马铃薯半胱氨酸蛋白酶抑制剂基因(PCPI)在马铃薯晚疫病抗性中的功能表征研究
2019-12-16韦献雅付绍宏2翠3陈志国
史 伟, 韦献雅, 付绍宏2, 阳 翠3, 陈志国
(1.成都农业科技职业学院, 四川 成都 611130; 2.成都市农林科学院, 四川 成都 611130; 3.四川省农业科院经济作物育种栽培研究所, 成都 611130)
马铃薯是第四大粮食作物,在全球粮食安全中发挥着重要作用。然而,对于马铃薯的基因和基因组研究却一直落后于大多数主要作物。
由致病卵菌引起的马铃薯晚疫病是马铃薯寄主(茄属)晚期枯萎病的原因,严重导致世界范围内马铃薯损失[1]。尽管已有针对性的研究,然而对马铃薯晚疫病攻击抗性和易感性的差别尚未完全了解。2011年出版的马铃薯基因组序列提供了新的数据,其对马铃薯遗传学和育种研究有新的见解。另一方面,测序技术的进步使得从研究基因活动的时间和空间分布的大量的组织、时间和空间点中获得完整转录物组(RNA-seq)的深度测序成为可能。基于这些技术,人们对DNA测序的关注已经开始从主要用于生产基因组序列的数据生成工具转移到基础研究和应用研究的功能基因组学工具上。在本研究中,使用对马铃薯晚疫病反应的转录序列,研究四倍体的马铃薯中调节基因对生物胁迫的响应作用。最近,一组分子质量从20到25 kDa不等且来自马铃薯块茎的蛋白质才被报道[2]。其中一些蛋白质被发现是多种多样的昆汀类蛋白酶抑制剂,这些抑制剂可能在马铃薯应对昆虫和植物病原体攻击的自然防御机制中发挥重要作用。它们通常存在于储存组织中,也可在树叶中检测到,以应对多种食草性昆虫和真菌病原体的攻击。例如,在液泡中发现能调动种子萌发时胚乳贮藏蛋白质的植物半胱氨酸蛋白酶。同样,也能在某些物质的胞外介质中找到半胱氨酸蛋白酶,如木瓜和无花果[3]。作为杀虫蛋白,半胱氨酸蛋白酶抑制剂的作用已被报道:奥雷扎·赛斯丁(riceCPI)Ⅰ和Ⅱ是不同品种的豆害虫生长发育迟缓的原因[4]。然而,要直接获得证明基因在决定特定性状作用的证据,必须采用功能基因组学方法。一种是常用于植物基因功能快速分配的转录后基因沉默(PTGS)法[5]。在植物中,细胞质RNA沉默可以通过农杆菌渗入法进行有效诱导,T-DNA载体的瞬时表达的策略。瞬时表达的单个或双链RNA组成的DNA编码,通常是一个发夹(hp)RNA。因为它们提供了一种快速、通用和方便的方法,以实现在叶子的独特和确定的区域达到高水平的基因表达,因此农杆菌介导的瞬态表达系统在诱导沉默过程中发挥了重要作用[6]。通过T-DNA编码hpRNAs(双链RNAs(dsRNAs)来源于反向重复的杂化)的瞬态表达的农杆菌渗入法,能有效诱导瞬态细胞质RNA沉默。与hpRNA转基因的农杆菌渗透实现瞬态RNAi诱导的沉默的方法,已经在尼哥提亚物种、葡萄藤叶、草莓果实、石斛兰的叶子中成功应用[7-9]。利用RNAi结构开发转基因土豆线需要很长时间,因此这项技术只能用于靶向有限数量的马铃薯基因[10-12]。由于瞬态基因检测技术能让基因功能的快速分析具有额外的优势,且可替代稳定转换,同样的细菌也可以被应用到生成稳定的转基因线上。在本研究中,通过农杆菌渗入法生产了针对PCPI的用于测试在马铃薯中获得瞬态基因沉默可能性的dsRNA构造,来确定候选基因在基因调控的马铃薯晚疫病抗性中的作用。
1 材料和方法
1.1 植物材料
