具有Allee效应捕食-竞争系统的时空动态分析
2019-12-16刘陈霖郑三强韩晓卓
刘陈霖,郑三强,韩晓卓
(广东工业大学 应用数学学院,广东 广州 510520)
当一个种群的密度减少到一定程度时,种群数量将会维持在一个很低的水平或趋于灭绝,这种现象称之为Allee效应[1-4]. Allee效应的产生机制在于,当种群密度过低时,种群个体将面临近亲繁殖和难以抵御天敌等问题,从而导致出生率减少,且死亡率增加[5-6]. 近几十年来,大量的野外观测和实验研究表明,Allee效应出现在生态系统各个尺度或水平的物种上[7-8],这极大地增加了物种局部甚至全球灭绝的风险[7,9]. 具有Allee效应的局域种群动态及实证研究已因此成为种群生态学关注的热点之一[10]. 已有研究表明Allee效应对捕食系统、竞争系统,甚至集合种群都产生重要影响[11-13]. 例如,Allee效应可以破坏捕食系统原有的稳定性[10,14],并引起种群数量随时间震荡;对于竞争系统,Allee效应也导致其出现明显的不稳定性[12-13]. 所研究的捕食-竞争系统多以Lotka-Volterra模型和Levins模型的集合种群模型为主. 在集合种群尺度上,当斑块的占有率低于一定的阈值时,集合种群则呈现负增长趋势,进而导致集合种群灭绝,该现象被称之为似Allee效应[15]. 似Allee效应直接诱导集合种群的全局密度降低[16]或集合种群内部各局部种群同步灭绝.
尽管Allee效应对局部种群动态的研究为种群生态学关注的热点之一,鉴于之前的研究发现,因为产生Allee效应的先决条件为种群密度过低[5-6],Allee效应产生的作用强度有可能受到种群初始密度的影响,也即初始种群密度与Allee效应之间的关系鲜有报道. 另外,上述研究过多涉及Allee效应指数本身对系统稳定性,特别是种群数量或种群密度的作用关系,而对于该效应是否对物种或种群的空间模式具有一定的调节作用,也未见报道. 因此,基于两点,本文以Kylafis和Loreau[17]提出的捕食-竞争系统为基础,考虑到捕食者的Allee效应,建立具有Allee效应的捕食-竞争系统,并利用相平面分析和耦合映像粒子模型[18-19]分别给出系统的稳定性分析和空间分布模式. 通过MATLAB软件进行数值模拟,探讨Allee效应和捕食者初值的共同作用对系统稳定性及各物种种群空间分布模式的影响.
1 模型
Kylafis和Loreau基于生态位构建理论提出了一个反映生态作用关系与环境之间具有正向反馈作用的捕食-竞争系统模型[17]. 该模型含有一个捕食者P、两个作为食饵的竞争物种A和B,以及固有自然资源R,其中两个竞争物种通过摄取自然资源R生存,捕食者P则以捕食两个竞争物种为生存前提,该模型可表示为
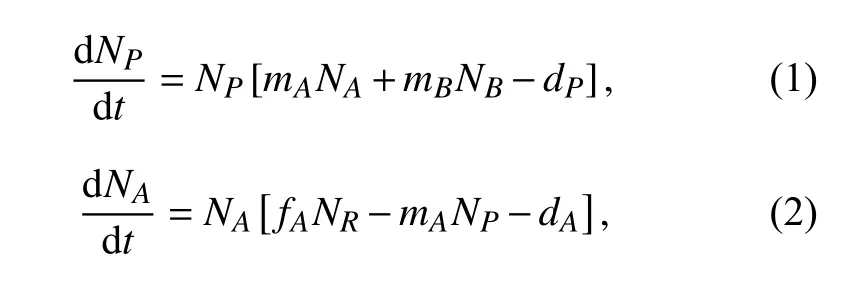
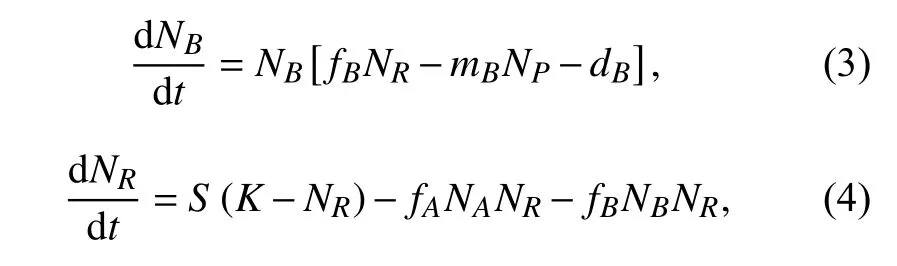
其中NP,NA和NB分别表示捕食者P、物种A和物种B的种群密度,NR为资源浓度;常数mA和mB分别表示捕食者P对物种A和对物种B的捕食率,fA和fB则表示物种A和物种B对资源R的摄取率;dP,dA和dB分别为物种P、物种A和物种B的自然死亡率.K表示资源R的供给总量,S表示资源R的转化再生率. 由方程(1)~(4)组成的系统记为系统(I),该模型详细地阐述了捕食者、被捕食竞争者与资源的关系:捕食者的初值变化不改变系统最终的平衡点,而被捕食竞争者的初值变化只影响其自身的平衡点.
