2]。另一方面,生物结皮在土壤表面形成一个致密层,显著降低土壤水分渗透性[3],还可能降低维管束植物种子进入土壤和幼苗萌发出土的机会[4-5],成为维管束植物生长发育的负面影响因子。有关生物结皮与高等维管束植物之间的关系一直是生物结皮生态功能研究的重要内容,已有学者就生物结皮对维管束植物种子萌发、种子库、物种入侵、植物群落等方面开展了不少研究,取得重要进展[6-8]。
干扰是自然界中常见现象,以藻类、苔藓类及地衣类等变水植物为主要成分的地表生物结皮,对各种自然和人为干扰(火烧、踩踏、机械碾压、翻耕等)非常敏感[9]。干扰极大地影响了生物结皮的盖度和组成,改变了其下层土壤养分循环和水文过程[10]。研究表明,干扰显著降低藻结皮和藓结皮的盖度,降低土壤有机碳、全氮、速效氮的含量,增加易氧化碳含量和有机碳累积矿化量[11-12]。其次,干扰可引起生物结皮层以及0~2 cm土壤细菌、真菌、放线菌的变化[13]。就对土壤水文过程方面,石亚芳等研究表明,干扰生物结皮可增加降水入渗,减小径流,改善土壤水分状况[14]。可见干扰可显著影响生物结皮土壤养分及水分状况,进而可能导致与其共生的高等维管束植物生长发育的变化。然而鲜有研究调研干扰生物结皮对高等维管束植物根系的影响,是生物结皮与高等维管束植物关系研究的薄弱环节。
根系是植物的重要器官,具有锚定植物、吸收输导土壤水分养分、合成和储藏营养物质等诸多生理功能[15]。根系具有很高的可塑性,能够快速响应土壤养分和水分变化,进而实现根系在获取与保守策略中的权衡调控[16]。根系生物量,占据了植物体总生物量的二分之一,不仅与植株地上部生长有关,同时可敏感反应植物获取水、土资源能力,具有重要的生态指示功能[17]。
生物结皮是黄土丘陵区退耕地上广泛发育的地被物[18],平均盖度可达60%~70%,是该区退耕地植被恢复演替的重要影响因子。明确高等维管束植物对干扰生物结皮的响应,对促进该区植被恢复具有重要意义。为此,本研究以黄土丘陵区有生物结皮发育的撂荒草地为样地,通过模拟不同强度羊只踩踏干扰(后文简称“干扰”),分析不同强度干扰对土壤理化属性以及根系生物量的影响,以期揭示干扰生物结皮对根系生长的影响及机制,为黄土丘陵区退耕地植被恢复提供理论依据。
1 材料与方法
1.1 研究区概况
研究区域位于陕西省吴起县合沟流域(36°58' N,107°50' E),属于温带大陆性季风气候,地貌类型为典型的黄土丘陵梁峁沟壑区,平均海拔1 400 m,年均气温7.8℃,年均日照时数2 400 h,年均降水量475 mm,雨季主要集中在7—9月。土壤类型为黄绵土,主要植被群落为碱菀(TripoliumvulgareNees)、长芒草(StipabungeanaTrin.)、委陵菜(PotentillachinensisSer.)、糙隐子草(Cleistogenessquarrosa(Trin.) Keng)、胡枝子(LespedezabicolorTurcz.)、黄耆(Astragalusmembranaceus(Fisch.) Bunge)、狗娃花(Heteropappushispidus(Thunb.) Less.)、狭裂白蒿(ArtemisiakanashiroiKitam.)、茵陈蒿(ArtemisiacapillarisThunb.)、龙胆(GentianascabraBunge)、远志(PolygalatenuifoliaWilld.)、中华小苦荬(Ixeridiumchinense(Thunb.) Tzvel.)等草本植物,植被盖度约25%。研究区退耕封禁约20年,生物结皮发育良好,与植被镶嵌分布,且未被扰动,覆盖度在70%以上,主要以藻结皮、藻藓混生结皮、藓结皮以及少许地衣结皮为主。
