干旱-复水对旱柳生长与光合生理特性的影响
2019-12-19管健古琳刘俊祥
管健,古琳,刘俊祥*
(1.辽宁生态工程职业学院,沈阳 110101;2.中国林业科学研究院林业研究所,国家林业和草原局林木培育重点实验室,北京 100091)
全球气候变化背景下,干旱发生的频率和强度不断加剧,给林木生长发育造成严重伤害。干旱胁迫导致净光合速率、蒸腾速率、气孔导度、胞间CO2浓度显著降低[1],光合机构受损[2],水分利用效率下降[3],使木本植物的碳平衡和水力传导失调,面临着死亡的威胁[4]。干旱胁迫下的生理、生长表征和复水后的生理、生长补偿共同决定了木本植物应对干旱胁迫的生存策略[5],复水后的补偿效应是检验旱害解除后木本植物恢复能力的标准[6]。研究干旱和旱后复水条件下木本植物动态的生长和光合生理响应,可为揭示其对干旱胁迫的适应机制奠定基础。旱柳(Salixmatsudana)为杨柳科(Salicaceae)柳属(Salix)植物,是我国北方地区重要的农田防护、平原造林和园林绿化树种,生态适应性广泛,生产应用价值高。本文以旱柳为试验材料,采用盆栽控水的方法研究了干旱-复水条件下旱柳的生长和光合生理响应,拟明确旱柳应对干旱胁迫的应对策略,解析胁迫强度对复水后生长补偿效应的影响,揭示旱柳对干旱-复水的适应性机制,以期为耐旱林木资源的筛选和培育提供理论参考。
1 材料与方法
1.1 材料培养
2015年4月12日取旱柳1年生枝条作为插穗,在中国林业科学研究院科研温室扦插培养,基质为草炭土∶珍珠岩=3∶1,常规养护管理。取大田20 cm表层园土,过2 mm筛,自然风干。园土田间持水量为26.68%,pH值7.35,有效磷42.69 mg/kg,有效氮104.43 mg/kg,有效钾107.59 mg/kg,有机质26.87 mg/kg,土壤全磷0.08%,全氮0.10%,全钾1.56%。2015年6月10日,选取株高 50 ± 5 cm的旱柳扦插苗,清水洗净根部后转移至装有3 kg园土的花盆中,保持园土含水量为田间持水量的80%,花盆规格19 cm(上口)×17 cm(下口)×21 cm(高)。
1.2 实验设计
2015年7月15日,选取株高106±9 cm的盆栽苗,进行水分控制。共设置4个处理:对照(CK,土壤相对含水量80%~85%);轻度干旱(T1, 60%~65%);中度干旱(T2, 40%~45%);重度干旱(T3,30%~35%)。每处理6盆,3次重复。控水期间,对照保持设定的水分含量,其他处理自然干旱达到预设梯度后维持其含水量。2015年7月19日,T3处理达到预设梯度,以此为实验0天。在干旱处理12天后,将T1、T2和T3复水至CK水平。干旱和复水期间每天称重补水,保持各处理的土壤含水量为预设梯度。分别于实验0天和干旱处理后12天、复水后12天测定枝径、叶片数量;于干旱处理后12天和复水后12天测定叶片相对含水量和根系生物量;于干旱处理后4天、8天、12天和复水后4天、8天、12天测定叶片气体交换参数、实际光化学效率。