在该捕食-竞争系统中,捕食者作为初始值较小的物种,其受到的Allee效应的影响最大,于是在捕食者上引入Allee效应,则由方程(1)变为如式(5)的捕食者动态方程:
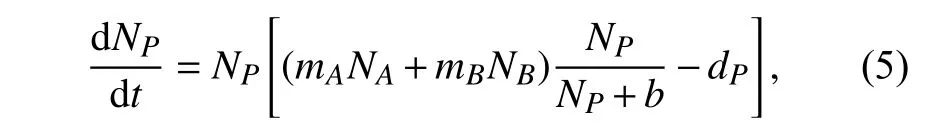
其中参数b为Allee效应强度. 由方程(2)~(5)组成的系统记为系统(II). 显然,当b=0时,系统(II)则退化为系统(I),其他参数含义同上.
本文将采用两种方法讨论捕食者具有Allee效应的捕食-竞争系统的时空动态. 首先,利用数值模拟和分岔图[20]检验Allee效应对于整个系统平衡动态的影响. 然后,利用耦合映像粒子模型对二维异质生境(n×n个斑块)上的具有空间结构的三物种进行空间分布模式的演化仿真. 为此,假设At(i,j)、Bt(i,j)和Pt(i,j)分 别表示在斑块(i,j)上t时刻时被物种A、物种B和捕食者P占据的概率,根据上述系统方程和概率转移模型[21-22]提出如下概率转移规则:

其中,∑At(i,j) ,Bt(i,j) 和∑Pt(i,j)分别为焦点斑块在t时刻时的邻体被物种A、物种B和捕食者P占据的概率之和. 同理,斑块 (i,j) 在t时刻的资源量Rt(i,j)演化动态为
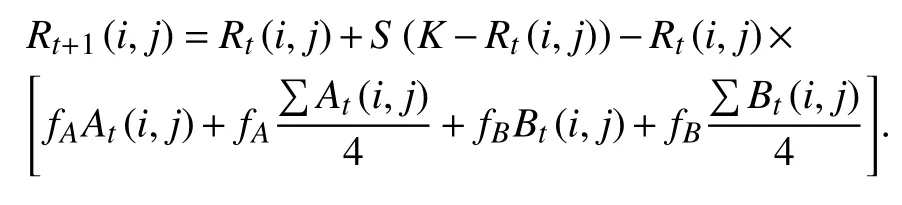
需要说明的是在模拟过程中,三物种初始分布均为随机产生,演化规则采用同步更新、冯诺依曼的临体结构和周期边界,以消除边界效应对结果的扰动.
2 结果
2.1 系统稳定性分析
根据稳定性理论,系统(I)存在一个非平凡平衡点为
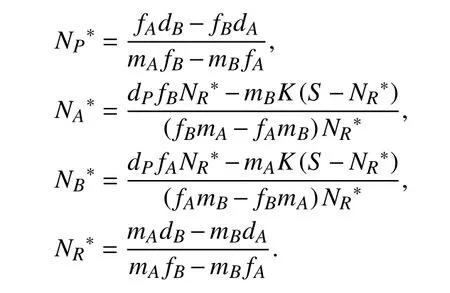
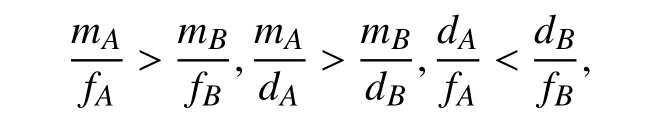
根据系统的生物学意义,当满足条件NP*>0,NA*>0,NB*>0且NR*>0时,意味着资源在不枯竭的前提下使得三物种持续共存. 此时,该系统中的参数关系应同时满足如下条件:即参数的选取遵循捕食者P对物种A的捕食率与物种A对资源R的摄取率的比值大于捕食者P对物种B的捕食率与物种B对资源R的摄取率的比值,捕食者P对物种A的捕食率与物种A的自然死亡率的比值大于捕食者P对物种B的捕食率与物种B的自然死亡率的比值,物种A的自然死亡率与物种A对资源R的摄取率的比值小于物种B的自然死亡率与物种B对资源R的摄取率的比值. 而对于系统(II),根据稳定性理论可得参数关系与系统(I)的参数关系相同.
