井冈山鹿角杜鹃群落灌木层功能多样性及其随海拔梯度的变化
2019-12-18陈芳清官守鹏王玉兵
向 琳,陈芳清,官守鹏,王玉兵,吕 坤
三峡地区生态保护与治理国际联合研究中心,三峡大学, 宜昌 443002
功能多样性是指某一群落内物种或特定生态系统内所有物种功能特征数值和分布的变化范围[1]。植物的功能性状是生态系统多性状环境适应策略的稳定组合,对生态系统功能有着重要影响[2]。功能多样性的变化可以通过功能性状变化反映出来[3],而功能多样性对生态系统功能影响的研究可以通过量化功能性状来实现[4]。相较于物种多样性主要体现的是生物与环境间的复杂关系以及生物种类的丰富程度[5],功能多样性更多地强调群落内不同物种功能性状间的差异,反映共存物种的互补和冗余[6],并可以根据物种的性状特征来表征不同的生态系统功能[7]。物种多样性和功能多样性的特征及其相互之间的关系是生态系统功能研究的重要内容[8]。有研究表明群落生态系统多样性很大程度上取决于其功能多样性[9-10],但也有人认为将基于功能性状的功能多样性与基于物种的物种多样性结合研究,能更好的认识和了解群落植物多样性与生态系统功能的关系[11-12]。由于不同环境梯度上光照强度、温度、降水等生态因子有着较大的变化,群落的物种多样性和功能多样性常随着环境梯度发生变化[13-14]。海拔梯度是一种能够在较小的空间范围呈现出不同生境类型的环境梯度。伴随着海拔梯度的变化,各环境因子呈现出梯度性变化,并由此引起植物功能性状发生相应的变化[15]。开展物种多样性、功能多样性与以海拔为主导的小尺度空间环境因子之间关系的研究,能较好地揭示海拔梯度上功能多样性与生态系统功能的关系及其维持机制,对深刻认识群落的多样性和资源的可持续利用具有重要的指导意义[7]。
井冈山位于南岭山地罗霄山脉中段,是江西境内分布的典型二元山体结构,地形多样,具有特殊的盆岭相间格架。复杂的地形地貌孕育了丰富的物种多样性,该地区是我国生物多样性分布的关键区域之一[16]。杜鹃属(Rhododendron)植物是江西省分布最广、类型最多、面积最大的一类灌丛植物。鹿角杜鹃(Rhododendronlatoucheae)是杜鹃花属中垂直分布最宽的种类之一,对环境的变化具有较强的适应性。灌丛群落物种多样性随海拔梯度的变化已经有了较多的研究[17-18],而对于不同海拔梯度下灌丛群落功能多样性的研究相对较少。本文以鹿角杜鹃灌丛群落为研究对象,对分布于井冈山地区不同海拔的该群落灌木层的植物种类及其功能性状进行了实地调查与分析,旨在解决以下科学问题:鹿角杜鹃群落灌木层植物物种多样性和功能多样性随着海拔梯度的变化有何适应性变化;鹿角杜鹃群落灌木层植物物种多样性和功能多样性间有何相关性;地形及土壤因子对鹿角杜鹃群落灌木层植物功能多样性有何影响规律。期望通过解答这些科学问题,揭示该群落物种多样性、功能多样性的特征及其相互关系以及该群落类型对环境变化的响应趋势,为井冈山鹿角杜鹃群落资源和生物多样性的保护和管理提供科学依据。
