模拟氮沉降对青海湖流域高寒草原植物群落组成及稳定性的影响
2019-12-12董世魁赵珍珍韩雨晖刘世梁董全民周华坤土旦加祁星民王浩善
张 静,董世魁,赵珍珍,李 帅,韩雨晖,沙 威,沈 豪,刘世梁,董全民,周华坤,土旦加,祁星民,王浩善
(1. 北京师范大学环境学院,北京 100875;2. 北京大学深圳研究院,广东 深圳 518000;3. 青海大学青海省畜牧兽医科学院,青海 西宁, 810003;4. 中国科学与西北高原生物研究所,青海省寒区恢复生态学重点实验室,青海 西宁 810003;5. 青海省铁卜加草原改良试验站,青海 共和 813000)
在不断加剧的人类活动以及气候变化的影响下,全球陆地生态系统的氮输入量持续增加[1],使得氮沉降成为近几十年来备受关注的全球变化问题之一。氮是限制植物生长的关键营养元素,氮沉降可以补充土壤养分,促进植物生长,但持续增加的氮沉降会引起土壤酸化、高寒草甸植被和土壤碳氮再分配[2],改变植物群落结构及物种多样性[3-4]。随着人类活动加剧,青藏高原地区的大气氮沉降十分明显,1990-2003年氮沉降量在8.7~13.8 kg·(hm2·a)-1,并呈逐年增加趋势[5-6]。氮沉降的持续增加明显改变了陆地生态系统的碳、氮循环,进而影响整个生态系统的结构、功能及稳定性[7-8]。群落稳定性是群落各组分受到干扰后保持平衡状态的能力,关于群落稳定性的研究主要集中在多样性-稳定性关系上,其关系大致可以分为正相关、不相关和负相关3种。有研究结果表明,物种多样性高的群落,其抵抗力强,群落稳定性高,但May用数学方法推导出群落稳定性与物种多样性负相关,群落稳定性随物种多样性的增加而降低,也有学者认为群落稳定性与物种多样性之间不存在简单的相关关系[9-11]。因此,明确氮沉降对高寒草地植物群落组成及稳定性的影响对于积极应对全球氮沉降量增加具有极大的现实意义。
目前,青藏高原高寒草地氮添加试验大多从物种对可利用氮素增加的响应角度,研究施肥条件下植物群落结构、植物多样性、草地生产力等的变化,发现不同功能群的物种对氮添加的响应不同,施肥显著增加了禾本科植物的盖度、生物量和重要值,降低了豆科和杂类草的盖度、生物量和重要值,莎草科植物变化不明显或有所增加,施肥量增加到一定程度后各功能群的盖度、生物量、重要值和物种多样性均显著降低,导致群落生物量,物种组成和物种多样性改变[12-14]。高寒草地群落的稳定性研究主要集中在围栏封育,不同修复年限对群落稳定性的影响等方面[9,15],关于氮沉降背景下,高寒草原植物群落的稳定性研究较少。基于此,提出科学假设“氮沉降会改变高寒草原群落中不同功能群物种的组成,降低物种多样性,从而改变群落稳定性”,为了验证这一科学假设,选择青海湖高寒草原区为研究样点,基于青藏高原氮沉降现状模拟氮沉降梯度,探究青藏高原高寒草原群落组成、物种多样性及生态系统稳定性对氮沉降的响应模式,为解决未来持续氮沉降带来的生态环境问题提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于青海省海南藏族自治州共和县铁卜加草原改良试验站(37°02′ E,99°35′ N),海拔3 270 m。该区属典型高原大陆性气候,日照时间长,夏凉冬寒,年均气温在0 ℃左右,年最低温和最高温分别为-11.17 ℃和11.57 ℃,无绝对无霜期。年降水量为377 mm,年蒸发量在1 484 mm左右。该区草地类型属高寒草原,以草地早熟禾(Poa pratensis)、短芒披碱草(Elymus breviaristatus)、猪毛蒿(Artemisia scoparia)为主要建群种,物种数在17种左右,土壤以栗钙土和暗栗钙土为主[6,16]。