根据如上参数选取范围,通过MATLAB软件给出系统(I)和系统(II)的三物种相平面图,如图1所示.初值取NP=0.2,NA=0.5,NB=0.5,NR=1. 其他参数取值为mA=0.4,mB=0.35,fA=0.40873,fB=0.36,dP=0.05,dA=0.018,dB=0.018,b=1,S=1,K=1. 由图1可知:两个系统中的3个物种的相空间曲线最终螺旋汇聚于内部平衡点,两个系统最终达到稳定. 在汇聚的过程中,系统(II)(见图1(b))的螺旋圈数远远多于系统(I)(见图1(a)),说明Allee效应导致系统(II)产生持续性阻尼震荡,从而延长了系统达到稳定的时间. 结果表明Allee效应可较大程度上改变系统的时间动态.
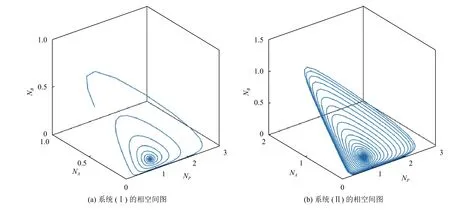
图 1 系统(I)和系统(II)的相空间图Fig.1 The phase space diagram of system (I) and system (II)
2.2 两系统中物种种群密度的时间动态分析
事实上,具有Allee效应的捕食者的初始密度也将对系统产生较大的影响. 在模拟中,固定其他参数不变,观察当捕食者初始密度NP分别取0.1,0.182 和0.25时系统(I)和系统(II)的三物种时间动态图(见图2).其中参数为mA=0.4,mB=0.35,fA=0.40873,fB=0.36,dP=0.05,dA=0.018,dB=0.018,b=1,S=1,K=1,NA=0.5,NB=0.5,NR=1. 显然,捕食者P初值的变化对于无Allee效应的系统(I)无明显影响,捕食者P和物种A、B的种群密度都先呈现阻尼振动,约500时间步长后均达到稳定状态,且均收敛到相同的数值,如图2(a)、图2(c) 、图2(e)所示. 然而,上述结果与系统(II)的三物种时间动态图作对比则发现:捕食者P初值较小时,系统出现无规则的振动状态(见图2(b)),系统无法达到稳定;当捕食者P初值增加时,Allee效应则使系统各物种种群密度产生剧烈的阻尼振动,且震荡的振幅和持续时间都大幅增加(见图2(d));捕食者P初值取值为0.25时,Allee效应仍然引起系统的阻尼震荡(见图2(f)),但相比图2(d),振幅明显减小,且快速收敛到稳定状态. 该结果表明Allee效应可增加系统的复杂性,所产生的结果在一定程度上依赖于初值的选择,这一结果与图1相吻合.