1 材料和方法
1.1 研究地概况
井冈山地处26°27′—26°40′ N,113°39′—114°23′ E,整个山体呈东北-西南走向,最高海拔为1841 m,最低海拔为202 m,相对高差较大。该地区位于南亚热带与中亚热带的过渡地带,为亚热带湿润季风气候。土壤以山地黄壤为主,其他各类土壤随海拔梯度的增加呈现有规律的垂直带谱分布。年平均气温为14—17℃,年降雨量1856.2 mm。区域内的典型植被为常绿阔叶林,拥有灌木林、山顶矮林、针叶林、针阔叶混交林、落叶阔叶林等多种森林群落类型,森林覆盖率高达81.2%[16,19]。以杜鹃属植物为建群种的群落主要分布在井冈山较高海拔的山地,其中鹿角杜鹃主要分布于300—1200 m的范围内,群落环境具有湿度高、温度低、营养资源丰富的特点。
1.2 样地调查与样品采集
以井冈山地区海拔600—1500 m为研究区域。在低海拔、中海拔和高海拔梯度挑选具有代表性的鹿角杜鹃群落,各设置3个20 m×20 m的样地,在每个样地随机设置3个5 m×5 m的样方。调查与测量样方内灌木层各植株的种类、株数、高度、盖度、冠幅,并且测量各样地的经纬度、含水量、海拔等环境因子。海拔、坡度、土壤相关含量的数据均以现场测量值表示。其中坡位数据以1、2、3分别代表上坡位、中坡位和下坡位;坡向数据以朝东为起点(即为 0°)顺时针旋转,每45°为一个区间:1表示北坡(247.5°—292.5°),2表示东北坡(292.5°—337.5°),3表示西北坡(202.5°—247.5°),4表示东坡(337.5°—22.5°),5表示西坡(167.5°—202.5°),6表示东南坡 (22.5°—67.5°),7表示西南坡(112.5°—167.5°),8表示南坡(67.5°—112.5°),数字越大,代表越向阳,越干热[20]。本研究中各样地的生态环境特征见表1。各样地均无高大乔木出现,草本层不发达。低海拔样地草本层优势种为麦冬(Ophiopogonjaponicus)和芒萁(Dicranopterisdichotoma),中海拔草本层优势种为贯众(Cyrtomiumfortunei)和十字薹草(Carexcruciata),高海拔草本层优势种为淡竹叶(Lophatherumgracile)、双蝴蝶(Tripterospermumchinense)和华南毛蕨(Cyclosorusparasiticus)。
1.3 物种多样性计算
选取了4个常见的物种多样性指数来评估井冈山鹿角杜鹃群落灌木层物种多样性状况[21]。先计算群落中各物种的重要值,然后再分别计算各物种多样性指数。