1.2 试验设计
为明确不同强度干扰对与生物结皮镶嵌分布的维管束植物的影响,试验设计轻(20%)、中(40%)、重(50%)3个干扰强度,以生物结皮破碎度表征干扰度。以不干扰为对照,每个处理设4个重复,共计16个小区。已有研究表明,当研究区生物结皮破碎度大于50%时,有加剧水土流失的风险[14],而控制水土流失是研究区生物结皮的重要生态功能,因此,本研究设计的最大干扰度为50%。
1.3 试验小区布设及干扰处理
2015年7月在研究区选择生物结皮发育良好,坡度一致(25°)撂荒坡地布设试验小区。小区规格为1.5 m×3.0 m,间隔3.0 m,采用铁皮圈建小区,随机区组试验设计。然后采用25点样方法(样方25 cm×25 cm)调查生物结皮的盖度和组成,采用样方法调查植被盖度,采样测定各小区0~10 cm,10~20 cm土壤全氮磷、速效氮磷、有机碳等养分含量,作为研究样地的背景资料(表1)。完成后,对小区进行干扰,干扰方法详细参见杨雪芹等[11]。干扰后,采用样方调查、统计得出野外用于试验的3个干扰梯度的实际干扰度分别为:19.18±0.52%,37.02±1.72%和48.04±3.21%(后文简写为19%,37%和48%干扰度)。此后,每2个月对小区进行调查、观测,如果生物结皮有所恢复,则采用原方法对其进行重复干扰,保证干扰度保持在设计干扰度的±5%以内。

表1 研究小区样地背景值Table 1 Background of the study area
1.4测定指标与方法
根系生物量:干扰1年后,于2016年9月于每个小区上、中、下部3个样点,用直径10 cm的根钻分层采集0~10 cm,10~20 cm的土芯,带回实验室,水中浸泡1~2 h,然后小心冲洗,收集根系。滤纸吸干表面水分后,放入铝盒中60℃恒温烘至恒重,得到根系干重(Root dry weight,RDW),采用式(1)计算根系生物量(Root Biomass,RB),用式(2)计算干扰后根系生物量的增量(ΔRB)。
RB(g·m-2)=RDW/(π×r2)
(1),
其中:RB—根系生物量;RDW—根系干重;r—根钻半径。
ΔRB(g·m-2)=干扰小区RB—不干扰小区RB
(2),
土壤理化属性测定:采集根系样品时,用便携式测定仪(TRIME-PICO)确定土壤含水量、温度和电导率。分别采集0~10 cm,10~20 cm土壤样品,将每个小区上、中、下部3个样点的同层土壤混合作为1个样品,带回实验室。测定土壤pH(水土比1∶2.5)、有机碳(重铬酸钾容量法)、全磷(氢氧化钠熔融—钼锑抗比色法)、速效磷(碳酸氢钠浸提-钼锑抗比色法)、全氮(凯氏定氮法)、速效氮(碱解扩散法)。具体见鲁如坤《土壤农化分析》第三版。
1.5 数据处理
运用Microsoft Excel 2010对数据进行统计、计算,采用GraphPad Prism 8.0进行绘图。运用Kolmogorov-Smirnov test检验数据的正态分布,利用Levene's test进行方差齐性检验。采用SPSS 18.0软件中的ANOVA单因素方差分析检验根系生物量及土壤理化属性在不同干扰度之间的差异性,并进行Duncan多重比较。采用Pearson相关分析法计算根系生物量与土壤理化属性的相关系数,并做进一步的回归分析。
2 结果与分析
2.1 干扰后生物结皮组成和盖度的变化