1.2 研究方法
1.2.1 样地设置及样方调查
2014年,在铁卜加草原改良试验站的围封高寒草原上设置了200 m × 200 m的样地,并在样地内划分18个2 m × 5 m的长方形小区,为避免不同处理间的干扰,小区之间设置1 m的隔离带。根据Lu和Tian[6]文章中所述的青藏高原年平均氮沉降量[8 kg·(hm2·a)-1]设置氮倍增试验,设CK、NⅠ、NⅡ、NⅢ、NⅣ和NⅤ共6个处理,氮沉降量分别为0、8、24、40、56和72 kg·(hm2·a)-1,每 个 处 理3组重复,共18组,将其完全随机布设在长方形小区中。综合考虑氮沉降量为累积过程以及野外试验的可行性,将不同处理所需的氮量折算为需要施加的硝酸铵(NH4NO3)量,并将全年氮添加量均分为两次,2015年和2016年分别在植物生长初期(5月中旬)和生长旺期(7月中旬)的傍晚(19∶00左右)采用土施法各施加一次硝酸铵(NH4NO3)模拟氮沉降。
2016年7月下旬在各小区随机布设一个1 m × 1 m的植物样方,共调查18个样方,并记录每个样方内出现的物种名称以及各物种的频度(针刺法)、盖度(目测法)、多度(样方内各物种的数目)等。
1.2.2 植物物种重要值
重要值是以综合数值来表示群落中不同物种的相对重要值,能较充分地显示出不同物种在群落中的优势地位,重要值 = (相对频度 + 相对盖度 +相对密度)/3。式中:相对频度为某一物种的频度占全部物种频度之和的百分比;相对盖度为某一物种的盖度占全部物种盖度之和的百分比;相对密度为某一物种的个体数占全部物种个体数目的百分比[17]。
1.2.3 物种多样性指数
物种多样性分析选取3种常用的物种多样性指数,即Patrick丰富度指数(R0)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)进行分析[18],具体公式如下:
Patrick丰富度指数:R0= S;

式中:S为群落内物种总数,Pi为第i个物种的个体数在群落个体总数中的比例。
1.2.4 植物群落稳定性测度方法
根据Godron稳定性测定方法[19],结合郑元润[20]对Godron稳定性测定改进的数学方法,计算出各施氮处理下的群落稳定性。首先将研究群落中各物种的频度由大到小排列,其次将整个群落的物种总数取倒数,按照植物种类排列的顺序计算总种数倒数的累积百分数,并将各物种的频度换算为相对频度按照植物种类排列的顺序计算相对频度的累积百分数,最后对二者做散点图,建立数学模型得出平滑曲线,同时做直线y = 100-x,其与平滑曲线的交点即稳定性参考点(x,y)。交点坐标越接近稳定点(20,80),群落越稳定,反之,越不稳定。因此,本研究采用欧式距离来表示群落的稳定性[21]。
平滑曲线模型为:

直线方程为:

将方程(2)代入方程(1)得:

解方程得:

求出两个交点坐标后根据研究方法,只保留x值小于100的交点坐标,即为所求的稳定性参考点(x,y),继而计算(x,y)与(20,80)的欧氏距离来表示群落稳定性。
1.3 数据分析