物种丰富度:S=样地内物种数
式中,Pi为种i的相对重要值;S为种i所在样方的物种总数。
1.4 功能多样性计算
参照董世魁等[8]的研究方法,选取13种植物形态特征和生殖特征作为其功能特征(表2)。其中植物的高度和盖度通过野外调查获得,其他功能特征通过查阅《中国植物志》[22]获得。
Mason[23]等将植物功能多样性分为功能丰富度、功能均匀度和功能离散度。其中功能丰富度主要衡量群落中现有物种占据的生态位空间大小,功能均匀度主要测量物种性状在所占据性状空间的分布是否规律,而功能离散度主要描述群落功能性状的多度分布在性状空间中的最大离散程度。本研究功能丰富度指数选用功能体积FRic[24]来作为代表,功能均匀度用多维功能均匀度指数FEve[24]来表征,功能离散度选取常用的二次熵指数Rao[25]、功能分散指数FDis[24]和功能离散指数FDiv[24]这3个指数来表征。将植物功能性状中的名称数据赋值转变为数值数据(表2),然后利用R3.4.4 FD软件包中的dbFD函数计算上述5个功能多样性指数。
(1)多维功能丰富度指数FRic:计算性状空间内生成的最小多边形的体积或面积。
(2)多维功能均匀度指数FEve:
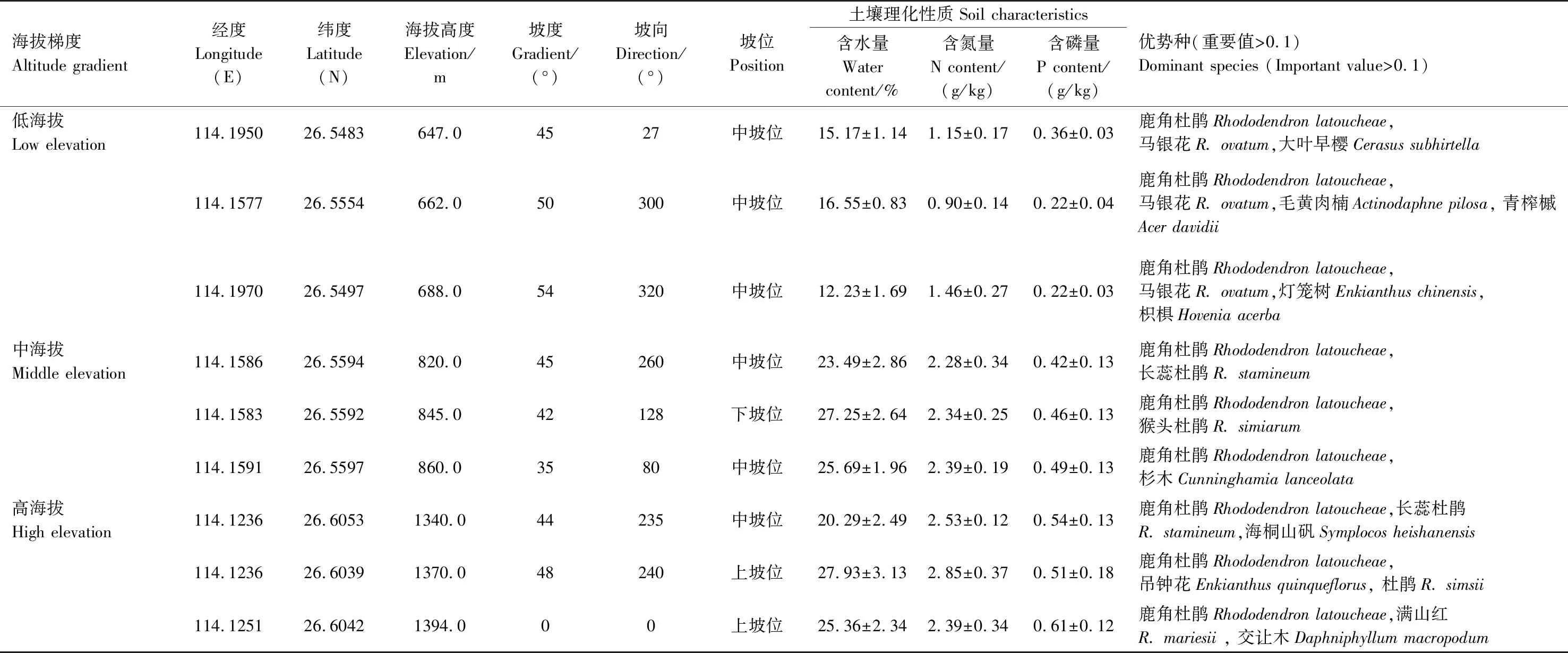
表1 不同海拔梯度群落环境特征与优势种组成
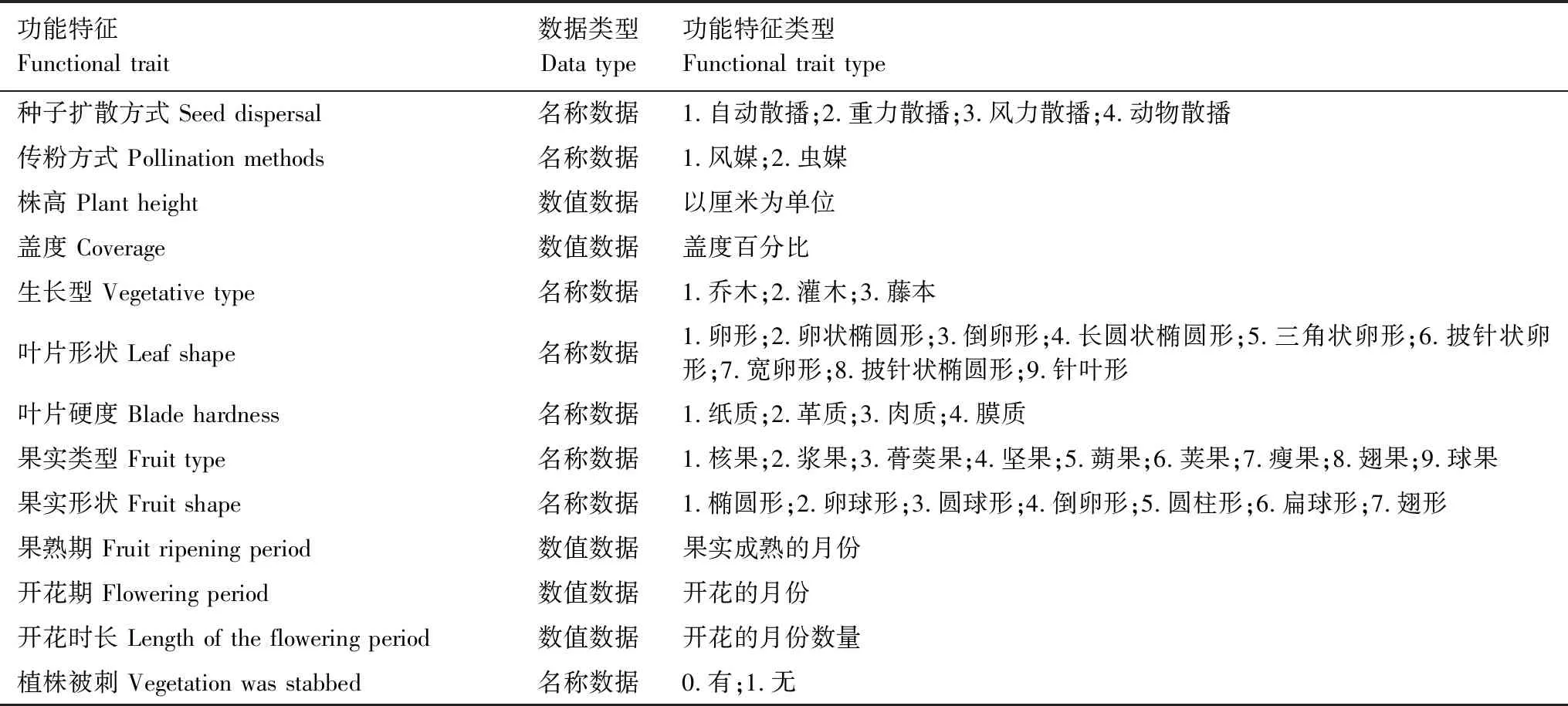
表2 植物功能特征类型
式中,EW为均匀度,Wi为物种j的相对多度。
(3)二次熵指数Rao:
式中,pi为i种的个体数占群落中总物种个体数的比例。
(4)功能分散指数FDis:
式中,aj为物种j的相对多度,xij为物种j第i个性状值。zj是物种j到重心c的加权距离。
(5)功能离散指数FDiv
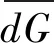
1.5 数据分析
利用SPSS 19.0统计分析软件进行以下数据分析:1)单因素方差分析法(One-Way ANOVA)。以海拔梯度为自变量,以群落的物种多样性指数和功能多样性指数为因变量进行差异显著性分析,揭示海拔梯度对物种多样性指数和功能多样性指数的影响;2)功能多样性指数与物种多样性指数间的Pearson相关性分析;3)环境因子与功能多样性指数的逐步回归分析。将环境因子作为自变量,功能多样性指数作为因变量建立线性回归模型,通过将环境因子逐个引入模型进行逐步回归分析。并利用Microsoft Excel 2010进行功能多样性指数与物种多样性指数间的关系拟合及作图。
2 结果和分析
2.1 不同海拔梯度鹿角杜鹃植物群落的物种多样性