干扰显著影响生物结皮的盖度和组成,随干扰度增强,藓结皮和藻结皮盖度显著降低(图1)。48%干扰度组藓结皮盖度为7.20%,较不干扰组降低66.69%。37%干扰度组藻结皮盖度为12.10%,较不干扰组降低50.21%。干扰显著增加植物盖度,37%干扰度组(33.20%)较不干扰组增加43.10%。调落物盖度在不同干扰度组间差异不显著。扰动藻结皮(本文指生物结皮经践踏、动物活动及试验干扰破损后又有所恢复的藻结皮)盖度随干扰度增强显著增加,48%干扰度组(28.41%)较不干扰组增加667.84%。
不干扰的小区,藻结皮、藓结皮、植物、凋落物的盖度百分比大致相当,分别为:24.30%,21.60%,23.20%,20.90%。19%干扰度下,凋落物盖度百分比最大,为26.10%。37%干扰度下,植物所占盖度百分比最大,为33.20%。48%干扰度下,扰动藻和植物所占盖度百分比较大,分别为28.41%和27.93%。总的来看,干扰显著降低结皮盖度,增加植物和扰动藻的盖度。

图1 不同干扰度对生物结皮覆盖组成的影响Fig.1 Effect of the different disturbance intensity on the composition of surface coverage
2.2 干扰对植物根系生物量的影响
干扰显著影响植物根系总生物量(图2A),干扰1年后,随干扰度增强,根系总生物量呈现先增加后降低的趋势。37%干扰度组的根系总生物量为427.73 g·m-2,较不干扰组增加104.55%。19%和48%干扰度组与不干扰组差异不显著。
37%干扰度组显著增加0~10 cm土层根系生物量(344.10 g·m-2),与不干扰组相比,Δ根系生物量为231.13 g·m-2,增幅达204.59%。19%,48%干扰度组亦增加了0~10 cm根系生物量,但Δ根系生物量较小,与不干扰组差异不显著。10~20 cm土层根系生物量有随干扰强度增加而降低的趋势,但与不干扰组差异不显著(图2A、B)。
不干扰组0~10 cm和10~20 cm土层根系生物量分别占总根系生物量的54.03%和45.97%,差异不显著,0~10 cm与10~20 cm土层根系生物量比值为1.18。干扰显著影响了根系剖面分布,19%干扰度组0~10 cm和10~20 cm土层根系生物量分别占总根系生物量的64.54%和35.46%,0~10 cm与10~20 cm土层根系生物量比值为1.82。37%干扰度组0~10 cm土层根系生物量占总根系生物量的80.45%,10~20 cm土层根系生物量占总根系生物量的19.55%,0~10 cm与10~20 cm土层根系生物量比值为4.11。48%干扰度组0~10 cm和10~20 cm土层根系生物量分别占总根系生物量的64.75%和35.25%,0~10 cm与10~20 cm土层根系生物量比值为1.84。表明干扰生物结皮可影响根系的空间分布,导致根系分布浅层化(图2C)。
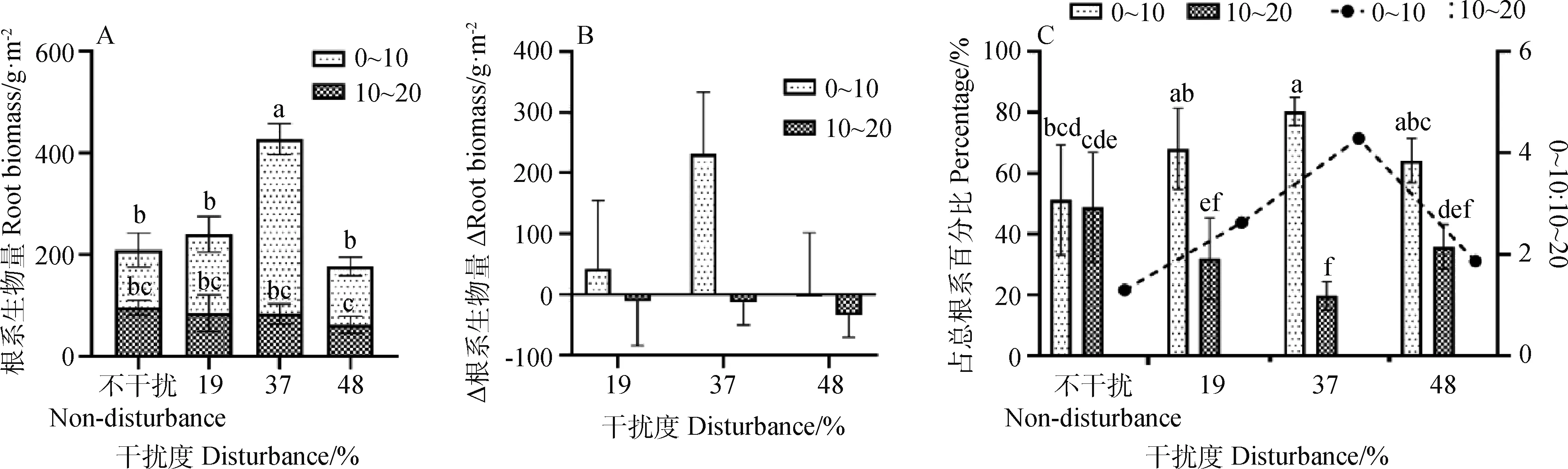
图2 不同干扰度对植物根系的影响Fig.2 Effect of the different disturbance intensity on the root biomass注:A:根系生物量;B:Δ根系生物量;C:百分比;不同小写字母表示不同干扰度不同土层间差异显著(P<0.05),下同Note:A:root biomass;B:Δ root biomass;C:percentage;Different lowercase letters indicate significant difference between soil layers with different disturbance intensity at the 0.05 level,the same as below
