丹江口水源涵养区退耕还草对土壤微生物和线虫群落的影响
2019-12-12周广帆杨殿林李青梅赵建宁
周广帆,杨殿林,秦 洁,李青梅,赵建宁
(1. 农业农村部环境保护科研监测所,天津 300191;2. 农业农村部产地环境污染防控重点实验室 / 天津市农业环境与农产品安全重点实验室,天津 300191;3. 沈阳农业大学园艺学院,辽宁 沈阳 110866)
退耕还草是我国重大的生态建设工程。退耕还草后土壤团聚体稳定性增加,土壤结构改善[1],土壤微生物和线虫群落结构改变[2-4],土壤固碳效益提高[5],土壤生态系统功能得以可持续发展[6]。
土壤微生物在物质循环和能量流动过程中起重要作用,其群落结构和多样性与土壤生态系统稳定性关系密切[7-8]。土壤微生物群落结构在不同土地利用类型之间存在着明显的差异[9],植被类型影响微生物群落碳源利用能力[10],种植豆科植物可以增加土壤微生物生物量[11],退耕还草可以改变土壤微生物群落特征[12]。土壤线虫常被用作指示土壤食物网状况和土壤健康的后生动物[13-14],不同的土地利用方式和植被类型改变土壤线虫的群落结构[15-18]。土壤微生物和土壤线虫之间的相互作用构成了土壤微食物网最重要的一部分,影响土壤有机质分解和养分循环[19],从而调节土壤生态系统功能[20]。
丹江口水源涵养区位于南北气候分界的过渡地带,是南水北调中线工程核心水源区、国家级生态示范区和鄂西北国家级重点生态功能保护区,也是国家集中连片特殊困难地区。丹江口水源涵养区生态环境脆弱,80%以上土地属于土石山区和丘陵沟壑区。频繁的人类活动造成了土地退化,退耕还草是其进行生态保护的重要举措。目前针对丹江口水源涵养区的研究大多集中于水土流失,鲜有土壤生态效应相关报道。为此,研究丹江口水源涵养区退耕还草土壤微生物和线虫群落变化特征,探讨土壤微生物和土壤线虫群落相互作用机制,可为全面分析和评估水源涵养区退耕还草的土壤生态效应提供基础数据,也为库区生态环境保护和土地可持续利用提供科学的依据。
1 材料与方法
1.1 研究区域概况及试验设计
试验区位于湖北省十堰市郧阳区,隶属秦巴山区,地处32°25′-33°16′ N,110°07′-111°16′ E,亚热带大陆性季风气候,年均气温15.4 ℃,年均降水量769.6 mm,无霜期244 ~ 255 d。在研究区内选取3个具有代表性的退耕3年的紫花苜蓿(Medicago sativa,MS)草地作为退耕样地,每块退耕样地均选取相邻(< 10 m)长期种植玉米(Zea mays,ZM)的农田作为对照,土壤类型为泥质岩黄棕壤。样地海拔高度、坡向坡位基本一致。紫花苜蓿地和玉米农田均按常规管理:紫花苜蓿年刈割4次,玉米收获后秸秆不还田,紫花苜蓿地施肥量为N 120 kg·hm-2,P2O575.0 kg·hm-2;玉米田施肥量为N 315 kg·hm-2,P2O5100 kg·hm-2。
1.2 土壤样品采集
2017年9月,在每个取样区各布设6个10 m ×10 m的样方,按照“等量、多点、混合”的原则,每个样方内按“S”形选取10个采样点,用内径为5 cm的土钻,按0-10和10-20 cm分层取样,四分法混匀后取1 kg左右的新鲜土样放入无菌自封袋内,及时放入4 ℃冰盒带回,共72份土壤样品。将土样分成3份,一份于室内自然风干待测土壤理化,一份土样保存于-70 ℃冰箱待测土壤微生物,一份新鲜土样用于土壤线虫的分离鉴定。