不同海拔梯度鹿角杜鹃群落的丰富度指数、Shannon多样性指数、Pielou均匀度指数、Simpson优势度指数变化趋势基本一致,均随着海拔的升高呈减小趋势(表3)。其中丰富度指数、Pielou均匀度指数在不同海拔间的差异不显著(P<0.05);中海拔与低海拔和高海拔的Shannon多样性指数不具有显著性差异,但是低海拔与高海拔间具有显著性差异(P<0.05);低海拔和中海拔的Simpson优势度指数间不存在显著性差异,但两者与高海拔之间存在显著性差异(P<0.05)。丰富度指数在647m处最大,1340m处最小,分别为23、10;Shannon多样性指数在647m处达到最大值2.7336,在845m处达到最小值1.8219;Simpson优势度指数在647m处达到最大值,在1394m处达到最小值,分别为0.9057和0.7969。
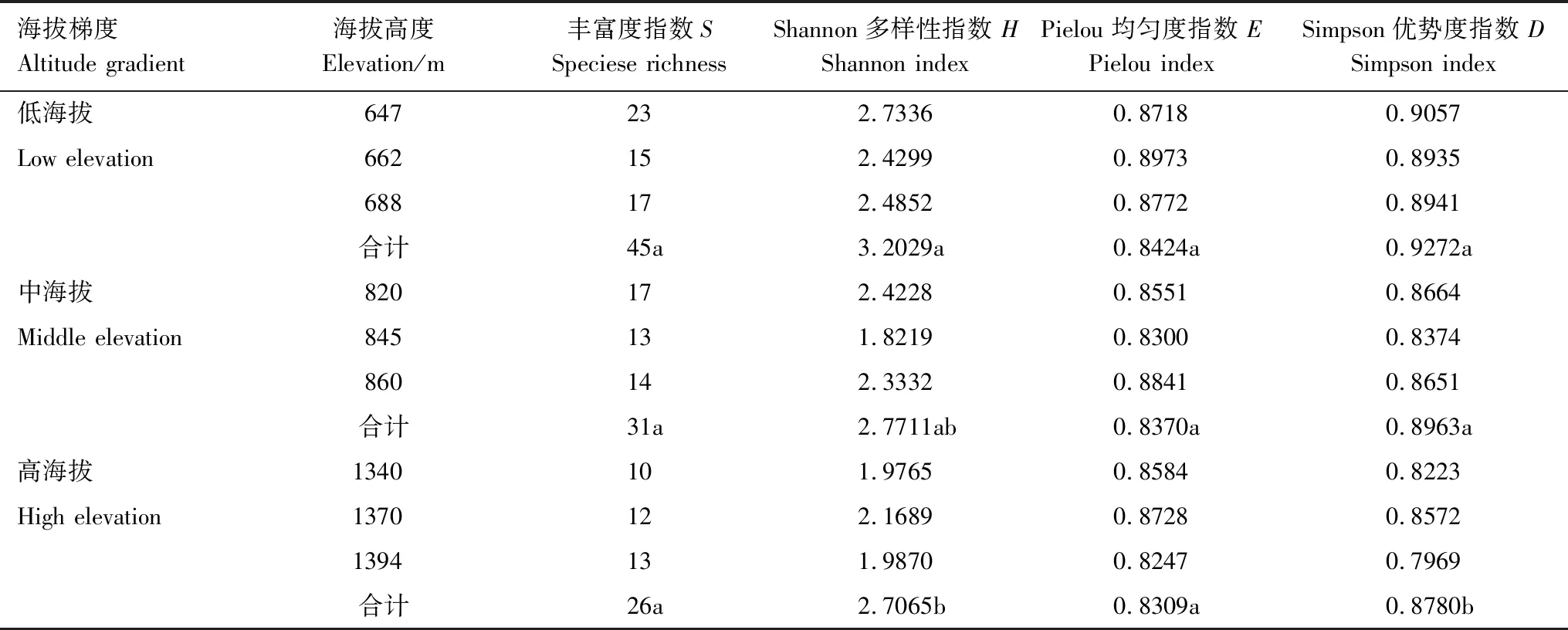
表3 不同海拔梯度鹿角杜鹃群落的物种多样性
不同字母代表物种多样性指数在不同海拔间的显著性差异(P<0.05)
2.2 不同海拔梯度鹿角杜鹃植物群落功能多样性
鹿角杜鹃群落的功能多样性指数在海拔梯度上呈现不同的变化趋势(图1),其中FRic、FEve、FDis和Rao在不同海拔梯度上具有显著差异性(P<0.05),而FDiv在不同海拔之间的差异不显著。FRic、FEve和FDis均随着海拔的升高而减少,从低海拔到高海拔群落,FRic均值分别为1.037、0.641、0.090,FEve均值分别为0.901、0.902、0.855,FDis均值为3.433、3.367、3.293,高海拔的FRic、FEve、FDis分别比低海拔的减少91.3%、5.1%、4.1%;Rao随海拔的升高而增加,从低海拔到高海拔分别为0.663、0.754、0.816,高海拔比低海拔增加了23.0%;FDiv随着海拔升高呈现增加趋势,高海拔比低海拔增加了6.0%。
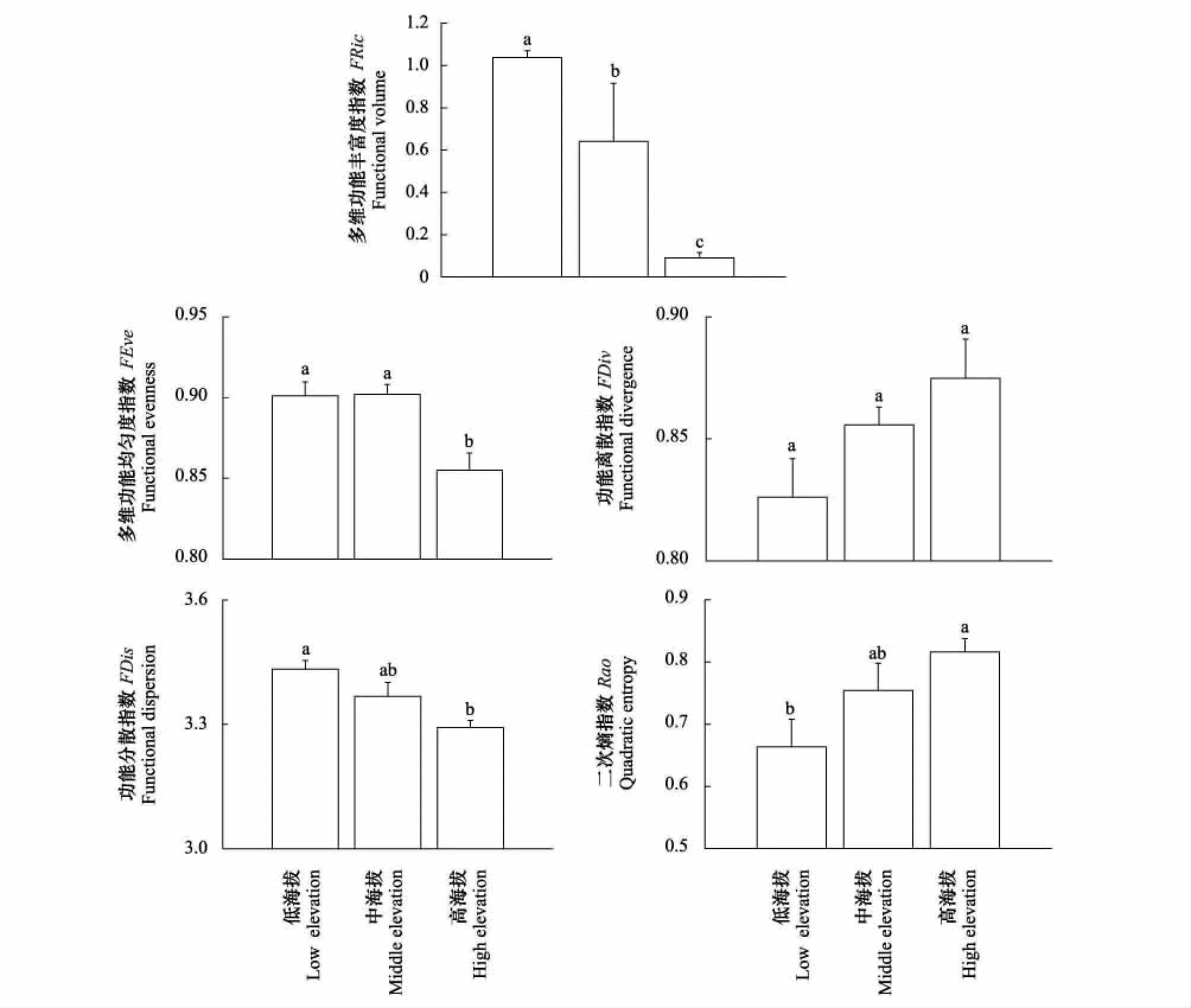
图1 鹿角杜鹃群落植物功能多样性随海拔梯度的空间变化Fig.1 Spatial variations of functional diversity indices of R. amurensis communities distributed along elevation gradient FRic: functional volume; FEve: functional evenness; FDiv: functional divergence; FDis: functional dispersion; Rao: quadratic entropy
2.3 植物群落功能多样性和物种多样性间的相关性
