干旱胁迫下菊芋各器官生物量及物质分配规律
2019-12-18朱铁霞李志华
朱铁霞,高 阳,高 凯,*,李志华
1 内蒙古民族大学农学院, 通辽 028043 2 扎鲁特旗科学技术信息研究中心, 通辽 028043
全球温度不断升高,必然导致更多地方受到干旱胁迫[1]。干旱胁迫将成为植物生长过程中的主要限制因素之一[2]。植物对干旱胁迫的适应机制和如何提高植物对干旱的抵御能力也一直是生态学者们研究的热点问题,同时也是我国农业和生态文明建设急需解决的难题[3]。
菊芋(HelianthustuberosusL) 为菊科向日葵属植物,又称鬼子姜或洋姜,主要分布在北美温带地区,以块茎繁殖为主,对生境要求较低,具有喜肥耐贫瘠,喜湿耐旱,喜温耐寒,耐盐碱等优点[4-6]。我国主要分布于江苏、山东沿海地区和陕西、甘肃、宁夏等干旱和半干旱地区[7]。近年来,我国菊芋的种植面积逐年增加,尤其在甘肃、宁夏等干旱和半干旱地区。关于菊芋的研究主要集中在耐盐机理[8-9]和如何通过水肥管理、种植密度、收获时间等农艺措施提高其块茎生物产量等方面[10-14]。关于菊芋干旱胁迫适应机制方面的研究相对较少,研究内容主要集中在半干旱地区不同生育期菊芋生长特性与气体交换特征、半干旱地区菊芋品系植株表型与光合特性分析、干旱-复水对菊芋苗期根、茎、叶形态特征的影响、干旱胁迫下菊芋叶片光合变化规律、蔗糖磷酸合成酶及转化酶活性、叶片保护酶活性、膜脂过氧化作用和可溶性碳水化合物的积累及分配规律等相关研究[15-17]。基于上述原因,本研究拟通过对不同干旱胁迫条件下菊芋生物量、茎叶比、根冠比和物质分配规律进行研究,探讨菊芋抗旱生物量机制,为菊芋旱地栽培提供理论依据。
1 材料与方法
1.1 试验设计
试验于2016年5月至2016年9月在内蒙古民族大学农学院试验农场(43°36″N,122°22″E)遮雨棚下进行。采用盆栽方式种植,花盆直径29cm,高度32cm。装盆土壤为灰色草甸土,是当地主要土壤类型,装盆前将土壤混合均匀,每盆装入5.5kg土壤。土壤有机质含量18.23g/kg、碱解氮62.41mg/kg、速效磷38.61mg/kg、速效钾184.56mg/kg、pH值8.2。菊芋品种为当地主要种植的红皮菊芋和白皮菊芋,种植时选取重量为30—40g,无病、无伤的块茎作种,每盆种植3—4枚菊芋块茎,最终保证每盆一株菊芋。
出苗30天每盆保留一株菊芋,并开始进行干旱处理,根据田间持水量(W)按质量百分比设定50%W、40%W、30%W、20%W、10%W共5个水分梯度,每天18∶00用土壤水分测定仪进行水分测定,通过计算后,补充散失水分量,确保土壤含水量为设定梯度。每个梯度15个重复,共计75盆。
1.2 生物量测定