1.3 土壤微生物磷脂脂肪酸的测定分析
土壤微生物磷脂脂肪酸的提取与分析采用Bligh-Dyer法[21]:取保存于-70 ℃超低温冰箱内的土壤样品在冷冻干燥机内进行冻干处理;取3 g冻干土于特氟龙离心管,加单相提取剂(氯仿∶甲醇∶柠檬酸 = 1∶2∶0.8)水平震荡离心提取总脂,加缓冲液过夜保存,提取下层氯仿相,氮气吹干;经氯仿活化的SPE柱收集磷脂,氮气吹干;将所得磷脂加入甲醇甲苯混合液(1∶1)进行甲酯化,氮气吹干,用正己烷溶解后保存于样品瓶内,然后采用HP6890气相色谱-HP5973质谱联用仪(GCMS)进行分析。测定时,以十九烷酸甲酯(19:00)作为内标,脂肪酸的定量方法采用峰面积和内标曲线法。然后根据不同微生物群落磷脂脂肪酸的分子结构和基团位置,可将其分为放线菌、细菌、革兰氏阳性细菌、革兰氏阴性细菌和真菌[22]。细菌总量以细菌、革兰氏阳性菌和革兰氏阴性菌加和表示,土壤微生物PLFAs总量以检测得到的特征脂肪酸加和表示(表1)。
1.4 土壤线虫鉴定与群落分析
土壤线虫分离采用浅盘法[23]:分离-过筛-冲洗后,加入固定液保存,在解剖镜和体式显微镜下分别进行计数和鉴定。鉴定参照Bongers的分类图[24]和尹文英[25]《中国土壤动物检索图鉴》利用形态学方法鉴定。土壤线虫数量折算成每百克干土中含有的线虫条数。在分析土壤线虫群落时,可根据线虫取食生境和线虫形态学特征将土壤线虫分为食细菌类群(bacterivores,Ba)、食真菌类群(fungivores,Fu)、植物寄生类群(plant parasites,Pp)、捕/杂 食 类 群(predators/omnivores,Op)[26],营养类群均按相对丰度计算。群落结构指数使用通路 指 数(nematode channel ratio,NCR)、富 集 指 数(enrichment index,EI)、结 构 指 数(structure index,SI)来表示[27]。
土壤理化采用实验室常规方法测定[28]。
1.5 数据分析
退耕样地与未退耕样地之间的土壤微生物群落显著性差异(t检验,P < 0.05)采用SPSS17.0分析,用平均值和标准误表示测定结果,运用OriginPro 9.1软件作图;土壤微生物与土壤理化因子和土壤线虫群落之间采用Pearson相关性分析;退耕还草微食物网结构差异分析采用主成分分析,土壤理化因子、土壤微生物和土壤线虫群落三者之间关系采用冗余分析,用Canoco 5软件作图。
2 结果
2.1 退耕还草土壤微生物群落变化
退耕还草土壤微生物群落组成发生改变。退耕种植紫花苜蓿草地中,共检测出44种生物标记的特征磷脂脂肪酸,在对照玉米田中,仅检测到42种特征磷脂脂肪酸。在与未退耕农田相比,退耕种植紫花苜蓿增加了土壤微生物群落磷脂脂肪酸含量。