2.3 干扰对土壤理化属性的影响
水分:随干扰度增强,0~10 cm土壤含水量显著增加,48%干扰度组土壤含水量较不干扰组增加22.37%,10~20 cm土壤水分先增加后降低,37%干扰度组较不干扰组增加25.36%(图3A);温度:干扰显著降低下层土壤温度,呈现出先降低后增加的趋势,其中37%干扰度组土壤温度较不干扰组降低5.85℃(图3B)。
养分:干扰后生物结皮下层土壤有机碳含量有下降趋势,但差异不显著(图3C);全氮:干扰增加0~10 cm土层全氮含量,37%和48%干扰度组较不干扰组分别增加8.20%和6.56%,且差异显著(图3D);有效氮:随干扰度增强,0~10 cm土壤有效氮含量呈现先降低后增加趋势,10~20 cm土壤有效氮含量呈递增趋势(图3E);全磷:不同处理及土层间全磷含量波动较小,无显著差异(图3F);有效磷:0~10 cm土壤有效磷含量呈现随干扰度增强先增加后降低趋势,但差异不显著,10~20 cm土壤有效磷含量差异也不显著(图3G)。不同土层之间,0~10 cm土壤水分和养分均高于10~20 cm土层。
其它:生物结皮干扰之后,0~10 cm和10~20 cm土壤pH、电导率随干扰强度无显著变化(图3H;图3I)。

图3 不同干扰度对生物结皮下层土壤理化属性的影响Fig.3 Effect of the different disturbance intensity on thesoil physical-chemical properties
2.4 干扰影响植物根系生物量的机制
为了解析干扰对草地根系分布特征与土壤属性之间的关系,对根系生物量与土壤水分、温度、养分进行相关及回归分析(表2,图4)。结果表明,根系生物量与土壤水分呈显著正相关(皮尔逊相关系数0.432),二次函数拟合较好(图4A),表明干扰增加了降水的入渗,促进了植物根系的生长。根系生物量与土壤温度呈显著负相关(皮尔逊相关系数-0.383),线性函数拟合较好(图4B),表明炎热的夏季,干扰生物结皮在一定范围内(27.93~22.08℃)降低午后土壤温度,有利于植物根系的生长。根系生物量与土壤全氮呈极显著正相关(皮尔逊相关系数0.583),线性函数拟合较好(图4C)。根系生物量与土壤有效氮呈显著正相关(皮尔逊相关系数0.369),线性函数拟合较好(图4D)。根系生物量与土壤全磷呈显著正相关(皮尔逊相关系数0.357),二次函数拟合较好(图4E)。根系生物量与土壤有效磷呈极显著正相关(皮尔逊相关系数0.520),二次函数拟合较好(图4F)。表明干扰生物结皮增加下层土壤全氮含量,土壤有效氮、全磷、有效磷呈现增加趋势,为植物根系提供更多可供吸收利用的养分,进而提高了根系生物量。在黄土丘陵地区,通过干扰生物结皮影响土壤氮磷的养分循环以及水分、温度的重新分配,促进与之镶嵌分布的植物根系生长。

表2 根系生物量与土壤理化属性的相关分析Table 2 Correlation analysis between root biomass and soil physical-chemical properties
注:*代表显著相关(P<0.05),**代表极显著相关(P<0.01)