9月30日对每盆菊芋进行齐地表刈割,地上部分进行茎、叶、花和叶柄分离,地下部分进行清洗,及根系和块茎分离。将根、茎、叶、叶柄、花和块茎先在105℃条件下杀青30min,之后在75℃条件下烘干,测定干重。
1.3 相关计算
地上生物量=茎秆生物量+花生物量+叶片生物量+叶柄生物量
地下生物量=根系生物量+块茎生物量
总生物量=地上生物量+地下生物量
根系贡献率=根系生物量/总生物量
茎贡献率=茎秆生物量/总生物量
叶贡献率=叶片生物量/总生物量
叶柄贡献率=叶柄生物量/总生物量
块茎贡献率=块茎生物量/总生物量
花贡献率=花生物量/总生物量
根冠比=地下生物量/地上生物量
茎叶比=茎生物量/叶生物量
1.4 数据分析
采用 Excel 2007和DPS 14.50软件对数据进行统计分析。采用单因素方差分析法检验测定参数之间的差异。
2结果与分析2.1不同灌水处理对菊芋总生物量的影响
图1可以看出,随着灌水量的增加,红皮菊芋和白皮菊芋的总生物量、地上生物量和地下生物量均呈现逐渐增加的变化趋势,50%灌水条件下白皮菊芋和红皮菊芋的总生物量、地上生物量和地下生物量均显著高于其他灌水处理(P<0.05),40%灌水条件下白皮菊芋和红皮菊芋的总生物量、地上生物量和地下生物量均显著高于30%、20%和10%(P<0.05),30%、20%和10%灌水条件下差异不显著。白皮菊芋地上生物量30%灌水条件下显著高于20%和10%(P<0.05),20%和10%之间没有差异;40%条件下红皮菊芋总生物量显著高于白皮(P<0.05),50%和30%条件下白皮菊芋地上生物量显著高于红皮(P<0.05),40%条件下红皮菊芋地上生物量显著高于白皮(P<0.05),其他相同灌水处理条件下白皮菊芋和红皮菊芋的总生物量、地上生物量和地下生物量之间均无差异。
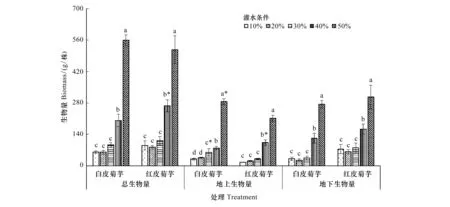
图1 灌水量对菊芋生物量的影响Fig.1 Effects of irrigation on biomass of Jerusalem artichoke 不同小写字母表示相同品种不同灌水量之间在0.05水平下差异显著;*表示相同灌水量不同品种之间在0.05水平下差异显著
2.2 不同灌水处理对菊芋各器官生物量的影响
50%灌水条件下白皮和红皮菊芋的根系生物量、茎秆生物量、叶片生物量、叶柄生物量、花生物量和块茎生物量均显著高于10%、20%、30%和40%(P<0.05);10%、20%和30%三个灌水条件下白皮菊芋的根系生物量、叶片生物量和叶柄生物量之间没有显著差异,且均显著低于40%灌水条件下根系生物量、叶片生物量和叶柄生物量(P<0.05);红皮菊芋根系生物量和叶柄生物量在10%、20%、30%和40%之间没有显著差异,40%灌水条件下花生物量显著高于10%、20%和30%(P<0.05),而10%、20%和30%三者之间没有显著差异;白皮菊芋茎秆生物量30%条件下显著高于10%、20%和40%(P<0.05),40%显著高于10%和20%(P<0.05);红皮菊芋茎秆生物量10%和20%显著低于40%(P<0.05)(表1)。
