氮添加对马尾松人工林凋落物分解的影响
2019-12-10王万才闫文德李忠文梁小翠
王万才,闫文德,4,李忠文,梁小翠
(1.中南林业科技大学,湖南 长沙 410004;2.南方林业生态应用技术国家工程实验室,湖南 长沙 410004;3.城市森林生态湖南省重点实验室,湖南 长沙 410004;4.湖南芦头森林生态系统国家定位观测研究站, 湖南 平江 414000)
大气氮沉降已成为全球变化的一个主要因子[1]。近年来,全球氮沉降呈现迅速增加的趋势,对陆地生态系统结构和功能产生了重要影响[2]。随着工农业的发展,我国氮沉降量从1980年的1.32 g·m-2增长到2010年的2.11 g·m-2[3],目前仍在逐年增加。大气氮沉降通过影响植物生长和物候变化,引起生态系统中环境因子和氮素有效性的改变,进而对凋落物的分解过程产生影响[6-7]。目前的大多数研究主要集中在氮素缺乏的温带地区,缺乏对森林类型比较复杂的热带和亚热带地区大气氮沉降对凋落物分解过程的影响研究[8]。例如,韩雪等[4]研究表明不同形态、不同水平的氮沉降对温带森林凋落物的分解产生了抑制作用,且随氮沉降水平的增加凋落物分解速率降低。李媛媛等[5]指出氮添加对温带森林细根分解具有阶段性影响,分解前期促进细根分解,分解后期抑制分解,这些研究都没有得到一致的结论。因此,进一步加强氮沉降对热带与亚热带森林凋落物分解过程的研究是极为必要的[8]。另外,有报道也指出氮沉降持续增加背景下引起环境因子的改变对土壤有机质和凋落物分解的影响已经成为陆地生态系统与全球变化研究的新生长点和科学研究前沿[10]。以往的研究大多数侧重于凋落叶的分解,缺乏对不同凋落物组分分解过程的研究[9]。然而,植物各组分凋落物的化学成分不同,物理结构也存在差异,造成其木质素和纤维素比例不同,因而具有不同的分解速率[11]。所以,要加强氮沉降对凋落物各组分分解过程的影响研究[9]。
马尾松Pinus massoniana是亚热带典型针叶林树种的代表,也是该地区重要的用材和特种经济树种,具有较强的适应能力和广泛的分布区域,且生长迅速、高产丰产、综合利用程度高。因此,比较研究马尾松林凋落物不同组分(枝、叶)分解速率及其质量、养分含量变化过程对不同氮添加水平的响应,阐明氮添加对亚热带主要森林凋落物分解过程的影响,可为在氮沉降背景下科学合理经营马尾松人工林,深入了解全球植被凋落物分解对氮沉降响应的格局提供科学依据。
1 材料与方法
1.1 研究地概况
研究地设置在湖南省森林植物园(113°02′~ 113°03′ E,28°06′~28°07′ N)内,属于典型的亚热带湿润性季风气候,年均气温约为17.2 ℃,1月份和7月份平均气温分别为4.7 ℃和29.4 ℃。极端最低气温为-11.3 ℃,极端最高气温40.6 ℃,年平均降水量约为1 422 mm,年平均日照时数为1 677.1 h,全年无霜期为270~310 d,海拔为50~100 m,坡度12°~21°,属于典型红壤丘陵区,是由第四纪更新世的冲积性网纹红土和砂砾发育而来的红壤土。林下植被主要有青桐Cordia dichotomaForst.F.、 铁线蕨Adiantum fimbriatumChrist、芒箕Dicranopteris amplaChing et Chiu、狗脊蕨Woodwardia unigemmata、鳞毛蕨Dryopteris podophylla等。
1.2 研究方法
1.2.1 样地设置及氮添加方法
依据当地氮沉降水平和参考同类研究[12],设置对照处理(CK:0 g N·m-2yr-1),低浓度氮处理(LN:5 g N·m-2yr-1),中浓度氮处理(MN: 15 g N·m-2yr-1)和高浓度氮处理(HN:30 g N·m-2yr-1) 4 个氮添加水平。每个氮添加水平分别设置3 块面积均为20 m×20 m 的固定样地,相邻样地之间均设置10 m 宽的缓冲带,一共设置12 块固定样地。以硝酸铵(NH4NO3)作为唯一的氮素添加来源,将NH4NO3充分溶解于20 L 自来水中,使用喷雾器均匀喷洒至各样地内,对照样地仅喷洒20 L 自来水。分别于每年的5月和10月分两次进行等量施氮处理[13]。
1.2.2 凋落物分解方法