对不同海拔所有样地群落功能多样性和物种多样性指数的Pearson相关检验显示(表4),FRic与丰富度指数和Simpson优势度指数呈显著正相关;FEve与丰富度指数呈显著正相关;Rao与Pielou均匀度指数和Simpson优势度指数呈显著负相关;FDis与丰富度指数、Shannon多样性指数呈极显著正相关,与Simpson优势度指数呈显著正相关;FDiv与Shannon多样性指数、Pielou均匀度指数、Simpson优势度指数均呈显著负相关。其他功能多样性指数与物种多样性指数间无显著相关关系。
对海拔梯度上鹿角杜鹃群落植物功能多样性指数和物种多样性指数进行回归分析,得到5种功能多样性指数与4种物种多样性指数间的拟合关系(表5)。FRic与丰富度指数和Simpson优势度指数具有较强的相关性,与丰富度指数具有最优拟合对数方程;FEve与丰富度指数存在较强的对数函数关系,与另外3种物种多样性指数不具有显著相关性;Rao与Pielou均匀度指数、Simpson优势度指数均存在线性负相关关系,与Simpson优势度拟合效果最好;FDis与丰富度指数、Shannon多样性指数、Simpson优势度指数均具有显著相关性,与Shannon多样性指数和Simpson优势度指数最佳拟合方程为线性函数,而与丰富度指数的最优拟合方程为对数函数,且拟合度最高;FDiv与Shannon多样性指数、Pielou均匀度指数和Simpson优势度指数均具有显著负相关关系,与Pielou均匀度指数具有最优拟合线性函数。

表4 群落功能多样性指数和物种多样性指数间的Pearson相关性分析
*P<0.05; **P< 0.01;S: Speciese richness;H: Shannon index;E: Pielou index;D: Simpson index;FRic: functional volume;FEve: functional evenness;FDiv: functional divergence;FDis: functional dispersion;Rao: quadratic entropy
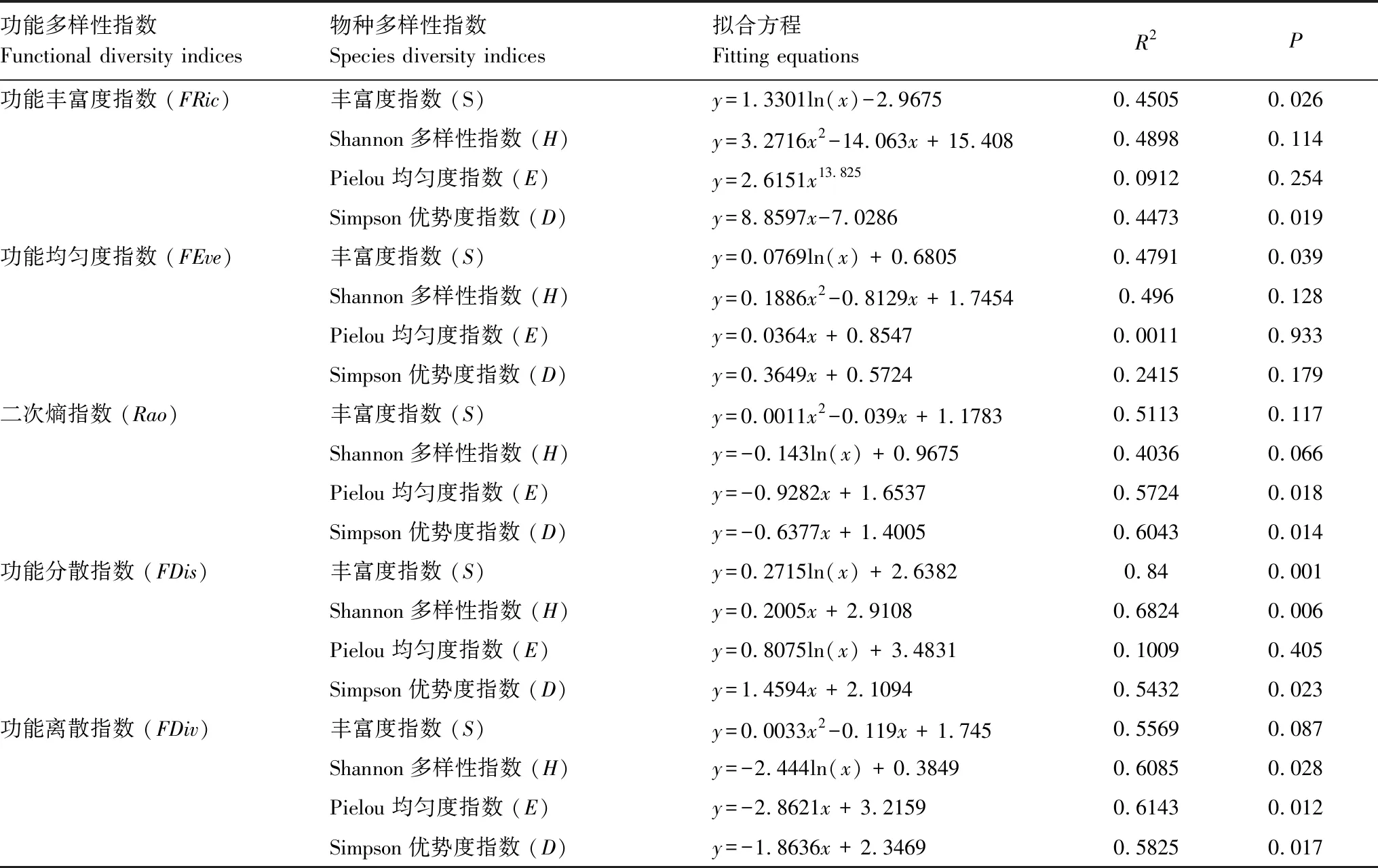
表5 群落功能多样性指数和物种多样性指数的关系拟合
S: Speciese richness;H: Shannon index;E: Pielou index;D: Simpson index;FRic: functional volume;FEve: functional evenness;FDiv: functional divergence;FDis: functional dispersion;Rao: quadratic entropy
2.4 海拔梯度上功能多样性与环境因子的逐步回归分析