Note:* indicates significant correlation at the 0.05 level,** indicates extremely significant correlation at the 0.01 level
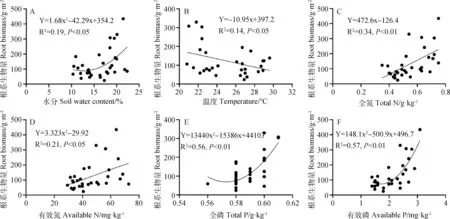
图4 根系生物量与土壤理化属性的回归分析Fig.4 Regression analysis of root biomass and soil physical-chemical properties
3 讨论
根系是植物生态进化的驱动器,在植物群落演替,养分水分吸收,土壤微生物驱动,碳氮循环等过程中具有重要作用[19]。根系的生物量、分布及构型,反映了植物地上部的生长潜力[20]。本研究结果表明,研究区植物根系主要分布在表层土壤,0~10 cm土层根系生物量高于10~20 cm土层,呈“T”型分布,与前人研究结果相一致[21-22]。
然而,干扰生物结皮显著影响了植物根系生物量,且影响程度因土层而异,0~10 cm土层,随着干扰强度的增加,植物根系生物量呈现先增加后降低的趋势,其中37%干扰度组根系生物量最大,呈现浅层化趋势。可能是由于根系分布特征不仅与其自身遗传信息有关,同时取决于外界环境因子的影响[23]。干扰改变了生物结皮的组成与分布[24],影响下层土壤养分、水分的分布格局,进而影响植物根系的生长发育。正如本研究中,干扰增加了雨水的入渗,随干扰度增强,下层土壤含水量的增加,有利于植物根系生长。此外,与不干扰相比,干扰显著降低夏季午后土壤温度,其中37%干扰度组降低5.85℃,可能与该干扰度促进了植物生长,地上部盖度较大(增加43.10%),遮阴较好,降低了土壤温度,间接影响了植物根系生长有关。
此外,干扰增加了扰动藻类结皮的盖度,可能促进了结皮层氮素养分随入渗水分向下的迁移,因此37%和48%干扰度组显著增加0~10 cm土壤全氮含量。随着干扰度的增强,0~10 cm土壤有效氮含量呈现先降低后增加趋势,影响植物的吸收利用。王闪闪等[12]研究结果也表明干扰降低了结皮层有效氮的含量。但10~20 cm土层有效氮含量呈现逐渐增加趋势,并且有效氮与根系生物量总体呈正相关关系,表明干扰生物结皮增加了下层土壤氮的养分供给,进而增加了根系生物量。0~10 cm土壤有效磷含量随干扰度增强,呈现先增加后降低趋势,与根系生物量变化趋势相同,是由于磷元素在土壤中移动性较差,而植物根系的新陈代谢以及分泌的有机酸活化了土壤中的磷元素[25],提高了土壤有效磷的含量。有效磷的增加进一步又促进了植物地下部的生长发育,提高了根系生物量,因此二者之间呈极显著正相关关系。总的来看,干扰影响了生物结皮下层土壤养分含量、剖面分布以及土壤水分、温度等,进而促进植物根系的生长。
根系生物量的增加及浅层化,不仅可以稳定土壤结构,增强土体抗剪强度[26],而且可以增加土壤渗透能力,提高土壤抗侵蚀能力[27]。本研究发现干扰生物结皮增加了植物根系生物量,从根系角度揭示踩踏干扰生物结皮对植被的影响,为今后黄土丘陵区植被恢复、水土流失治理以及草牧业发展提供借鉴意义。
4 结论
干扰显著降低结皮盖度,增加维管束植物盖度;干扰生物结皮增加了根系生物量,改变了植物根系剖面分布,上下层根系生物量比值较不干扰增加0.64~2.93倍,导致草地根系浅层化;干扰增加0~10 cm土壤含水量,降低夏季午后土壤温度,增加土壤全氮含量;干扰生物结皮影响了土壤养分循环以及水文、温度的重新分配,促进了与之镶嵌分布的植物根系的生长。