0-10 cm土层中,土壤微生物PLFAs的总量增加了59.13% (P < 0.05),放线菌PLFAs含量增加了64.35% (P < 0.05),细菌PLFAs含量增加64.34%(P < 0.01),真 菌PLFAs含 量 增 加135.19% (P <0.01),革兰氏阳性菌PLFAs含量增加68.88% (P <0.05),革兰氏阴性菌PLFAs含量增加85.83% (P <0.01);10-20 cm土层中,土壤微生物PLFAs的总量增加了62.58% (P < 0.05),真菌PLFAs含量增加100.00% (P < 0.05),革兰氏阳性菌PLFAs含量增加71.05% (P < 0.05),细菌PLFAs、放线菌PLFAs和革兰氏阴性菌PLFAs含量分别增加52.18%、48.62%和57.92%,但均未达显著差异(P > 0.05) (图1)。在退耕还草样地和未退耕农田中,均以细菌PLFAs含量占比最高,土壤中真菌PLFAs含量增长较快,真菌/细菌比值有增加趋势,但未达显著性差异,退耕还草对土壤真菌群落的影响高于细菌群落(图1)。

图 1 退耕还草土壤微生物PLFAs特征Figure 1 Phospholipid fatty acid (PFLA) characteristics of the soil microbes during the conversion of cropland to grassland
2.2 退耕还草条件下土壤微生物与理化因子之间的关系
退耕还草土壤微生物群落特征脂肪酸含量与土壤理化因子存在显著相关关系(P < 0.05)。土壤放线菌、细菌、革兰氏阳性菌、革兰氏阴性菌、微生物PLFAs总量与土壤含水量、硝态氮极显著正相关(P < 0.01),与土壤pH和C/N比极显著负相关(P < 0.01);土壤真菌PLFAs与土壤C/N比显著负相关(P < 0.05);土壤放线菌、细菌、革兰氏阳性菌、革兰氏阴性菌、真菌和微生物PLFAs总量与土壤有机碳和全氮含量极显著正相关(P < 0.01)(表2)。F/B与土壤含水量显著负相关(P < 0.05),与土壤pH显著正相关,与土壤有机碳、全氮、硝态氮含量极显著负相关(P < 0.01)。

表 2 土壤微生物群落与土壤理化因子的相关性Table 2 Correlation between the soil microbial community and soil physical and chemical factors
2.3 退耕还草土壤微生物和土壤线虫群落的相互作用
退耕还草土壤微生物群落和土壤线虫群落相互作用。土壤微生物PLFAs含量与土壤线虫数量之间极显著正相关(P < 0.01),其中革兰氏阴性菌与食细菌线虫负相关(P < 0.05),与食真菌线虫正相关(P < 0.05)。细菌总量与通路指数显著负相关(P <0.05)。革兰氏阴性菌与通路指数极显著负相关(P <0.01),与土壤线虫富集指数和结构指数显著正相关(P < 0.05) (表3)。
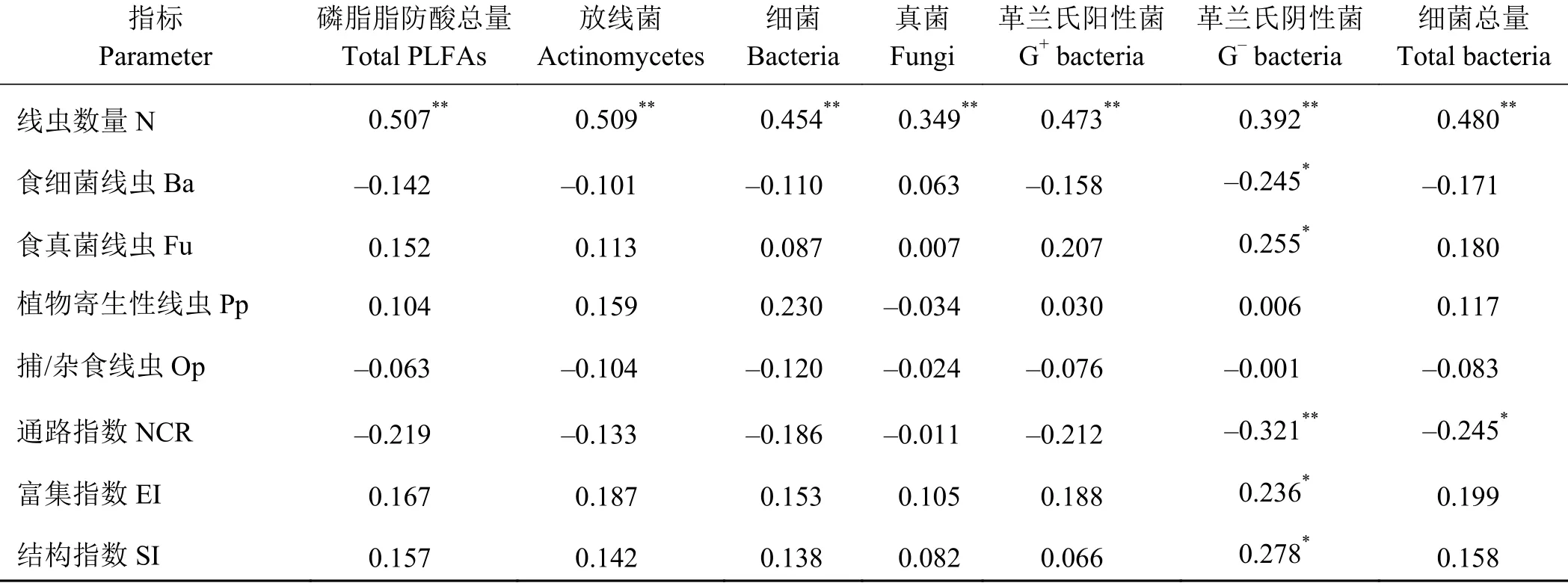
表 3 土壤微生物群落与土壤线虫的相关性Table 3 Relationship between the soil microbial community and soil nematodes
2.4 退耕还草土壤环境因子变化对土壤微食物网的影响
主成分分析表明,退耕还草以土壤微生物和土壤线虫群落为主的土壤微食物网结构发生明显的变化(图2)。PC 1和PC 2共解释了总变异的94.53%,其中Axis 1解释了总变异的89.86%,Axis 2解释了总变异的4.67%。退耕还草样地和未退耕农田在PC 1上明显分离,退耕还草土壤微食物网结构发生分异。