基础数据进行整理和分析后,计算物种多样性指数和群落相似性系数,采用SPSS19.0 (IBM,Armonk, NY, USA)数据统计软件对不同实验处理下的物种多样性指标进行单因素方差分析(One-Way ANOVA),用Duncan检验进行不同氮添加处理间植物群落组成以及Patrick丰富度指数、Shmnnon-Wiener多样性指数和Pielou均匀度指数的多重比较(P < 0.05),用Pearson相关分析不同氮添加处理下植物群落稳定性和物种多样性的关系,最后利用Origin 9.4软件(Origin Lab Corporation, Northampton,MA, USA)制图。
2 结果与分析
2.1 不同氮添加处理对高寒草原植物群落物种组成的影响
通过比较不同氮添加处理下功能群和群落优势种的重要值,发现随着施氮量增加,群落物种组成发生了改变(表1)。在功能群水平上,NⅠ、NⅡ、NⅢ、NⅣ、NV处理下禾本科和杂类草植物的重要值均与CK间无显著差异(P > 0.05),但NⅤ处理下禾本科植物的重要值最高,占群落总重要值的79.3%,显著高于NⅠ、NⅡ、NⅢ处理。杂类草的重要值则在NⅠ处理下最高,NV处理下最低,少于CK重要值的一半(P < 0.05)。不同氮添加处理下莎草科植物的重要值与CK相比增加了113.72%~280.39%,但与豆科植物相同,不同处理间的重要值差异不显著(P > 0.05)。
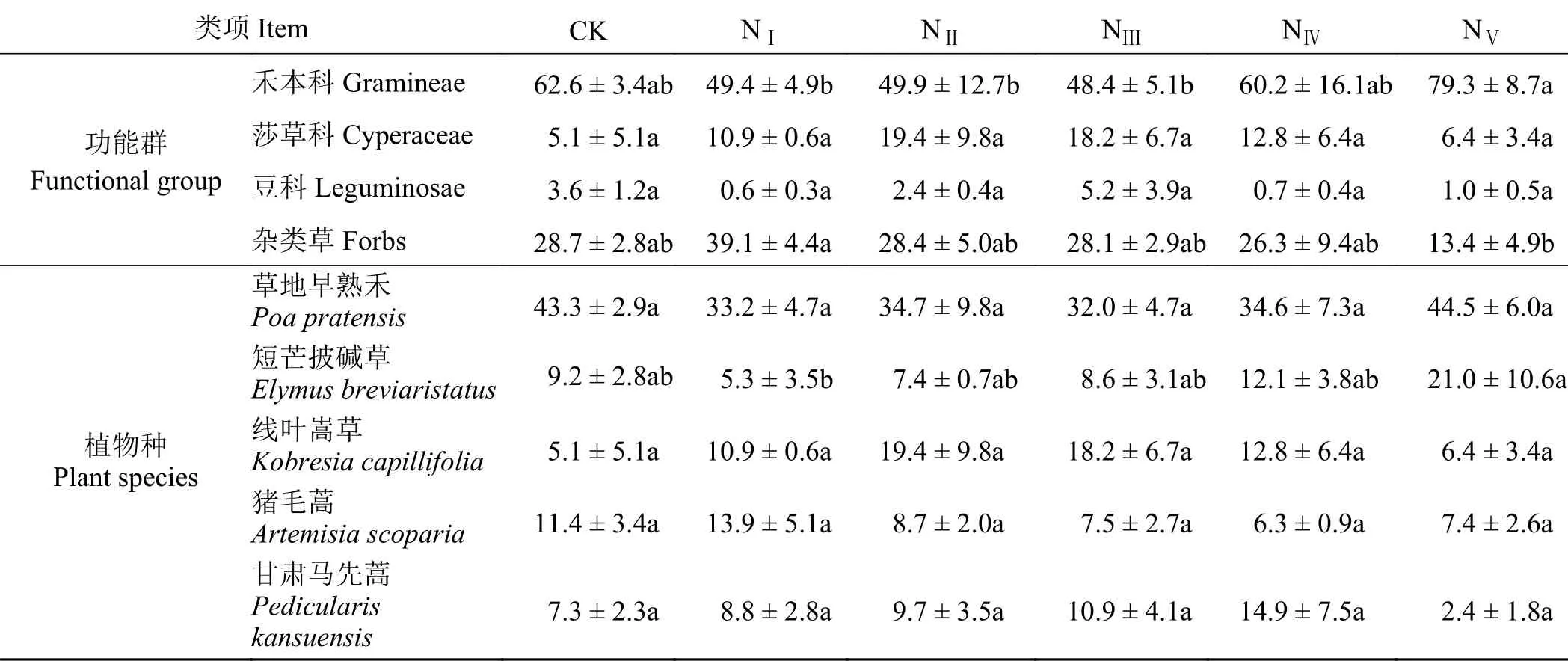
表 1 不同氮添加处理下群落的物种组成及其重要值Table 1 Species compositions and importance values of plant communities at different nitrogen levels
在物种水平上,主要分析了重要值较高的物种在不同施氮处理下的重要值变化规律。不同施氮处理下草地早熟禾始终是优势种,其重要值占群落总重要值的32.0%~44.5%。NⅣ和NⅤ处理下短芒披碱草的重要值明显增加,分别是CK处理的1.32和2.28倍。NⅠ、NⅡ、NⅢ、NⅣ处理下线叶嵩草(Kobresia capillifolia)的重要值先升高后降低,甘肃马先蒿(Pedicularis kansuensis)的重要值呈现随施氮量增加而增加的趋势。除NⅠ处理外,与CK相比各施氮处理下猪毛蒿的重要值均有所降低,降幅最大为44.74%,但均未达显著水平(P > 0.05)。
2.2 不同氮添加处理对高寒草原植物群落物种多样性的影响
不同氮添加处理对高寒草原群落物种多样性的影响如图1所示,少量的氮素添加促进了NⅠ处理的Patrick丰富度指数,最高值为14.5,最低值出现在NⅤ处理,相比于NⅠ处理显著(P < 0.05)降低了33.31%,但各施氮处理均与CK处理无显著差异(P > 0.05)。NⅢ和NⅣ处理的Shannon-Wiener多样性指数高于CK处理,而NⅤ处理的Shannon-Wiener多样性指数显著(P < 0.05)低于CK处理,降幅为13.47%。不同施氮处理下的Pielou均匀度指数的变化趋势与Shannon-Wiener多样性指数相同,在NⅢ和NⅣ处理下增加,在NⅤ处理下呈下降趋势,但各施氮处理之间均无显著差异。
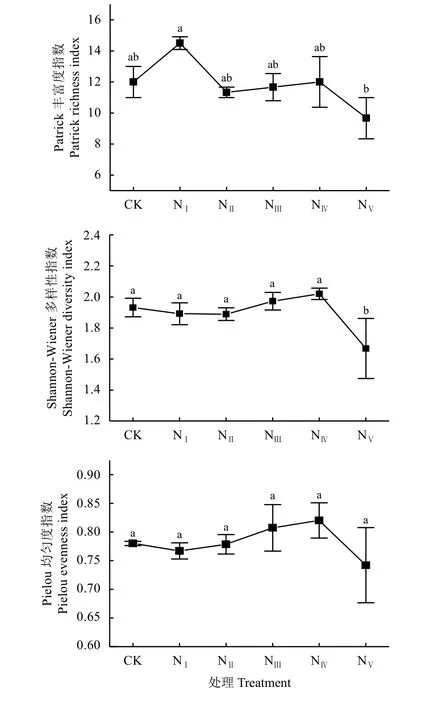
