细菌降解低分子量多环芳烃的研究进展
2019-12-09周子康崔洁许平唐鸿志
周子康,崔洁,许平,唐鸿志
综 述
唐鸿志 上海交通大学生命科学技术学院/微生物代谢国家重点实验室教授、博士生导师 (2014年破格晋升)。2012年12月–2013年12月美国麻省理工学院访问学者。研究方向为环境微生物学。目前担任中国微生物学会-环境微生物专业委员会委员兼秘书。中国生物工程学会-合成生物学专业委员会 (筹) 委员。2016年获得教育部自然科学一等奖 (排名第二),2013年获得“明治乳业生命科学奖”。曾获得国家自然科学基金-优秀青年基金、上海市曙光学者、上海市青年科技启明星、上海市教委“晨光”计划、上海交通大学“晨星计划”、首届“仲英青年学者”,已于、、、、、、、、等期刊发表SCI论文80篇(第一/通讯作者45篇)。

细菌降解低分子量多环芳烃的研究进展
周子康1,2,崔洁1,2,许平1,2,唐鸿志1,2
1 上海交通大学 生命科学技术学院 微生物代谢国家重点实验室,上海 200240 2 上海交通大学 代谢与发育科学国际联合实验室,上海 200240
具有“三致”效应的多环芳烃污染造成了巨大的环境危害,威胁人类健康和生存。目前能够降解低分子量多环芳烃的细菌已有广泛的研究。细菌通过多层次的调控分析和适应性进化提高它们的降解能力。本文基于国内外文献调研,简要总结了生物修复在低分子量多环芳烃降解领域的研究进展。拟通过多层次的调控分析和适应性进化来产生多种分解代谢途径,为生物降解能力强化的未来降解技术提供支撑。
多环芳烃,低分子量多环芳烃,生物修复,芴
多环芳烃PAHs (Polycyclic aromatic hydrocarbons)是指由两个或两个以上苯环以线状、角状或簇状排列的碳氢化合物,大多来源于有机物的不完全燃烧或高温裂解[1]。PAHs广泛存在于石油、煤炭中,具有潜在的致畸性、致癌性和基因毒性,且毒性随着苯环的增加而增加[2-3],对环境、生物体以及人类都有极大的危害。由于多环芳烃类化合物具有极低的水溶性,在环境中很难消除,因此,PAHs被美国环保局和欧共体同时确定为优先控制的污染物,并选定其中的16种多环芳烃类化合物作为环境污染的监测对象[4](表1)。
微生物扮演着生态系统中最重要的分解者的角色,可产生多种多样的复合酶系,催化复杂化合物的降解,其代谢多环芳烃的能力较强,代谢速率较高。因其能将污染物进行完全矿化,具有相比物化方法更低的处理成本、更高的安全性、更好的环境包容性等潜在的优点,微生物降解成为了有效解决PAHs污染物危害的最佳选择[5-7]。
研究发现,细菌可以通过代谢或共代谢的方式以PAHs为底物来满足自身生长需求[8]。其中的低分子量多环芳烃 (Low molecular weight PAHs, LMW PAHs,2个或3个苯环组成) 相对于高分子量多环芳烃 (High molecular weight, HMW PAHs,4个及以上苯环组成) 较易挥发,水溶性较高,因此它们更容易生物降解[9]。在LMW PAHs中,萘、蒽和菲在整个环境中广泛分布,并且被认为是原型PAHs。它们的核心结构骨架存在于许多致癌PAHs中,因此常用作检测PAHs污染的标志性化合物。它们还被用作模式PAHs,以确定高环芳烃的生物利用效率、生物降解潜力和环境中细菌降解速率等的影响因素[10]。同样被用作模式PAHs的还有三环芳烃——芴,它是具有致癌性的化石燃料的主要成分。与咔唑、二苯并噻吩、二苯并呋喃、二苯并二噁英等具有一定的结构关系,且同时具有芳香族和脂环族的化学结构[11-13]。本文基于国内外多环芳烃研究进展,并结合笔者实验室多环芳烃降解的研究近况,总结了LMW PAHs,包括萘、蒽、菲、芴的多种细菌代谢途径。
1 萘、菲、蒽的细菌分解代谢
萘 (Naphthalene) 是广泛存在于环境中的双环芳烃,早在1964年,Davies和Evans就报道了第一株能降解土壤中萘的假单胞菌[14]。研究发现萘的主要代谢途径如图1A所示。首先,在萘双加氧酶(NDO) 的催化作用下,萘的苯环的1,2位引入氧分子,生成顺式-(1R, 2S)-二羟基-1,2-二氢化萘 (顺式-萘二氢二醇) (图1A) 化合物A-Ⅱ)。该双加氧酶系统由3种成分组成,即铁氧还蛋白还原酶、铁氧还蛋白以及由两个不同的亚基α和β组成的铁硫蛋白 (ISP)。电子传递是通过在铁氧还蛋白还原酶中从NAD(P)H到FAD的单个双电子转移引发的,其产生完全还原形式的FAD。还原的FAD为铁氧还蛋白的铁硫簇 (2Fe-2S) 分别提供一个电子,这些电子最终转移到ISP并用于其活性位点,以促进氧分子的加成(图2)。NDO的底物特异性已经有了很详细的研究[15],其组分的三维结构也已被鉴定[16],Parales等[17]发现NDO催化结构域中的Asp-205对其活性至关重要,晶体结构显示NDO的另一种底物吲哚会在单核铁原子的附近被结合,这表明此残基是氧的活化位点[18]。后续研究通过定点诱变以测定几个活性位点残基对于催化活性的关键性,发现Phe-352的取代会导致顺式萘二氢二醇的形成,同时伴随着立体化学结构及联苯和菲的氧化位点的改变[19],此结果为NDO在其加氧酶组分中与底物的结合提供了良好的结构模型。