凋落物的分解运用尼龙网袋法。选用网眼为1 mm×1 mm,缝成25 cm×25 cm 的尼龙网袋,将各样地收集的凋落物按照落枝和落叶分类,烘干后装入分解袋中(20 g/袋),总共216 个分解袋。将准备好的尼龙网分解袋贴着土层水平放置,每个样地放置18 个,各分解袋之间保持一定的水平距离。放置好分解袋后,开始按照1.2.1 中的方法进行氮素添加处理。在一年的分解实验内,从12月开始第一次取样,每2 个月取样一次,每次每个样地取样3 个,直至试验结束。取回分解袋后,仔细去掉泥沙等杂物,在80 ℃条件下烘干至恒质量,再称其质量。算出凋落物的残留率和失重率,再粉碎凋落物的样品后做养分分析。
1.2.3 凋落叶、凋落枝的分解速率计算
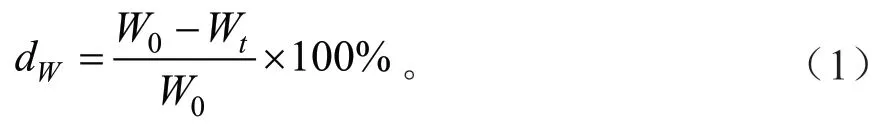
式(1)中:dW为凋落物的失重率(%);Wt为t时段所取样品的质量(g);W0为初始凋落物的质量(g)。
1.2.4 数据测定和处理
分别测定不同氮添加处理下不同分解时间的凋落物养分(C、N、K、Ca、Mg):C 含量用浓硫酸-重铬酸钾法测定;全N 含量用凯氏定氮法测定;全P 含量用钼锑抗比色法测定;全K 含量用火焰光度法测定;Ca、Mg 等元素含量用原子吸收分光光度计测定。
利用Excel 2010 对试验结果进行计算和整理,通过SPSS 20.0(SPSS Inc.,Chicago,IL,USA)进行单因素方差分析(One-Way ANOVA),显著性水平为0.05。
2 结果与分析
2.1 凋落物的初始元素含量
由表1可见,凋落物初始元素含量大小顺序为:C >N >Ca >K >S >Mg >P。其 中,C、N 是凋落物最主要的养分,含量范围分别为446.79~480.22 g·kg-1、4.61~6.66 g·kg-1。凋 落物中C、N 元素含量都呈现出:叶>枝,而Ca 元素含量和C/N 比都呈现出:枝>叶。其余元素的含量都在0.20~2.31 g·kg-1之间变化且同一元素在凋落物不同组分之间的变化不是很明显。

表1 马尾松凋落物各组分中元素的初始含量Table1 The initial content of elements in the litter of Pinus massoniana
2.2 不同氮添加水平下凋落物干质量残留率的变化
在不同浓度的氮添加处理下,马尾松凋落枝分解的干质量残留率变化如图1所示。可以明显的看出,马尾松凋落枝的初期分解速率比较缓慢,分解时间达到10 个月以后马尾松凋落物枝的分解速率明显变快。在分解时间达到12 个月,CK、LN、MN、HN 处理马尾松凋落枝的残留率均在60%以下。根据干质量残留率的变化规律可以发现,分解12 个月后,HN、MN 和LN 处理的凋落枝干质量残留率分别是对照处理(CK)的1.13、1.10和1.07 倍。
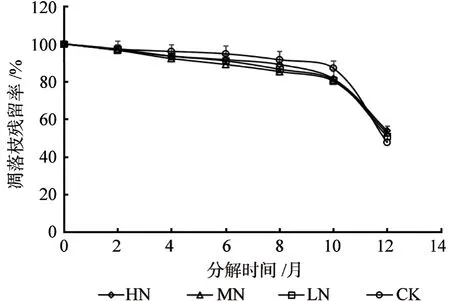
图1 不同处理下马尾松凋落枝分解的残留率变化Fig.1 The residual rate of Pinus massoniana branches litter decomposition under different treatments
在不同浓度的氮添加处理下,马尾松凋落物叶分解的干质量残留率变化如图2所示。可以看出,马尾松凋落叶的干质量残留率在实验周期内的变化都非常缓慢。在分解时间达到12 个月,CK、LN、MN、HN 处理马尾松凋落叶的残留率均在60%以上。分解12 个月后,对照处理(CK)的凋落叶干质量残留率是LN、MN和HN处理的1.02倍。

图2 不同氮浓度处理下马尾松凋落叶的干质量残留率变化Fig.2 The residual rate of Pinus massoniana leaf litter decomposition under different treatments
在整个试验期(12 个月)内,马尾松凋落枝的干质量残留率要比凋落叶的干质量残留率低,说明马尾松凋落枝比凋落叶分解更快。然而,在不同浓度氮添加处理下,马尾松凋落枝和叶的干质量残留率与相应的对照处理相比变化趋势不同。结果表明,氮添加对马尾松凋落枝的分解有一定的抑制作用,相反施加氮素对马尾松凋落物叶的分解有一定的促进作用。