相同灌水条件下白皮菊芋叶柄生物量均显著高于红皮(P<0.05),红皮块茎生物量均显著高于白皮(P<0.05),10%、20%和30%条件下白皮菊芋根系生物量均显著低于红皮(P<0.05),40%和50%条件下白皮菊芋根系生物量均显著高于红皮(P<0.05),30%条件下白皮茎秆生物量显著高于红皮(P<0.05),20%、40%和50%条件下白皮叶片生物量显著高于红皮(P<0.05),40%和50%条件下红皮花生物量显著高于白皮(P<0.05)(表1)。

表1 不同灌水处理对菊芋各器官生物量的影响
2.3 不同灌水处理对菊芋物质分配规律的影响
白皮菊芋根系贡献率最小值出现在30%灌水条件下,显著低于10%和40%(P<0.05);茎贡献率最大值出现在30%灌水条件下,显著高于10%、20%、40%和50%(P<0.05);叶贡献率20%条件下显著高于10%、30%、40%和50% (P<0.05),10%显著30%、40%和50%(P<0.05);50%条件下花贡献率显著高于30%和40%(P<0.05)。红皮菊芋根系贡献率随控水梯度的增加呈现逐渐降低的变化趋势,10%和20%显著高于30%、40%和50%(P<0.05);茎贡献率呈现先增加后降低的变化趋势,10%条件下最低,其值显著低于20%、30%、40%和50%(P<0.05);叶片贡献率和叶柄贡献率呈现先降低后增加的变化趋势,50%灌水条件下红皮菊芋叶贡献率最高,显著高于10%、20%和30%(P<0.05),;花贡献率呈现先升-降-升的变化趋势(表2);相同水分处理条件下,各器官贡献率因品种不同表现出不同的大小关系。如:10%、20%和30%条件下红皮菊芋根系贡献率显著高于白皮(P<0.05),40%条件下白皮根系贡献率显著高于红皮(P<0.05);30%条件下白皮菊芋茎贡献率显著高于红皮(P<0.05),40%条件下红皮茎贡献率显著高于白皮(P<0.05);白皮菊芋叶片贡献率在不同灌水条件下均显著高于红皮(P<0.05);10%、20%、30%和40%条件下白皮菊芋叶柄贡献率显著高于红皮(P<0.05);30%、40%和50%条件下红皮块茎贡献率显著高于白皮(P<0.05);40%条件下红皮菊芋花贡献率显著高于白皮(P<0.05)(表2)。
2.4 不同水分梯度对不同品种菊芋茎叶比的影响
由图2可知,随着灌水量的增加,白皮和红皮两种菊芋的茎叶比均呈现先增加后降低的变化趋势,30%灌水条件下白皮菊芋茎叶比最大,显著高于其他处理(P<0.05),其他处理之间没有显著差异;20%灌水条件下红皮菊芋茎叶比最大,其中20%、30%和40%灌水条件下茎叶比显著高于10%和50%(P<0.05),20%、30%和40%之间以及10%和50%之间没有显著差异。相同灌水条件下,红皮和白皮菊芋茎叶比仅20%和40%条件下表现出显著差异(P<0.05),其他灌水条件下没有表现出显著差异。