图 2 退耕还草土壤微生物和线虫结构的主成分分析Figure 2 Principal component analysis of the soil microorganisms and nematodes in the conversion of cropland to grassland
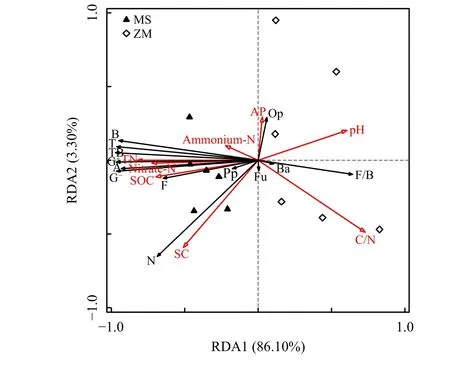
图 3 土壤理化因子与土壤生物群的冗余分析Figure 3 Redundancy analysis between the soil properties and soil biota
土壤环境因子与土壤微生物和线虫群落冗余分析(图3)表明,RDA排序图的典型轴1和轴2分别解释退耕还草土壤微食物网的总体变异的86.10%和3.30%,其中pH (P = 0.046),土 壤 有 机 碳(P =0.018),硝态氮(P = 0.008),全氮(P = 0.002),C/N(P = 0.008)对土壤微生物和线虫群落产生显著影响,表明退耕还草土壤环境因子改变了土壤微食物网结构。从RDA排序图上可以看出,土壤环境因子对土壤微食物网结构差异具有较大贡献。土壤有机碳、全氮和硝态氮含量与土壤微生物群落正相关;土壤pH和C/N与土壤微生物群落负相关,表明土壤微生物群落受到土壤环境因子的调控作用极强。土壤线虫群落与土壤微生物相比对土壤微食物网稳定性贡献更大。土壤微生物、细菌、真菌、放线菌与植物寄生性线虫数量和土壤线虫数量正相关,之间存在极强的相互作用,土壤食细菌线虫与土壤微生物群落负相关,但与细菌/真菌比正相关,表明土壤食细菌线虫的捕食作用改变了土壤微生物群落结构。退耕还草通过改变土壤环境因子影响土壤微生物和线虫群落变化,进而稳定了土壤食物网结构。
3 讨论与结论
土壤微生物群落结构和多样性受植被、土壤环境因子和凋落物的影响[29-31]。退耕还草通过植被、耕作和管理方式的变化对土壤微生物群落结构产生一定的影响。本研究通过PLFA法测定土壤微生物群落发现,丹江口水源涵养区退耕还草提高了土壤微生物磷脂脂肪酸含量,改变了土壤微生物群落结构,与多数研究结果一致[3,12,32],这是因为玉米长期种植导致土壤微生物总磷脂脂肪酸含量较低[33],而紫花苜蓿具有极强的固氮能力,根系分泌物增加,不仅提高土壤养分含量,也为土壤微生物提供了更多的碳源,土壤微生物增加[34]。退耕还草土壤微生物群落中真菌比细菌有较高的增长速率,可能是因为土壤真菌和细菌偏好利用具有不同碳氮比的有机物。退耕还草土壤微生物群落与碳、氮养分正相关(P < 0.05),土壤微生物促进碳、氮循环过程,在物质循环和能量流动过程中起重要作用,提升土壤生态系统服务功能[35]。土壤pH和C/N是表征土壤资源有效性的主要指标[36-37],土壤pH和C/N降低与土壤微生物PLFAs负相关[37-38],可能是因为pH的改变,土壤有机物分解过程发生变化,产生具有不同碳氮比的有机物质,食物资源的改变引起土壤微生物结构变化,但土壤微生物对不同有机物质的利用能力还有待进一步研究。退耕还草显著影响土壤微生物群落结构,有效的土壤管理措施和科学的利用方式有助于恢复土壤生态功能[39]。