所有的实验都是在马铃薯中薯5号上进行。供试植物均在温室中含泥炭苔藓的罐子里栽培(温度23±1 ℃,16 h光周期,湿度80%)。
1.2 转录组分析
为了在下一代测序(NGS)的时间内生成抗性转录组概况,在温室里,抗性品种中薯5号的3份复制品接种了疫病菌隔离种群。使用RNAzol,于不同时间点接种分离mRNA之后,与3个复制品的制备方法一样,制备一个合并的样本。使用NGS,5500 XL固体应用生物系统对分离的mRNAs进行测序,序列读取被组装成片段,归一化,分析折叠变化,根据CLC基因组学工作台4.8(64位)软件读取每百万个碱基的数字。根据与对照组比较的基因的表达水平,选择部分表达水平上调的基因,使用BLASTn,与NCBI数据库中马铃薯基因组序列排成一行。
1.3 总RNA提取、cDNA合成和克隆
在液氮中使冷冻叶片(100 mg)均质化,利用核自旋RNAⅡ试剂盒提取总RNA。
用纳米滴定法测定核酸的浓度和纯度,通过在1%琼脂糖凝胶的电泳实验检查完整性。根据制造商的指示,从纯化的RNA方法合成了cDNA。该产品被用作基因克隆和表达分析的模板。使用聚合酶扩增672-bp对应NM_001318627编码序列(CDS),设计一种克隆引物以扩增如下的CDS:PCPIF:5-TGATGAAGTCGATTAATATTTTGAG-3和PCPIR 5-GCTCC TAC GCC TT GATGAACACAGATG-3。PCR反应条件如下:94 ℃预热4 min,94 ℃加热30 s重复35次,57 ℃加热30 s,72 ℃加热40 s,72 ℃下延长7 min。用1.2%(w/v)琼脂糖TAE凝胶的电泳法分析全部PCR产物。利用pGEM-T Easy载体系统(Promega,美国),将这些PCR产物克隆到TA克隆载体中,然后测序。序列验证了pGEM-T Easy的PCR产物。PCR产物被转化为DH5alpha大肠杆菌,对克隆的质粒DNA测序验证插入情况。序列证明PCR与S.tuberosum的PCPI相同。
1.4 构造hpRNAi载体
PCPI(NM_001318627)基因的全长度cDNA克隆编码用作RNA合成的模板。分析质粒DNA,用于PCR反应的模板。在本研究中,500个基点的片段被选择为RNA沉默的靶点。从克隆的的DNA聚合酶链反应扩增2个500个基点的DNA片段。
设计引物,获得2个PCR产物(“正向”和“反向”片段),随后以相反方向克隆获得发夹构造。用于扩增感官片段的引物是PCPIXhoIF-5′-CGGCTCGAGATGAAGTCGATTAATATTTTG-3,PCPIKpn 1 R 5-AGCGGTACCTCATAATCTTGAAGATGTC-3,分别在它们的5端包含一个Xho1和一个KpnⅠ限制站点(在其中用下划线标注的序列)。用于扩增反义片段的引物为PCPIXba 1 F 5′-GCGTCTAGAATGAAGTCGATTAATATTGAG 3和PCPIClaR 5-ACGATCGATTCATAATCTTGAAGATGTC-3,在5端包括Xba 1和Cla 1位点(序列中用下划线标注)。用50μL总反应容积进行DNA扩增。5μL cDNA与4μL的扩增混合物进行混合,扩增混合物包括每种dNTP 50 mM,每种引物120 nM,1 mM MgCl2,1 U Taq DNA聚合酶和10μL Taq缓冲液。