图 1 不同氮添加处理下高寒草原群落物种多样性变化Figure 1 The change in species diversity index of alpine steppes under different nitrogen levels
2.3 不同氮添加处理对高寒草原植物群落稳定性的影响
对不同氮添加处理的物种累计百分数和累积相对频度百分数进行曲线拟合(图2),拟合曲线的R2> 0.9,拟合效果较好(表2)。各处理交点坐标与稳定点(20, 80)的欧氏距离分别为12.30、13.32、10.99、15.53、14.23和8.97,高寒草原研究区群落稳定性的判定结果为NⅢ、NⅣ处理的稳定性距离明显大于CK、NⅠ和NⅡ处理,即NⅢ、NⅣ处理的施氮量会导致群落稳定性降低,但NⅤ处理的植物群落与其他处理相比更加稳定。这与NⅤ处理下优势物种重要值占比增加有关,草地早熟禾和短芒披碱草的重要值总和占群落总重要值的65.5%,其相对频度累积分别为47.17%和68.77%,决定散点图的前两个点,使得NⅤ处理的拟合曲线高于其他处理下的拟合曲线,稳定性参考点更加接近稳定点(20, 80) (图2)。Pearson相关分析表明,不同氮添加处理下植物群落稳定性与Shannon-Wiener多样性指数和Pielou均匀度指数呈显著负相关关系(P < 0.05),相关系数分别为-0.872和-0.839,表明NⅤ处理下群落稳定性较高与群落内Shannon-Wiener多样性和Pielou均匀度下降有关。

图 2 不同氮添加处理下的稳定性拟合曲线Figure 2 M. Godron stability simulated curves of different nitrogen levels

表 2 不同氮添加处理下高寒草原群落稳定性分析Table 2 The stability of alpine steppes under different nitrogen levels
3 讨论
物种组成是生态系统稳定性、生产力、营养动态等功能的重要决定因子,而物种多样性是群落功能复杂性和稳定性的重要度量指标[22]。因此,物种组成和物种多样性的变化是外界环境变化对群落结构和功能的影响的直观体现。本研究中,不同施氮处理下功能群重要值与CK处理相比虽无显著变化,但与NⅠ处理相比,NⅤ处理下的施氮量明显改变了植物群落内功能群的重要值,主要表现为禾本科植物的重要值增加,杂类草的重要值降低,群落内物种数减少,莎草科和豆科植物重要值无明显变化。这与在贝加尔针茅草甸草原进行的增水增氮研究结果相似,即在年际变化中增氮对多年生根茎禾草有促进效应,对多年生杂类草有抑制效应,群落结构发生改变[23]。同时,有学者指出,不同功能群(禾草类、莎草类、豆科植物和杂类草)对氮沉降的响应呈现出不同的规律,导致群落物种组成的变化和多样性的降低[24-25]。在高寒草原植物群落主要物种重要值分析中发现,随着施氮量的增加,草地早熟禾始终是高寒草原研究区群落的主要优势种,在NⅤ处理下短芒披碱草的重要值明显增加,其他物种的重要值随施氮处理的不同而不同。这可能是因为草地早熟禾和短芒披碱草是氮素这种限制性资源的有力竞争者,之前的研究发现,不同施氮处理下铁卜加高寒草原的N∶P < 14,氮素是其主要的限制性营养元素[26],施氮量的不断增加使草地早熟禾和短芒披碱草在群落中的丰度增加,最终成为群落的优势种[27]。