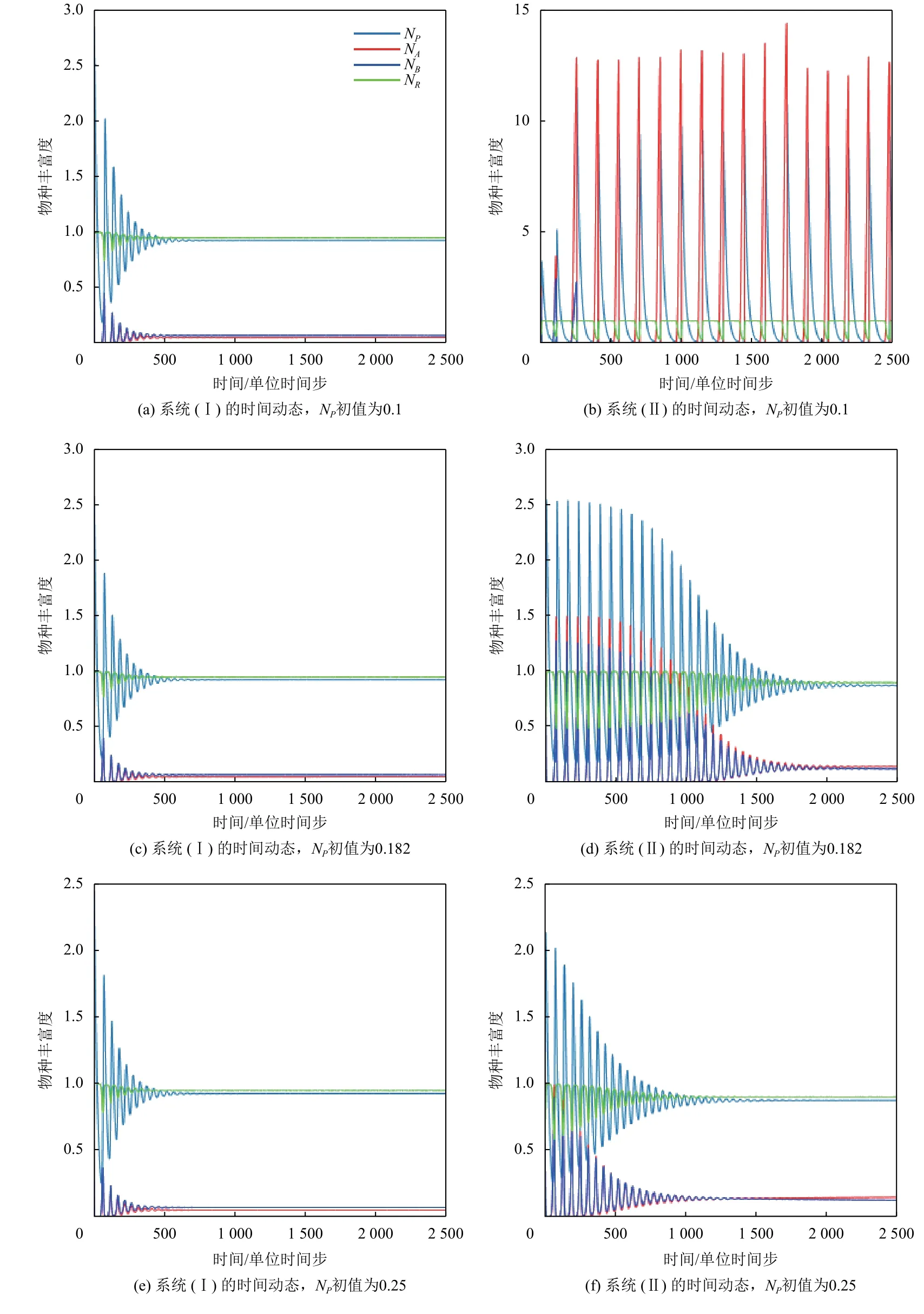
图 2 系统(I)与系统(II)的时间动态Fig.2 The temporal dynamics of system (I) and system (II)
2.3 捕食者初值及Allee阈值对系统稳定性影响
为了更清晰地表明捕食者P初值大小与Allee效应常数对系统稳定性的影响,笔者给出两组不同初值下Allee效应对系统分岔图的分析,如图3所示. 其他参数的取值为mA=0.4,mB=0.35,fA=0.40873,fB=0.36,dP=0.05,dA=0.018,dB=0.018,S=1,K=1. 对于捕食者P初值较大时(NP=0.2),Allee效应常数并未使系统产生分岔现象,并且随Allee阈值b的增加,捕食者P种群密度大幅减少(见图3(a)),而两食饵的种群密度都显著提高(见图3(b)和图3(c)). 对于捕食者P初值较小的情况(NP=0.1),Allee效应则使系统产生明显的混沌现象,特别是随Allee阈值b的逐渐增加,系统越发不稳定(见图3(e)和3(f)). 因此,在下文如无特别说明,模拟中取较大的捕食者初值.
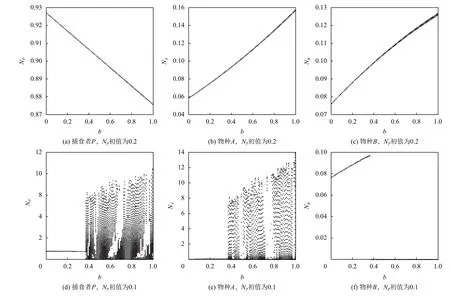
图 3 物种P、A和B的种群密度关于Allee阈值b的分岔图Fig.3 The bifurcation diagram of system (II) regarding Allee threshold b
2.4 系统三物种空间动态分析
图4为系统(I)和系统(II)的空间分布图,参考取值同图3. 从图4可以发现,3个系统处于稳定时的各个物种的空间分布呈现出聚集性[23]分布. 同时发现,在系统(I)和系统(II)中,物种A和物种B的分布呈现出斑块互补的现象. 在空间分布图上,首先以资源层为基础底层,然后在此基础上是以资源为生的被捕食者层,最后是以被捕食者为生的捕食者层. 在被捕食者层中,在物种A的密度较高的地方,物种B的密度较低,这便出现互补的现象. 图5为系统(I)和系统(II)捕食者P第1 200步到第2 000步(间隔为100步)的空间分布图,参考取值同图3. 同一时刻,系统(I)与系统(II)比较,可以清楚地得到具有Allee效应的系统在空间的分布动态上比起不具有Allee效应的系统,呈现出的聚集性分布更加明显.