2.3 不同氮添加水平下凋落物分解过程中元素含量的变化
由图3可见,不同氮添加水平下马尾松的凋落叶在分解过程C 含量呈现出释放-富集-释放的趋势。在整个实验周期内,随分解时间的变化,不同氮添加水平下凋落叶C 含量的变化幅度都较小。马尾松凋落枝分解过程中C 元素含量的变化在不同氮添加水平下有所不同。在HN 处理下,C的含量在前6 个月的分解过程中下降最为明显,并且在分解时间为6 个月时达到整个分解过程碳含量的最小值;而在LN 与MN 处理下,在分解时间达到8 个月时凋落枝中C 元素含量才达到最小值。在分解时间达到10 个月时,凋落枝中C 元素含量同时达到分解过程最大值。

图3 马尾松凋落物枝和叶分解中C、N 元素的含量变化Fig.3 Changes of the carbon and nitrogen contents in the branch and leaf litter of Pinus massoniana
马尾松凋落叶和枝分解过程中N 元素的含量也呈现出释放-富集-释放的趋势,但其下降的趋势都不是很明显。与LN 相比,HN 和MN 两种不同氮添加水平下马尾松凋落叶分解过程中N 元素含量的变化幅度较大。在整个试验过程中,马尾松凋落枝中的N 元素含量在不同氮添加水平下始终表现为HN >LN >MN,这与马尾松凋落枝中N 元素的初始含量大小顺序相一致。马尾松凋落枝中N 元素含量在前4 个月的分解过程中快速下降,并且在分解时间为4 个月时达到整个分解过程N 含量的最小值。
由图4可见,马尾松凋落叶在分解过程中K、Mg 元素的含量呈现出释放-富集-释放的趋势,而Ca 元素含量则呈现富集-释放的趋势。在不同氮添加水平下,K 元素含量在分解过程的前6 个月内都表现出一致的下降趋势。LN 处理下,K 元素含量分解时间8 个月后的变化不同于其他处理而呈现出释放的趋势。Ca 元素含量在分解过程的前10 个月内都表现出一致的上升趋势,并且在分解时间为10 个月时含量达到整个过程的最大值,之后随着分解时间的变化含量逐渐降低。不同处理之间的变化基本一致。Mg 元素含量则在分解过程的前8 个月内都呈现下降趋势。HN 处理下,在分解过程的前8 个月内其变化不同于其处理。
马尾松凋落枝分解过程中K、Ca、Mg 元素含量呈现出释放-富集-释放的趋势。在不同氮添加水平下,前8 个月的分解时间内K 元素含量变化一致,后期LN 和HN 处理与MN 有所不同。在不同添加水平下,Ca 元素含量在分解过程变化比较一致。与其他处理相比,在HN 处理下,Ca 元素含量的变化幅度较大。在分解时间为4~12月期间,HN 处理下Mg 元素含量的变化趋势与其他处理呈现出相反的趋势。
3 结论与讨论

图4 马尾松凋落物枝和叶分解中K、Ca、Mg 元素的含量变化Fig.4 Changes of the K,Ca,Mg contents in the branch and leaf litter of Pinus massoniana
森林生态系统中的凋落物分解过程是其物质循环和能量流动的重要环节[14]。研究表明,植物所吸收的90%以上的N、P 以及60%以上的矿质元素都来自于植被归还给土壤的养分再循环[15]。因此,森林凋落物分解的快慢对系统内养分循环具有重要影响。在本研究一年的分解时间内,马尾松凋落物的干质量损失率在29%~52%之间。这与Pausas 等人[16]的研究结果陆地森林生态系统中凋落物第一年的干质量损失率达到30%~70%和亚热带森林凋落物40%~70%的年分解速率[17-18]相一致。已有的研究表明,凋落枝和叶约占森林生态系统凋落物年归还总量的26%和70%[19],在凋落物各组分中占绝对优势[20-23]。所以,森林凋落物的分解快慢主要取决于凋落枝和叶的分解过程。已有研究大多数关注凋落叶、细根、粗母质残体(倒木等)的分解过程,较少涉及凋落枝[24-25]。本实验中,马尾松凋落物枝和叶表现出不同的分解过程。经过一年的分解,马尾松凋落枝比凋落叶分解更快,但是前10 个月枝和叶的分解速率基本一致。郭彩虹等[9]对岷江冷杉凋落枝分解实验表明,枝条冬季总质量损失比例明显高于生长季节。