PCR循环如下:在94 ℃时,20 s变性40次,52 ℃下退火30 s,72 ℃下退火45 s,72 ℃时延长退火时间到5 min。这些PCR产品被克隆到PCRTM2.1-TOPO®定向质粒转化为全球前10主管E杆菌细胞(表达载体)。使用相应的限制性内切酶消化DNA,随后配入pKANNIBAL载体,根据花椰菜花叶病毒(CaMV)35 S启动子和章鱼合酶(OCS)终止剂转录控制的适当方向的序列,然后将该结构引进大肠杆菌DH 5 alfa。使用GenElute质粒微型预置组件(Sigma)纯化质粒后,将足底表达的整个表达元素从限制性酶PstI和SacI中切除,并入pCAMBIA 1302载体。通过电穿孔进入ElectroMAXTM根癌土壤杆菌LBA 4404细胞(Invitrogen),引入获得的质粒,用于马铃薯的叶子渗透。
1.5 马铃薯叶子的农杆菌渗入
含有基因构造的农杆菌LBA 4404在50 mL的YM培养基上生长过夜,培养基含5μg·mL-1利福平和10μg·mL-1四环素,因为LBA 4404对利福平和四环素抗生素有耐药性。这些细胞以4 400 r·min的离心率分离10 min,重新悬浮于50 mL MMA培养基(MS盐,10 mM MES,pH=5.6,200μM丁香酮)。调整OD600为0.5~0.8用于瞬态表达研究,并在28 ℃中至少培养2 h。在温室中种植cv(中薯5号)4~5周,然后使用。之后将细菌悬液存于注射器中并渗透到叶的背面。从植物基部,使农杆菌属培养渗透到第3叶柄的3片树叶中。接种组植物,一种为PCIP构造,另一种是野生型pCAMBIA 1302载体。浸润3 d后,从每棵植物中收获1片叶子,在紫外光下观察,然后,通过拍照来检测GFP表达。

注:基因沉默马铃薯叶片(a)的晚疫病症状与对照的叶片(b)感染。图1 疫霉菌感染后7 d的叶片
1.6 分离叶片试验
在添加2%蔗糖的黑麦琼脂培养基中生长疫霉菌感染的avr 1、3、4、7、10、11分离株,时间为10~15 d,温度为15 ℃,以便在培养皿中诱导孢囊形成。为了从孢子囊释放游动孢子,在培养皿中加入了冰冷的水,接着在4 ℃下培养3 h。游动孢子的浓度调整到5×104孢子·mL-1。从植物的上半部分切除在浸润后生长并扩张的叶子,在有50μL的孢子囊悬浮液下端接种。在封闭的玻璃中,将接种的叶子用湿纸巾保存,在培养皿培养(15 ℃,16 h光周期),并每天检查有无损伤。使用数字卡尺(Helios DIGI-MET),在接种后3~6 d测量病变直径。
1.7 定量逆转录PCR(RT-qPCR)
在15 d的渗透时间内,收集被浸润植物的叶子。采集与接种同一阶段的叶子,用于分析PCPI的转录水平。从叶子组织中提取总RNA。使用合成cDNA的方法生成cDNA,用于确定转录水平。遵循制造商的方法,基于SYBR绿色检测系统,使用StepOne定量PCR系统(应用生物系统),由两步测量qRT-PCR技术确定内源性mRNA水平。使用引物表达软件(应用生物系统),设计qRT-PCR引物。用以下引物扩增PCPI的转录:qF:5-ctctttggttgcctttgctc-3,qR(5-cgtttgggcatgtaaggttt-3)。PCR循环条件包括95 ℃下持续5 min的1个周期,95 ℃下持续20 s的45个周期,60°下持续40 s的45个周期。通过与马铃薯伸长因子1 α(EF1A)基因(AB 061263)比较,对相关的基因表达进行归一化。基因(AB 061263)在许多组织中都有表达,是马铃薯中一种有用的内部实时PCR内源线控制。使用形成的熔融曲线确定扩增反应的特异性。对于每个样本,反应进行3次生物复制,每个实验重复3种技术。采用DDCt方法确定与EF1A内源性控制相关的目标基因数量。