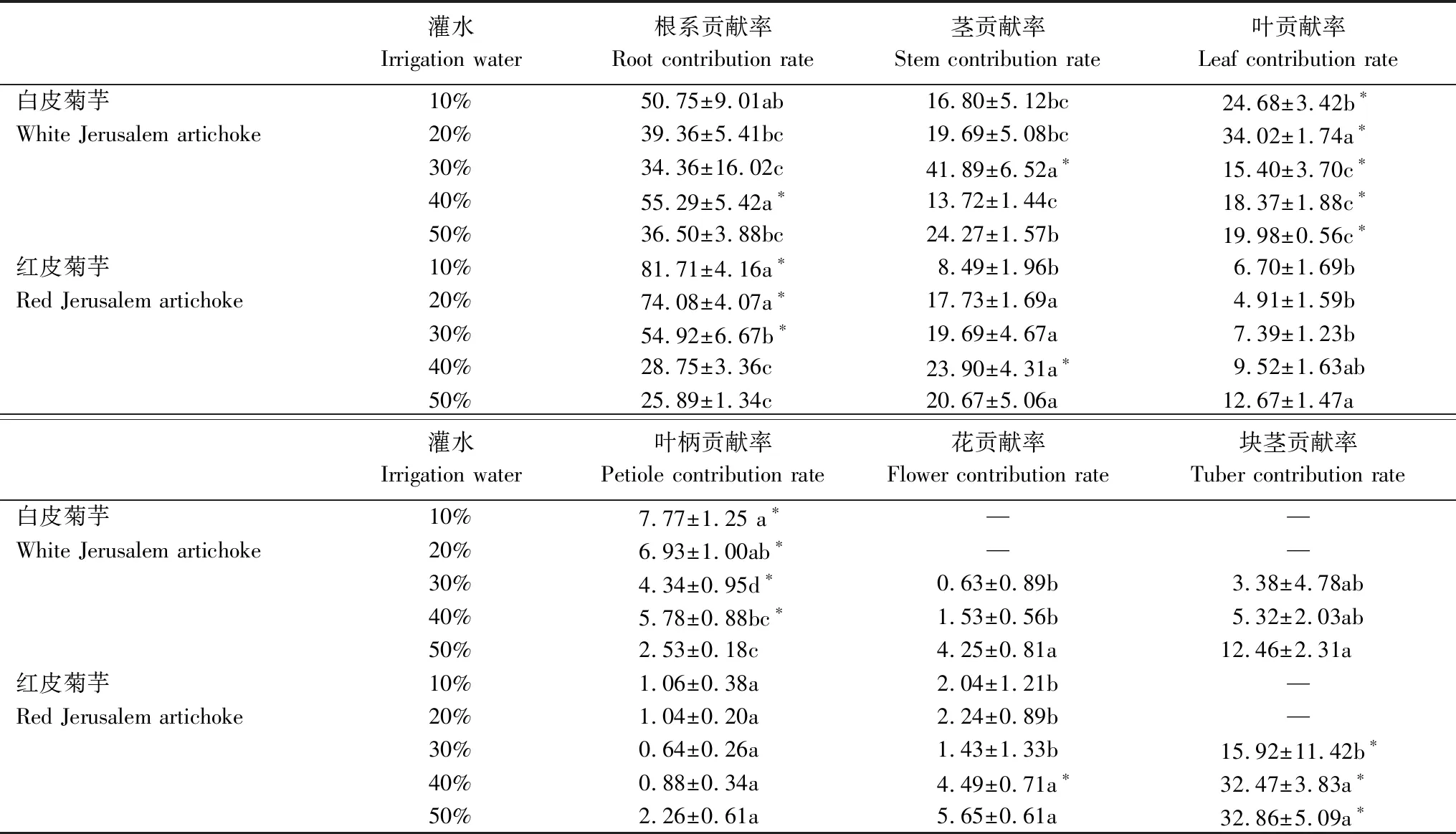
表2 不同灌水处理对菊芋物质分配规律的影响
2.6 不同水分梯度对不同品种菊芋根冠比影响
由图3可知,红皮菊芋根冠比随着灌水量增加呈现逐渐降低的变化趋势,10%条件下根冠比最高,显著高于其他各处理(P<0.05),20%条件下显著高于30%、40%和50%(P<0.05),50%显著低于30%和40%(P<0.05),30%和40%之间没有显著差异;10%、20%和30%灌水量条件下红皮菊芋根冠比显著高于白皮(P<0.05),40%和50%灌水条件下白皮和红皮根冠比之间没有显著差异。

图2 灌水量对菊芋茎叶比的影响Fig.2 Effects of irrigation on stem/leaf ratio

图3 灌水量对菊芋根冠比的影响Fig.3 Effects of irrigation on root/shoot ratio
3 讨论
随着灌水数量的增加,红皮和白皮菊芋各器官生物量均呈现逐渐增加的变化趋势,说明尽管菊芋具有一定的耐旱性,但水分仍然是提高菊芋生物量的重要因素[5,18]。而干旱胁迫对菊芋根系、茎秆、叶片、叶柄、花和块茎生物量降低的原因却有所差异。根系、茎秆、叶片、叶柄生物量随灌水量的降低其减少的原因主要是由于菊芋在生长发育过程中,没有充足的水分供应,影响到光合作用,同化产物合成减少,进而影响了干物质的积累数量和速度,最终表现为各营养器官生物量降低[19-20]。而花和块茎作为菊芋的生殖器官,其形成要滞后于营养器官,其物质的来源也主要是由营养器官所积累的物质向生殖器官运输而获得[21]。因此花和块茎生物量的减少更多原因应该是干旱胁迫通过影响营养器官所导致。
植物干物质分配被认为是植物适应逆境胁迫的重要策略之一[22]。许多研究证明,干旱胁迫条件下,许多植物通过调整自身的物质分配格局,可以有效地提高自身对干旱胁迫的适应性[23-24]。如有的植物通过改变干物质总量及其在根、冠之间的分配比例来提高自身对干旱胁迫的适应性[25-26]。本文中红皮菊芋对干旱的适应性则显示出类似的变化规律,即灌水量从50%降低到10%其根冠比呈现逐渐增加的变化趋势,同时期根系贡献率也呈现逐渐增加的变化趋势,其目的是增加地下根系,从而提高根系的吸水功能,降低地上茎、叶干物质数量,来减少蒸腾作用,进而减少水分的散失。这种通过增加根系而减少地上部分干物质分配的干旱适应机理是植物普遍的干旱适应策略[27-28]。而白皮菊芋则表现出不同的干旱适应策略,这种差异可能是由于遗传所决定。