目前,关于氮添加对群落物种多样性影响的研究报道很多,大量草地氮添加试验表明,增氮显著降低了群落物种多样性[28-29],但也有一些研究表明,氮素添加对物种多样性无显著影响[11]。本研究发现,NⅠ、NⅡ、NⅢ和NⅣ处理对高寒草原的物种多样性影响不显著,但NⅢ和NⅣ处理下的物种多样性高于CK,NⅤ处理显著降低了Shannon-Wiener多样性指数,Patrick丰富度指数和Pielou均匀度指数虽有所下降但不显著,这表明适量的氮素添加有利于物种多样性的增加,但高倍氮的施加会影响植物的正常生长,降低物种多样性,这一结果与高海燕等[30]的研究结果相似,即在自然降水条件下物种多样性随着施氮量增加,整体呈现先上升后下降的趋势。导致这一研究结果的原因除了氮素作为植物生长的关键因子,适量添加可以增加土壤养分,促进植物生长,但过量则会引起土壤酸化,改变土壤碳氮分配,影响植物的正常生长外,还可能与草地早熟禾等高大禾草在群落中的优势度增加导致的光竞争有关,从而降低了群落内物种多样性[23]。
青藏高原高寒草原的稳定性受到种间竞争、气候变化等因素的影响,本研究采用改进的M.Godron稳定性测定方法计算高寒草原植物群落稳定性,结果表明,NⅢ、NⅣ处理的稳定性距离较大,群落稳定性较低,而NⅤ处理的稳定性距离最小,群落更加趋于稳定,但CK、NⅠ和NⅡ处理的稳定性无明显规律。研究表明,青海湖流域草本植物群落多样性与稳定性呈负相关关系[10],本研究也得出了相似结论,即群落多样性和均匀度越低,群落稳定性越高,这与May通过数学推导认为生态系统的稳定性随物种多样性的增加而下降相一致[11]。根据质量效应,群落稳定性主要受优势种的影响,禾本科作为优势功能群其稳定性大于杂类草的稳定性[31-32],植物群落受到氮沉降的影响时,禾本科作为优势功能群其高度和生物量会有显著的增加,杂类草作为从属功能群其高度和生物量显著低于禾本科植物,将会受到优势功能群光竞争的抑制,优势种的重要值增加,稳定性参考点更加接近(20, 80),导致植物群落整体稳定性增加。因此,利用改进的M. Godron稳定性测定方法得出的稳定性结果受到物种多样性和优势物种重要值占比的影响。另外,稳定性本身具有复杂多层次性的特点,后续研究中应该结合生产力稳定性和土壤肥力等指标综合评估高寒草原在持续氮沉降现状下的群落稳定性。
4 结论
氮添加改变了高寒草原植物群落功能群和物种组成,首先莎草科植物的重要值随着施氮量的增加而增加,当施氮量增加到一定程度后禾本科植物的重要值开始增加,杂类草的重要值明显降低,豆科植物重要值无明显变化规律,但草地早熟禾作为群落中的主要优势种始终未发生改变。氮添加对高寒草原Patrick丰富度无显著影响,但与NⅠ处理相比,NⅤ处理下群落内物种数显著降低,Shannon-Wiener多样性和Pielou均匀度随施氮量增加总体呈现先增加后降低的趋势。通过Pearson相关分析得出不同施氮处理下群落稳定性与Shannon-Wiener多样性和Pielou均匀度呈显著负相关关系。综上所述,经过连续两年施氮处理,本研究发现,8~72 kg·(hm2·a)-1氮倍增试验对高寒草原植物群落组成和物种多样性有一定的促进作用,但72 kg·(hm2·a)-1处理下的施氮量开始对此表现出了一定的抑制作用,进而对群落稳定性产生影响,这一结论可以为氮沉降背景下青藏高原高寒草地的适应性管理提供科学基础,具有重要理论和实践意义。