2 结 果
2.1 PCPI基因的克隆和RNAi表达载体的构建
在cDNA和基因组DNA(基因库,NM_001318627)中均成功克隆了PCPI基因,并在相应克隆体的测序分析中发现了672 bp羊痘疮(ORF)的存在。基因组复制体中没有内含子,这是一种对抑制蛋白所熟悉的模式。在pKANNIBAL质粒的35 S启动子后,将500 bp的部分序列置于正义和反义取向,最后使用Sac1和Pst1限制克隆酶,在pCAMBIA 1302中克隆。该载体有第2个35 S启动子,驱动GFP转录。农杆菌渗入后的72 h,监测GFP,确定在浸润部位农杆菌介导的质粒的成功转化。
2.2 分离叶片试验
使用马铃薯晚疫病菌株接种的15 d旧沉默叶片,同时选择cv叶子,中薯5号被用作抵抗性参照物,确定PCPI候选基因的沉默是否影响到晚疫病的易感性。接种的叶片进行了3~7 d后期接种(dpi)的视觉观察,测量病变尺寸,绘制病变生长随时间的关系曲线(图1和图2)。在7 d的实验中,没有观察到野生型中薯5号的病变扩张和水浸润,而广泛的水浸润性病变在沉默的植物中很明显。感染后3 d,在野生型叶上出现过敏性反应(HR)病变。在沉默的叶子中,3~7 dpi期间病变尺寸的发展呈稳步增长趋势(图2)。在7 dpi时,沉默叶子上接种部分有明显的芽孢。相比之下,在7个dpi时间内,具有抗性的中薯5号参照植物的叶片上看不到病灶生长,没有检测到孢子。因此,PCPI基因的沉默导致了马铃薯晚疫病抗性的降低。
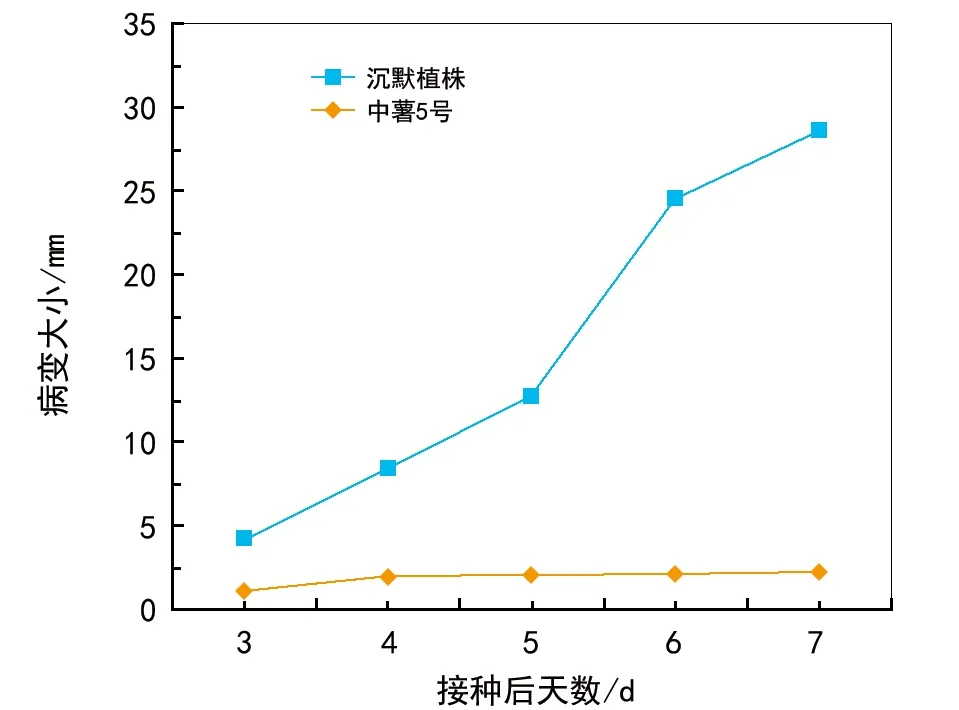
图2 抗性控制cv(中薯5号)和沉默植物的 接种叶片的病灶大小发展
2.3 PCPI转录的表达分析
为了探讨RNAi对靶基因丰度的影响,用RNAi和空构法浸润植物,从一组沉默的和参照的植物中提取出总RNA。结果表明,在沉默的植物中,该基因的转录丰度明显减少,尽管显示不同的降低范围取决于所测试的植物(图3)。总的来说,与非沉默的参照植物对比,在沉默的植物中,PCPI的转录水平降低了87.8%~93.94%(图3)。在15 d的测试中,这些降低的水平仍然很低,表明持续的沉默和对马铃薯晚疫病的敏感性。
3 讨 论
为了提供潜在抗性因子的基因特异性分析,采用基因沉默方法,测试一种特定基因产物参与限制病灶发展的过程。在本研究中,由于结构产物简单、公共供应、均一的植物易感性、绿色荧光蛋白(GFP)的存在,因此选择农杆菌介导转化与载体pKANNIBAL和pCAMBIA 1302联合。此外,在2个载体中,Sac 1和Pst 1 sites的存在保证了克隆的成功。农杆菌渗透测定法被成功应用于筛选N.benthamiana、西红柿、马铃薯、葡萄藤中激活R基因介导的抗病性所需的候选信令组件[13-15]。使用了类似的技术筛选了晚期抗病性所需的候选基因,如PCPI基因。在我们的方法中,通过农杆菌浸润沉默目标基因,将RNAi结构引入抗虫病马铃薯植株中。未沉默抗性中薯5号植物的感染会导致出现超敏表型。如果该基因不参与抵抗,沉默候选基因不会影响超敏反应(HR)表型。如果沉默候选基因(在这种情况下是PCPI)参与马铃薯晚疫病抗性,就会观察到晚疫病的症状。在沉默植物样本中,浸水病灶的分布超过75%被感染的叶面积,而在参照叶感染中没有出现这种情况。由于一周时间沉默效果并不明显,因此GFP表达是初始渗入成功的重要控制方式。