对功能多样性与环境因子进行逐步回归分析进一步表明海拔是影响功能多样性的一个重要因素(表6)。海拔对FRic、FEve、Rao、FDis、FDiv的标准偏回归系数分别为-0.946、-0.740、1.015、-0.835、1.049。除海拔因素以外,FEve还受到坡位的影响,标准偏回归系数为0.481;土壤P对FRic、Rao、FDiv也有显著有影响,标准偏回归系数分别为-0.824、1.277、0.828;FDis也受到土壤N的影响,标准偏回归系数为-0.772。其他环境因子对功能多样性指数的影响不显著。
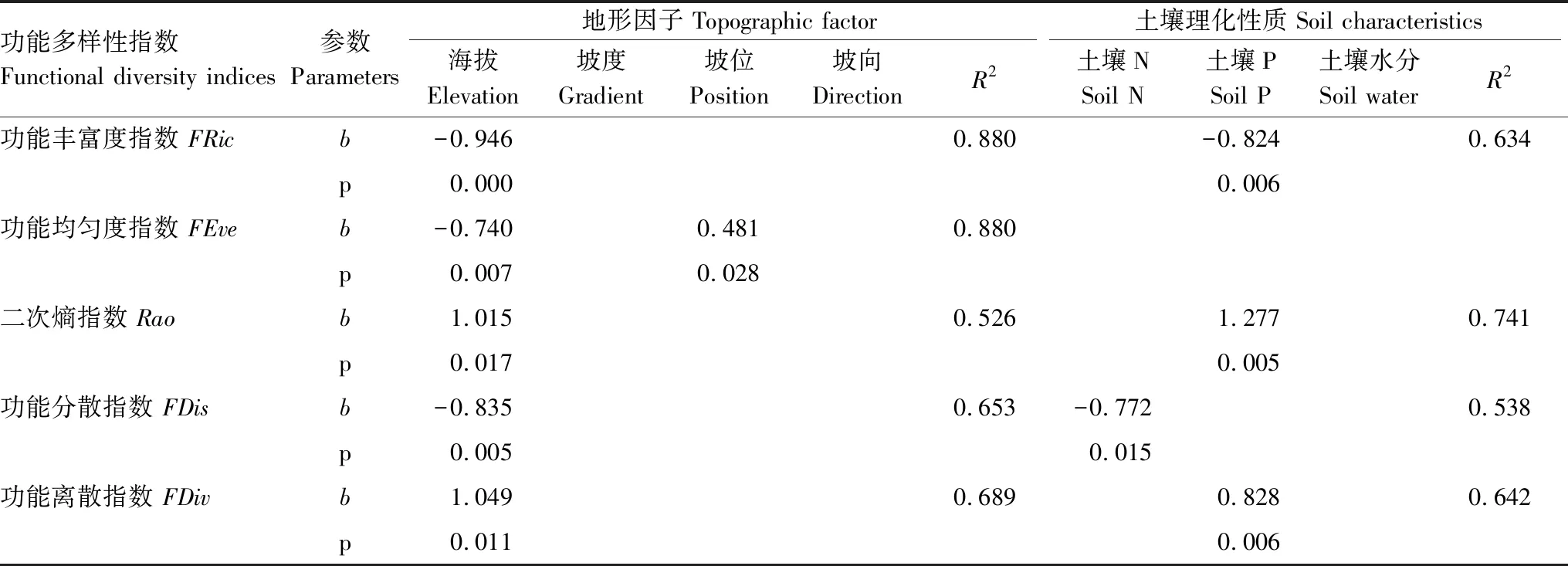
表6 功能多样性指数与环境因子间的逐步回归分析
b: 标准偏回归系数 Standard patrial regression coefficient;FRic: functional volume;FEve: functional evenness;FDiv: functional divergence;FDis: functional dispersion;Rao: quadratic entropy
3 讨论
3.1 群落植物功能多样性与物种多样性间的关系
群落中物种多样性和功能多样性往往具有复杂的相关性,在不同的群落类型和环境下会表现出不同的关系[26-28]。功能丰富度FRic与物种丰富度往往具有显著正相关关系[29-30],早前的研究中普遍认为物种丰富度可以代替功能丰富度[31-32]。随着相关研究的逐步深入,该结论被其他研究所质疑。Li等[33]和Niu等[34]认为物种丰富度并不能够完全代替功能丰富度,因为功能多样性能够提供一些不同于物种多样性的关于形态、生理、繁殖等信息;功能均匀度指数FEve与物种丰富度之间一般被认为无显著相关性[7,35-36],因为FEve更多地反映性状所占据的空间分布均匀程度[21],然而姚兰[37]和郑丽婷等[38]的研究却发现FEve与物种丰富度呈负相关关系;邵丹[39]对云蒙山国家森林公园植被的多样性进行研究时发现只有二次熵指数Rao和功能离散指数FDiv与Pielou均匀度指数有显著相关性,而功能分散指数FDis与物种丰富度、Pielou均匀度和Shannon多样性均没有显著相关性;赵小娜等[40]在研究山西文峪河上中游森林群落的多样性时发现物种多样性指数与功能多样性指数间的相关性不显著,只有FDiv与Shannon多样性指数呈显著负相关,而造成这个的原因是环境变化引起的物种增加往往为亲缘关系相近种,对功能多样性并没有显著影响;董世魁等[8]在研究高寒草地植物时发现高寒草原植物的FDis和FDiv与Simpson优势度指数具有显著相关性,而在高寒草甸中却没有此现象。物种多样性和功能多样性呈现出较高的相关性,也有可能是因为其运算方法上的相似性造成的。因为测量功能多样性所使用的方法与物种丰富度具有显著相关关系,各功能多样性指数均要在物种特征及其物种丰富度的基础上进行计算[41-42]。