3 讨论
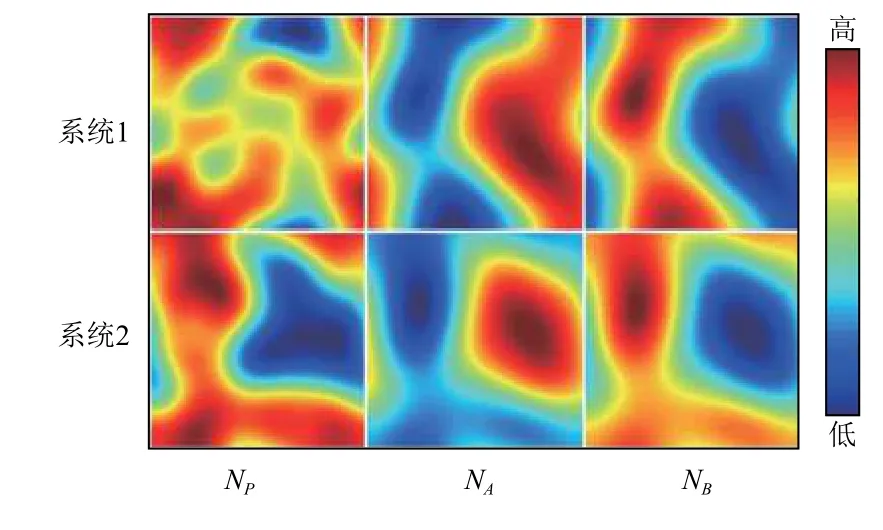
图 4 系统(I)和系统(II)空间分布图Fig.4 Spatial dynamics of systems (I) and (II)
本文以一个捕食-竞争系统为基础,构建了具有Allee效应的捕食-竞争系统,之后对此系统进行稳定性分析和时空动态分析. 结果表明,(1) Allee效应可以使稳定的系统变得不稳定,这具体表现在:当初始时刻捕食者丰富度较高时,Allee效应导致系统各物种种群数量阻尼振动的振幅和持续时间都大幅增加;当初始时刻捕食者丰富度较低时,Allee效应导致系统的某些物种种群数量无规则变动且不会趋于稳定状态,另一些物种甚至面临灭绝风险. (2) Allee效应降低了个体的空间聚集程度,这主要体现在较短的时间尺度上;若时间很长,分散分布的个体会逐渐形成聚集状态.
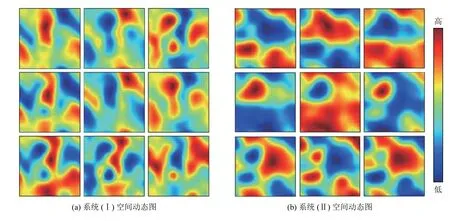
图 5 系统(I)和系统(II)从第1 200步到第2 000步的空间动态(从左到右, 相隔100步), 分别见(a)和(b)Fig.5 Spatial dynamics of system (I) and system (II) from the 1 200 th step to the 2 000 th step are respectively shown in (a) and (b)
对于Kylafis和Loreau提出的捕食-竞争系统[17],韩晓卓与莫昌健[24]的研究表明,生态位构建不会影响系统的稳定性,反而增强了物种的竞争能力. 本文则探讨了Allee效应对系统稳定性和空间分布模式的影响,因此本文的研究有助于进一步了解捕食-竞争系统稳定性的演化机制. 之前关于Allee效应对捕食和竞争系统影响的研究表明,Allee效应破坏了系统原有的稳定性[25-26];本研究虽然也得到了相同的结论,但还表明Allee效应对系统稳定性的影响也受制于种群初始数量:若初始时刻种群数量较高,系统在经过长时间的阻尼振动后仍可趋于稳定状态;但若初始时刻种群数量较低,系统无法达到稳定状态.
本文建立具有Allee效应的捕食-竞争系统,探讨Allee效应和捕食者初值的共同作用的变化对其影响. 研究结果可为保护生物多样性提供理论基础,有助于人们加深对生态系统稳定性的了解,提高人们保护物种的意识. 本研究仅在捕食者上加入Allee效应,但食饵也可能具有Allee效应[11],因此将Allee效应加到食饵上会对捕食-竞争系统造成何种影响有待于进一步解决.