1.3 实验方法
1.3.1 枝径相对增长量、叶片数相对变化量的测定。从每处理固定1株旱柳盆栽苗,记录展开叶片数量;用游标卡尺测定盆土面上方10 cm处的枝条直径;按照下面公式计算干旱期间和复水期间枝径相对增长量、叶片数相对变化量。
干旱期间:
枝径相对增长量=(S1-S0)/S0
叶片数相对变化量=(L1-L0)/L0
复水期间:
枝径相对增长量=(S2-S1)/S1
叶片数相对变化量=(L2-L1)/L1
式中:S0-实验0天的枝径(mm);S1-干旱处理12天后的枝径(mm);S2-复水后12天的枝径(mm)。L0-实验0天的叶片数;L1-干旱处理12天后的叶片数;L2-复水后12天的叶片数。
1.3.2 根系生物量和叶片相对含水量和的测定。从每处理随机选2株旱柳盆栽苗,取植株根系,去离子水洗净,105 ℃杀青20 min,80 ℃烘干至恒重,测定根系生物量;取3片完全展开叶片,参考高俊凤(2006)的方法测定各处理的叶片相对含水量。
1.3.3 气体交换参数和实际光化学效率的测定。从每处理随机取2株,每株选2片完全展开叶片,用Li-6400光合仪(Licor,美国)在上午9:00至下午15:00期间测定叶片气体交换参数,包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)。红蓝光源光强1500 μmol/m2·s,相对湿度60%~70%,温度25 ℃,CO2浓度400 μmol/m2·s。
从每处理随机取2株,每株选2片完全展开叶片,用Imagine-PAM调制叶绿素荧光仪(Walz,德国)于上午9:00至下午15:00期间测定叶片实际光化学效率(ΦPSⅡ),活化光光强1176 μmol/m2·s。
1.4 数据分析
采用SPSS19.0对所得数据进行方差分析和DUNCAN多重比较,在0.05水平上检验同一天数不同处理间的差异显著性;采用Excel2007进行数据整理和图表绘制。由于干旱处理后4天, Li-6400的水分控制失效,导致胞间二氧化碳浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)的读数异常,而净光合速率(Pn)的计算与湿度无关,故干旱处理4天后的气体交换参数只保留了净光合速率。
2 结果与分析
2.1 干旱与旱后复水对旱柳生长的影响