本研究与以上研究结果不尽相同,鹿角杜鹃群落的功能丰富度指数FRic与物种丰富度呈显著正相关,其拟合方程为对数函数,表明物种丰富度并不能够完全代替功能丰富度。鹿角杜鹃群落的功能均匀度指数FEve与物种多样性的相关性相对较低,只与物种丰富度具有正相关关系,与其他物种多样性指数无显著相关性,造成这个现象的原因可能是因为不同的群落类型在不同的环境下物种的丰富程度对其分布的均匀程度也有一定的影响。鹿角杜鹃群落的功能离散度指数Rao、FDis、FDiv均与Simpson优势度指数呈线性相关关系且具有显著相关性,从空间尺度上看群落的Simpson优势度指数可以基本表征功能离散度。优势度高的群落优势种相对多度高,性状相对更加集中,功能离散度相对较小。
3.2 群落植物功能多样性对海拔梯度的响应
海拔是重要的地形因子,不同海拔梯度的太阳辐射强度、土壤养分、降水量等水热环境因子常存在异质性变化。海拔梯度的变化会影响植物的生长发育,并引起植物性状发生复杂的适应性变化,使植物形成不同功能性状组合的生活策略[15]。综合功能多样性指数常沿海拔梯度发生空间变化,大多数功能多样性指数随着海拔的升高呈减小趋势,与物种多样性的变化趋势一致[7,20]。本研究中,功能丰富度指数FRic、功能均匀度指数FEve、功能离散度指数FDis、Rao均与海拔具有显著线性关系,表明海拔梯度对功能多样性具有重要的影响,这与Zhang[43]关于海拔是群落结构分布、生物多样性的关键影响因素的结论一致。在井冈山鹿角杜鹃群落中,随着海拔的增加,群落的物种多样性呈下降的趋势,功能丰富度指数FRic、功能均匀度指数FEve、功能分散指数FDis也随之减小,这与国内外研究结论一致[7,40]。造成FRic随着海拔升高而减少的原因可能是因为海拔的升高导致温度降低,温度限制了植物的生长发育,造成物种种数减少,而物种的减少限制了功能性状的数值和范围[44],进而引起FRic的减少。FEve主要体现了植物对环境资源的利用效率[11],随着海拔的升高,FEve逐渐减少,说明高海拔地区鹿角杜鹃群落灌木层植物在低温环境下对环境资源的利用效率低,群落共存物种对资源的竞争更加激烈,群落一旦受到干扰,往往难以恢复。
FDiv和Rao定量描述了群落中物种功能性状特征的异质性[45]。本研究中,功能离散度指数FDiv、Rao均随着海拔的升高呈增加趋势,该结果与卜圣文[46]、Bai等[7]研究结论不一致,但与姚兰[37]和张钦弟等[20]的研究结论一致。FDiv和Rao随海拔的升高而增加,可能是由于高海拔地区温度较低,能够适应低温环境的物种较少,群落物种数量相应减小。在低效率高竞争的环境压力下,物种逐渐演变成为相似的能适应低温的形态,物种间功能特征相似性增加。该结果进一步表明了群落在高海拔更容易受到限制相似性的影响[23]。但Rao的增加会导致功能多样性(FD)的降低[45]。