退耕还草条件下土壤微生物群落和线虫群落之间极显著正相关(P < 0.01),这是因为土壤微生物数量的增加,为土壤线虫提供了丰富的食物资源,从而线虫数量增加,而土壤食微线虫对微生物的适度捕食会促进土壤微生物数量和多样性增加[40-42],这是一个相互作用的过程。表示食微线虫结构比例的通路指数(NCR)与土壤细菌总量、革兰氏阴性菌显著相关(P < 0.05),革兰氏阴性菌增加了食真菌线虫数量,降低了食细菌线虫数量,改变食微线虫结构,表明土壤线虫食微结构受到土壤微生物的调控。富集指数一般用于评估食物网对可利用资源的响应,而结构指数可以较好地指示干扰程度和食物网的变化[43]。土壤中革兰氏阴性菌数量与土壤线虫富集指数和结构指数正相关,说明退耕还草提高了土壤革兰氏阴性菌的活性,增加了可利用资源的有效性,降低了土壤受到的干扰程度,土壤微食物网更稳定。土壤微生物和土壤线虫之间的捕食关系构成了土壤微食物网最重要的一部分,影响土壤有机质分解和养分循环,增加了土壤可利用资源的有效性[44],从而促进土壤生态效应[20]。
土壤微食物网结构的改变能够影响土壤生态系统功能[45],植物根系及其分泌物是土壤微食物网的主要物质和能量来源[46],退耕还草会引起土壤微食物网的结构变化。退耕还草土壤微食物网结构发生分异。冗余分析表明土壤环境因子的变化对土壤微食物网产生显著影响。退耕还草土壤碳、氮养分和pH对土壤微生物和土壤线虫群落的改变有极大的贡献,这与Maaroufi等[47]研究结果类似,土壤碳、氮为土壤微生物提供了丰富的食物资源,土壤微生物反过来又可以分解和促进土壤养分循环。植物寄生性线虫与土壤微生物群落正相关,土壤中食微线虫结构受到了土壤中C/N的调控,且真菌/细菌显著影响了土壤食细菌线虫和植物寄生性线虫,说明退耕还草改变了植物群落和土壤养分条件,植物通过释放大量的根系分泌物,不仅向微生物提供资源,还向植物寄生性线虫提供资源[48],从而调控土壤微生物活性和土壤线虫结构。在土壤微食物网中,与土壤微生物相比土壤线虫群落稳定性更高,这是因为土壤线虫分布于土壤微食物网中多个营养级,对土壤环境因子变化具有缓冲作用。土壤线虫通过与土壤微生物的相互作用,加速凋落物分解和养分释放[49],促进了土壤碳氮养分积累和转化。土壤微生物和线虫群落还会影响土壤的代谢墒和有机碳库[4,50],土壤环境因子与土土壤微生物和线虫之间的相互作用还有待进一步研究。
综上,丹江口水源涵养区退耕种植3年紫花苜蓿草地土壤微生物群落结构发生改变,多样性增加,但真菌/细菌无显著性差异。土壤微生物与土壤碳氮养分正相关(P < 0.05),与土壤pH和C/N负相关(P < 0.05),退耕还草促进了土壤碳氮养分的积累与转化。土壤微生物和土壤线虫相互作用,土壤革兰氏阴性菌与土壤食细菌线虫负相关,与食真菌线虫正相关,改变了土壤食微线虫结构。土壤微生物与土壤线虫富集指数和结构指数显著正相关(P < 0.05),表明退耕还草土壤微生物和线虫的相互作用促进了土壤资源可利用性增加,降低了土壤受到的干扰,提高了土壤食物网稳定性。退耕还草土壤微食物网结构发生分异,环境因子对土壤微生物群落调控作用要大于土壤线虫群落,土壤线虫对土壤微食物网结构稳定性贡献更大。