由表1可知,干旱阶段,中度和重度干旱处理的枝径相对增长率、叶片数相对变化量均较对照显著降低,特别是在重度干旱条件下,枝径增长较对照下降68%,叶片脱落70%,地上部生长受到严重抑制。复水后,各处理的枝径相对增长量均恢复至对照水平,重度干旱处理的叶片数呈爆发式增加。
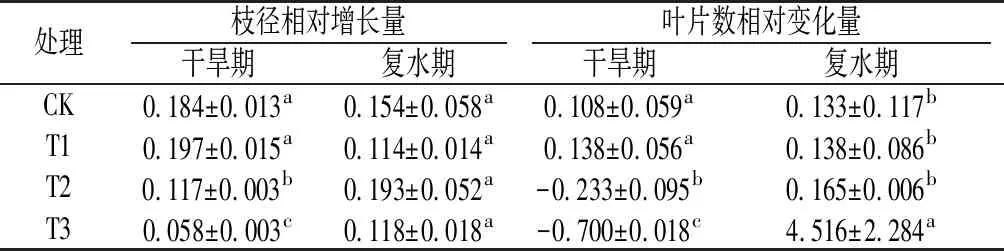
表1 干旱-复水条件下旱柳枝径生长和叶片数量的变化
注:表中同列数据小写字母不同表示在0.05水平上差异显著。
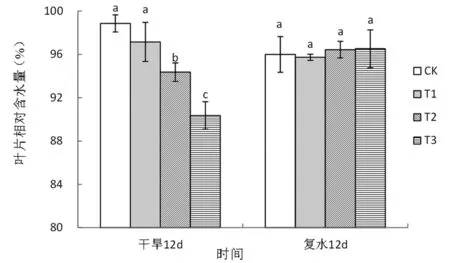
图1 干旱-复水条件下旱柳叶片相对含水量的变化
注:图中相同时间不同处理间小写字母不同表示在0.05水平上差异显著,以下图与此相同。
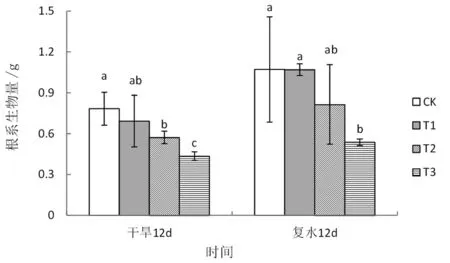
图2 干旱-复水条件下旱柳根系生物量的变化
图1的结果表明,干旱处理12天后,叶片相对含水量随干旱程度的增加显著降低,中度和重度干旱分别较对照显著下降了4.56%和8.60%,复水12天后,二者均恢复至对照水平。根系生物量受重度干旱胁迫显著抑制,处理12天后较对照显著下降44.44%,复水12天后仍显著低于对照49.86%(图2)。
2.2 干旱与旱后复水对旱柳叶片气体交换参数的影响
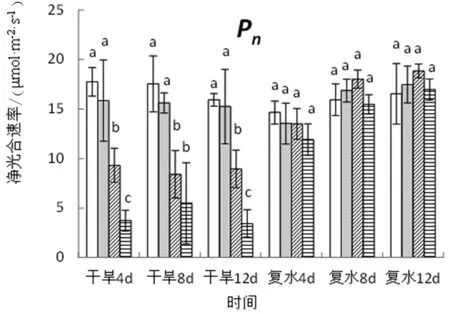
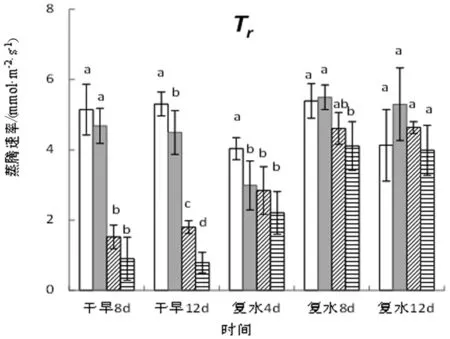
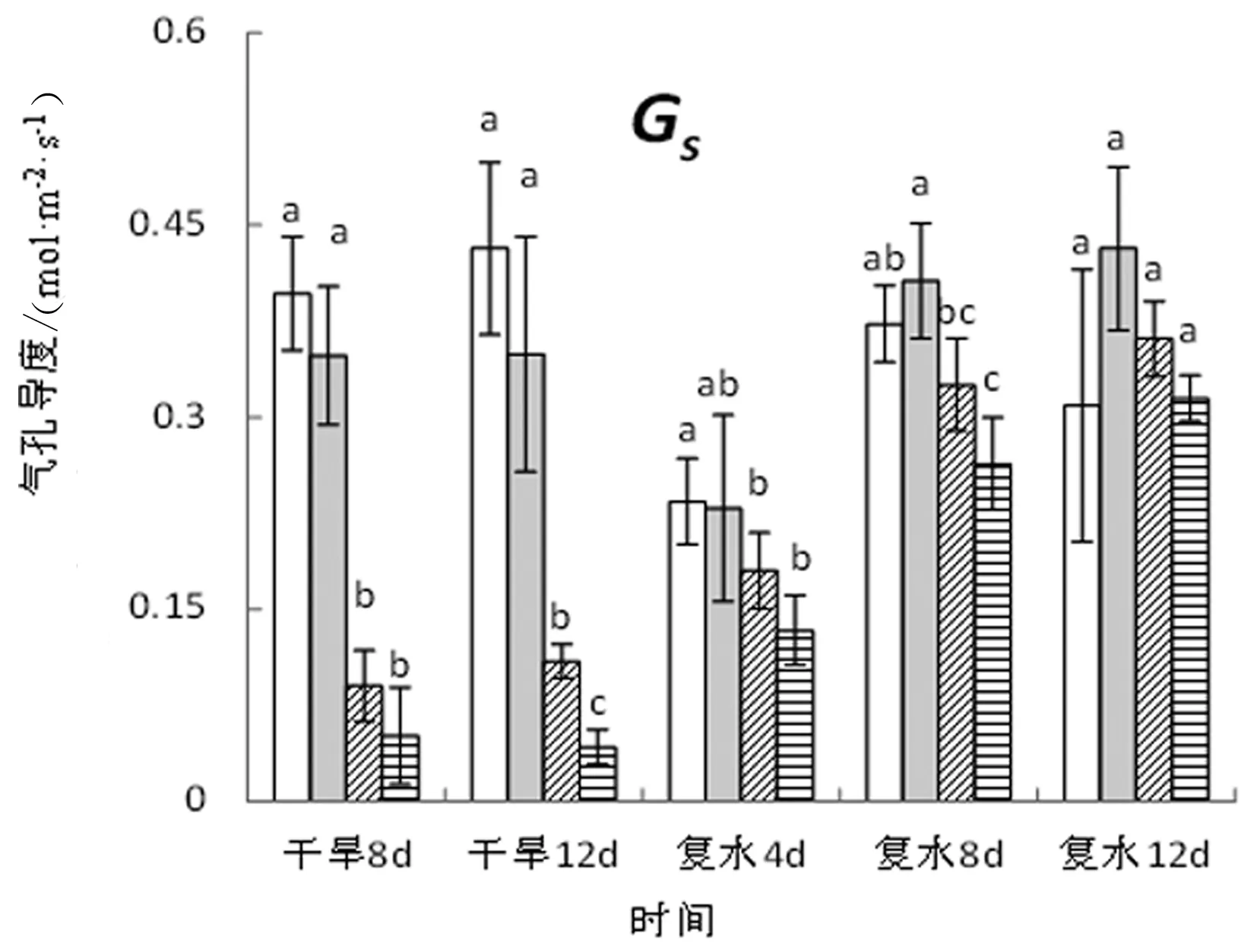
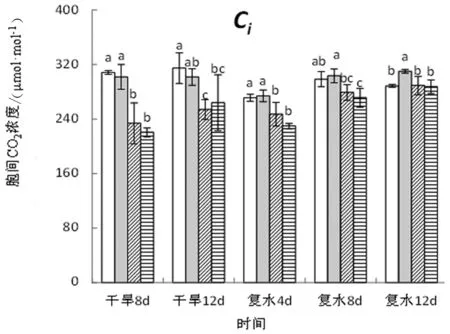
图3 干旱-复水条件下旱柳叶片气体交换参数的变化
由图3可知,在干旱阶段,旱柳叶片气体交换参数随干旱胁迫程度的加剧而显著降低,处理12天后,中度干旱处理的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)分别较对照显著下降43.78%、66.08%、74.60%、19.34%,重度干旱分别较对照显著下降了78.45%、85.04%、90.08%和 16.13%。复水4天后,各干旱处理的净光合速率即可恢复至对照水平,而对于其他3项参数,复水后的恢复受干旱胁迫强度的影响,强度越大,恢复越慢。
2.3 干旱与旱后复水对旱柳叶片实际光化学效率的影响
图4的结果表明,在干旱复水的整个过程中,轻度和中度干旱条件下旱柳叶片的实际光化学效率在均与对照无显著性差异。重度干旱处理后4、8、12天,叶片实际光化学效率分别较对照显著下降了58.95%、39.04%和22.90%,复水后4天恢复至对照水平。