这主要是由于冬季冻融循环特征及耐寒生物种群活动明显促进了寒冷季节凋落枝的分解过程[26-27]。另一方面,在生长季节,强光照射到林地内使得凋落物平均温度升高,促进了凋落物水分的蒸发,引起比较强烈的干湿交替过程,这可能不利于土壤无脊椎动物和微生物对凋落物的分解[28],导致此时凋落物分解过程比较缓慢。这也是本研究中出现凋落物枝和叶在前10 个月内分解速率基本一致,而凋落枝在10 个月后分解明显加快的主要原因。同时有研究表明,在凋落物后期的分解速率与凋落物初始N 浓度呈负相关[29],即在凋落物分解后期初始N 浓度高的凋落物比低的凋落物分解慢[30]。因此,在整个实验过程中表现出马尾松凋落枝比凋落叶分解更快。
从已有的研究结果来看,氮沉降对凋落物分解的影响并没有一致的结论,包括促进凋落物分解[31-32]、延缓凋落物分解[33]以及对凋落物分解无影响[34]三种情况。我们的研究结果显示:分解12个月后,CK、HN、MN、LN 处理马尾松凋落枝的残留率均在60%以下,并且HN、MN 和LN 处理的干质量残留率分别是对照处理(CK)的1.13、1.10 和1.07 倍;而CK、HN、MN、LN 处理马尾松凋落叶的残留率均在60%以上,并且对照处理(CK)的凋落叶干质量残留率是LN、MN 和HN处理的1.02 倍。可以看出,氮添加对马尾松凋落枝的分解有一定的抑制作用,随着氮浓度的升高,抑制作用也随之增强;相反施加氮素对马尾松凋落物叶的分解有一定的促进作用,但不同氮浓度水平之间的差异不显著。通常来说,外源氮的输入会促进高质量(高N,低C/N 等)的凋落物的分解[35],而抑制质量较低的凋落物的分解[36-38]。本研究中与凋落枝相比,凋落叶有高N 浓度和低C/N 比。因此,氮添加一定程度上促进了马尾松凋落叶的分解。另外,N 沉降可能通过改变生态系统中营养平衡、抑制相关动物和微生物的活动、阻碍调控凋落物分解的胞外酶活性以及增加木质素和纤维素含量来抑制凋落物的分解[39]。Berg 和Matzner 研究表明,随着氮浓度的增加苏格兰松Pinus sylvestris和挪威云杉Picea abies的凋落物中木质素含量呈现显著增加趋势[40]。所以,氮添加抑制了凋落枝的分解过程。
随着凋落物的分解,贮存在凋落物中的养分会在森林生态系统中不断流动,从而起到维持系统养分平衡和土壤肥力等重要作用[41],并且凋落物分解时养分的流动过程对森林土壤有机碳含量变化以及系统内的养分循环过程也发挥了非常重要的作用[42]。本研究中,凋落物初始元素含量与以往的大多数研究相一致[43-45],都表现为:C > N >Ca >K >S >Mg >P。经过一年的分解,在LN、MN、HN 处理下,马尾松凋落叶中C 含量分别下降20.0%、24.1%、14.8%,凋落枝中的C 含量分别下降了9.6%、13.6%、13.8%;凋落叶N 含量分别下降了21.5%、25.8%、34.7%;凋落枝N 含量分别下降了36.9%、7.4%、54.4%。已有研究表明,模拟氮沉降处理促进起始碳含量低的凋落物C 释放,起始氮含量高的凋落物N 释放[46],但这种促进作用也受到气候、树种、立地条件以及凋落物种类等诸多因素的影响[47]。因此,在本实验中氮添加促进了马尾松凋落枝中的C 元素和凋落叶中N 元素的释放,并且随着氮浓度的增加促进作用有所增强。由于金属离子易发生淋溶,K、Mg 浓度在分解前期呈递减趋势[48],但在后期呈富集趋势。同时,K 和Mg 的流动性较大,K 以可溶性盐基形式存在于植物体内,易于渗滤释放,Ca 是结构性元素,流动性差,不易被再吸收利用[49]。所以,本研究中马尾松凋落枝和叶分解前期过程中K、Mg 元素含量逐渐下降,而Ca 元素含量有所升高。总之,凋落物分解过程的养分动态是由物种组成、凋落物类型以及温度、湿度、降水、土壤、微生物等内外因素综合作用的结果[50-52]。而罗佳等[53]研究也表明凋落物叶和枝中养分含量的释放尤其受土壤总氮和速效磷的调节。因此,要在今后的研究中加强多种因素对森林凋落物分解过程和养分释放动态的影响研究。同时,也要加强单一和混合凋落物分解过程对全球变化的响应研究,为全面地认识全球变化对森林凋落物分解过程的影响提供参考。