3.3 群落植物功能多样性与环境因子的关系
海拔、坡位、坡度、坡向等地形因子主要是通过改变太阳辐射强度、温度、降水量以及土壤的条件来间接影响植物群落在物种分布格局、资源利用等方面的协调与平衡,这些非生物因子对植物群落的影响在小尺度区域中更加明显[38]。影响群落功能多样性的环境因子因群落类型及其分布地的不同有较大变化。朱云云等[47]认为坡向对黄土高坡草地群落功能多样性有很大的影响,田平等[14]认为坡位是影响太岳山辽东栎次生林灌木层功能多样性的主要因子。而Bai等[7]认为海拔高度对小五台山森林群落功能多样性具有重要影响,段晓梅等[48]对太岳山脱皮榆群落物种的相关研究也得到了相近的结论。本研究对井冈山地区鹿角杜鹃群落灌木层植物功能多样性与地形因子逐步回归分析发现海拔高度对功能多样性具有显著影响,而其他大部分地形因子的影响不显著,证明了海拔是影响群落组成结构和多样性的重要环境因子[49]。该结论的获得可能是本研究就是在海拔梯度的基础上研究功能多样性,一定程度影响到其他一些地形因子效应的表现。群落功能多样性在不同海拔表现出来的差异性并不仅仅只有海拔变化的影响,而是其他环境因子综合作用的结果。本研究还表明,FEve还受到坡位的显著影响,上坡位往往光照较强,温度较高,植物都能够很好地获取阳光,植物群落性状分布相对更加均匀,而在下坡位植物为了争取更多的阳光,可能会更多的生长在阳光充足的地方,物种多度分布不均导致FEve的减小[50]。
植物的生存往往也受到土壤因子的影响,土壤条件的变化影响了植物对营养元素的获取策略,并引起植物性状的变异,形成相适应的生存方式[51]。不同的群落功能多样性受土壤因子的影响不同。么旭阳[52]在研究长白山阔叶红松林功能多样性时发现土壤氮含量对植物群落生物多样性影响最大,而土壤磷含量对群落功能多样性影响最大。郑丽婷等[38]在研究庙岛群岛植物群落多样性时认为土壤含水量是影响植物群落最重要的土壤因素。田平等[14]在对太岳山功能多样性研究时发现华北落叶松-白桦混交林灌木层的功能多样性主要受到土壤pH的影响。与上述研究不尽相同,井冈山鹿角杜鹃群落灌木层植物FRic、Rao、FDiv主要受到土壤P的影响,FDis主要受到土壤N的影响。自然条件下生态系统的功能往往受到环境因子和植物功能性状的共同作用[53],海拔梯度、土壤化学成分等环境因子对井冈山地区鹿角杜鹃群落灌木层的功能多样性有着显著的影响,环境因子的变化会引起群落功能多样性的变化。
致谢:耿梦娅、谭向前和张行参加了野外调查,特此致谢。