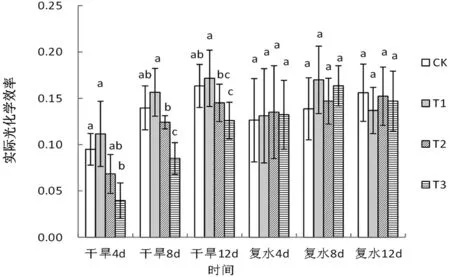
图4 干旱-复水条件下旱柳叶片实际光化学效率的变化
3 结论与讨论
3.1 干旱胁迫导致旱柳生长受抑、碳水失衡
干旱胁迫下,根系吸水困难,枝条水力传导受阻,叶片水分供给不足[8]。在中度和重度干旱胁迫下,旱柳通过降低叶片的含水量、气孔导度和蒸腾速率以缩减水分耗散,是一种对干旱胁迫的避性选择,表现出等水植物的特点[9]。但叶片对CO2的吸收依赖于水分的交换[10],水分耗散的减少将导致CO2供给受限,叶片净光合速率降低[11],同时干旱导致叶片脱落,总光合面积减少[12],最终导致了中度干旱胁迫下旱柳的生长受抑。除上述伤害表征外,在重度干旱胁迫下,旱柳叶片的实际光化学效率被显著抑制,使叶片的光合速率进一步降低,增加了碳饥饿的风险[13]。
3.2 旱后复水,旱柳生长的恢复受干旱胁迫强度的影响
干旱复水后,重度干旱处理的旱柳叶片数相对变化量显著高出对照水平,表现出超补偿效应,这与刘晓英等[14]的研究结果一致。复水对叶芽的发生发育有补偿作用,且这种补偿作用与干旱胁迫梯度正相关,重度干旱复水后叶片的爆发式增加是一种弥补碳亏缺的有效措施。干旱复水后,根系的响应与叶片不同。在中度干旱胁迫下,复水12天后根系生物量可恢复至对照水平,而重度干旱胁迫复水后根系生长未能得到恢复,郝树荣等[15]的研究也得到了相似的结果。重度干旱导致叶片严重脱落,根系主要依赖储存的碳库进行维持呼吸,复水12天后新生叶片的同化能力有限,暂时不能满足根系生长对同化物和同化力的需求。
3.3 旱后复水,光合速率的恢复不决定于气孔限制的解除
中度干旱处理的气孔导度、蒸腾速率和胞间CO2浓度在复水8天后恢复;重度干旱处理下,3项气体交换参数在复水12天后才得到恢复;气孔导度、蒸腾速率和胞间CO2浓度的恢复速度受干旱胁迫强度的负调控。而复水4天后,中度、重度干旱处理的净光合速率即可恢复至对照水平。因此,叶片光合速率的恢复并不决定于气孔限制的解除,这可能依赖于叶绿体光合活性的迅速提高。在复水4天后,各干旱处理的PSⅡ实际光化学效率即可恢复至对照水平。此外,推测光合色素光能吸收传递效率的提高[16]、类囊体垛叠结构的增加[17]及基质中Rubisco等暗反应酶活性的提升[18]等生理上的补偿可能均参与到叶片光合速率的快速恢复,需要在本研究的基础上进一步的验证和探索。
综上,碳饥饿是旱柳旱害的主要诱因。复水后,为增加碳的回补,旱柳表现出叶片爆发式增加、光化学效率和净光合速率快速恢复等表征。在复水后短期内,气孔限制因子并未解除,叶片光合活性的提高对光合速率的恢复起到决定性作用。