为了证实PCPI在马铃薯晚疫病抗性中的作用,使用包含自我补充的hpRNA构造的内部空间或内含子,提高沉默效率。在本研究中,我们只使用了PCPI(500个基点)编码序列的一部分。这将导致目标基因和任何其他紧密相关的基因沉默。为了检查对马铃薯半胱氨酸蛋白酶抑制剂基因在转录水平时的抑制,我们进行了定量RT-PCR分析,在叶片浸润后第15天,用pCAMBIA 1302携带的RNAi结构,分析内源性转录水平。检测到内源性PCPI的表达水平显著低于参照叶片。然而,从qRT-PCR基量化后观察到的低水平可以得知,PCPI的沉默导致PCPI转录大幅减少,但不会完全消除PCPI转录。残余转录水平是一个重要的因素,通过检测残余转录水平可以决定沉默是否是研究特定基因功能的合适方法。在具有调节功能的蛋白质中,低水平的转录可以直接指导足够的蛋白质合成,使它难以准确评估基因的功能。残留的转录并非是种障碍,一般认为,抑制剂蛋白质和靶向蛋白之间的相互作用,能进入在正常的细胞条件下不会被释放的一种结合状态。
PCPI转录的减少,与在接种了马铃薯晚疫病孢子的沉默植物叶片上存在大量的水浸泡和更快的病灶扩张率有关。这意味着在疫病菌蛋白酶与抑制剂蛋白之间存在相互作用。关于半胱氨酸及其抑制剂介导的防御机制,植物半胱氨酸蛋白酶在程序性细胞死亡诱导的过敏性(HR)反应及病原体攻击调控中发挥重要作用。在此病例中,半胱氨酸蛋白酶的沉默导致敏感性增强,而抑制剂沉默则会增强耐药性。这个假设已经得到验证:半胱氨酸蛋白酶的异位表达抑制诱导的半胱氨酸蛋白酶活性,进而阻断了程序性细胞死亡。另一方面,一些植物病原真菌可产生细胞外蛋白酶,其在参与疾病的发展中起到积极作用。植物蛋白酶抑制剂对植物防御机制的贡献依赖于由微生物产生的蛋白酶的抑制作用。蛋白酶抑制剂的活性是因其具备与目标蛋白酶形成稳定复合物的能力,阻断、改变或阻止进入到酶活性位点。本研究表明,PCPI在抑制由马铃薯晚疫病生产的细胞外半胱氨酸蛋白酶具有重要作用,在抗性品种中薯5号中具有抵抗能力。因此,抑制PCPI基因的结果能增强抗性品种的敏感性,暗示这些抑制剂是马铃薯防御机制应对病原体的一部分。笔者的结果也与几项以前的发现一致,指出植物合成抑制多肽可以抑制酶活性,应对由植物病原微生物产生的蛋白酶的攻击。这一现象首次在感染致病疫霉的番茄中记录,其中增加的胰蛋白酶和糜蛋白酶抑制剂水平与植物抵抗病原体有关。研究显示,马铃薯块茎积累了丝氨酸蛋白酶的蛋白质抑制剂,应对马铃薯晚疫病的感染。此外,荞麦种子的胰蛋白酶抑制剂抑制了植物性真菌中交链孢属替代物和镰刀菌属的孢子萌发和菌丝生长。马铃薯块茎中的胰凝乳蛋白酶抑制剂抑制了晚疫病的生长发育。此外,番茄中半胱氨酸蛋白酶C 14的沉默导致卵菌晚疫病易感性的增加。值得注意的是,尽管真菌疾病作为主要的生物胁迫因子对作物产量有显著影响,但是在植物蛋白酶抑制剂对病原微生物的直接作用的理解程度要比它们对昆虫的影响理解程度小得多。