表1 美国环保局认定的16种优先PAHs化合物
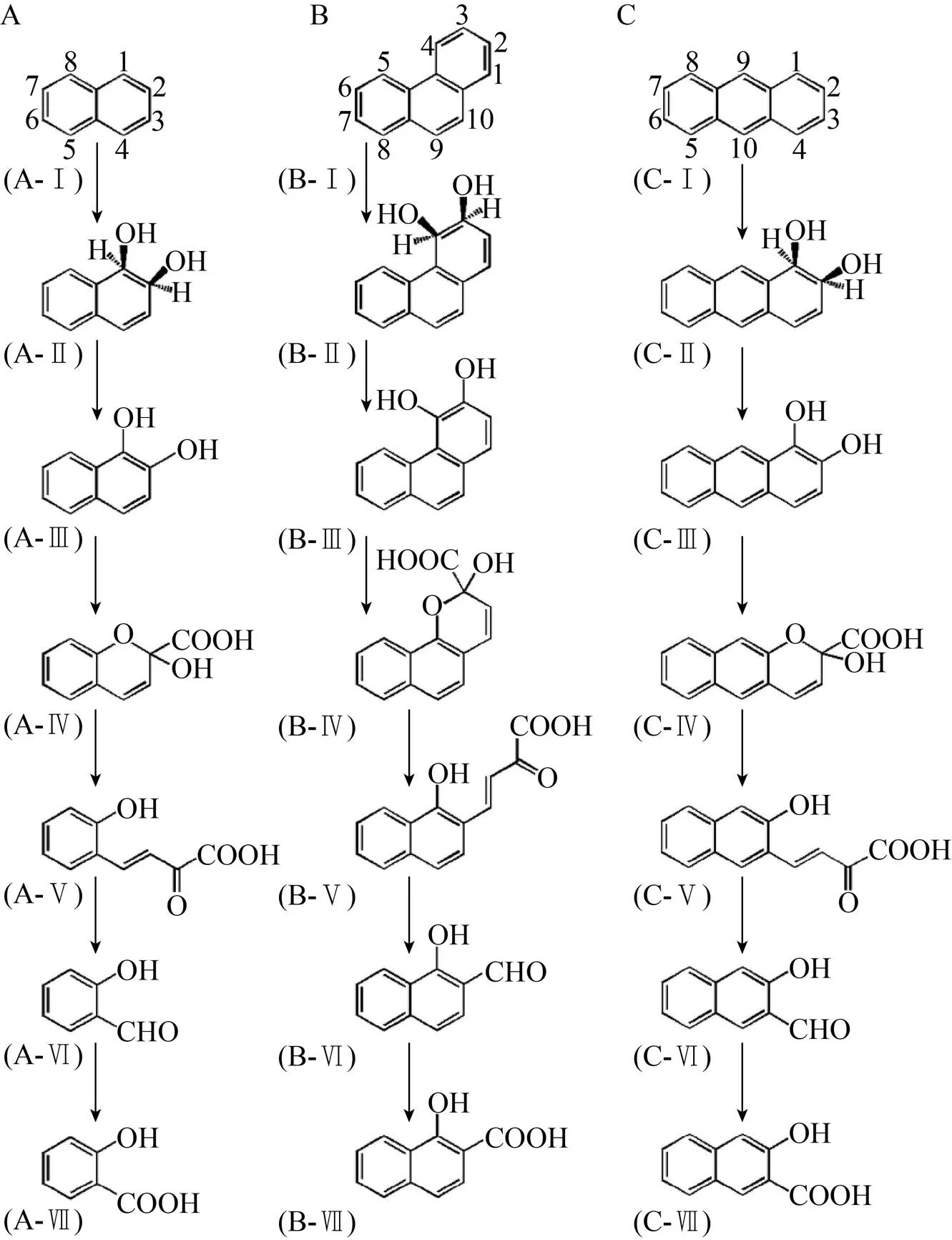
图1 萘 (A)、菲 (B)、蒽 (C)的上游分解代谢途径

图2 萘双加氧酶将萘初步氧化成顺式-1,2-二羟基-1,2-二氢化萘
第二步反应是在顺式-萘二氢二醇脱氢酶的催化作用下,顺式-萘二氢二醇脱氢形成1,2-二羟基萘 (图1A,化合物A-Ⅲ)。1,2-二羟基萘通过1,2-二羟基萘双加氧酶进行复合,得到的环裂解产物自发地形成2-羟基-2-氢-色烯-2-羧酸 (图1A),化合物A-Ⅳ)。通过异构酶和水杨酸酶-醛缩酶的酶促反应生成水杨醛 (图1A,化合物A-Ⅵ),然后在水杨醛脱氢酶的催化作用下转化为水杨酸 (图1A,化合物A-Ⅶ)。水杨酸通过邻苯二酚 (图3,化合物A-Ⅱ) 或2,5-二羟基苯甲酸 (图3,化合物B-Ⅱ)进一步代谢为TCA循环中间体。
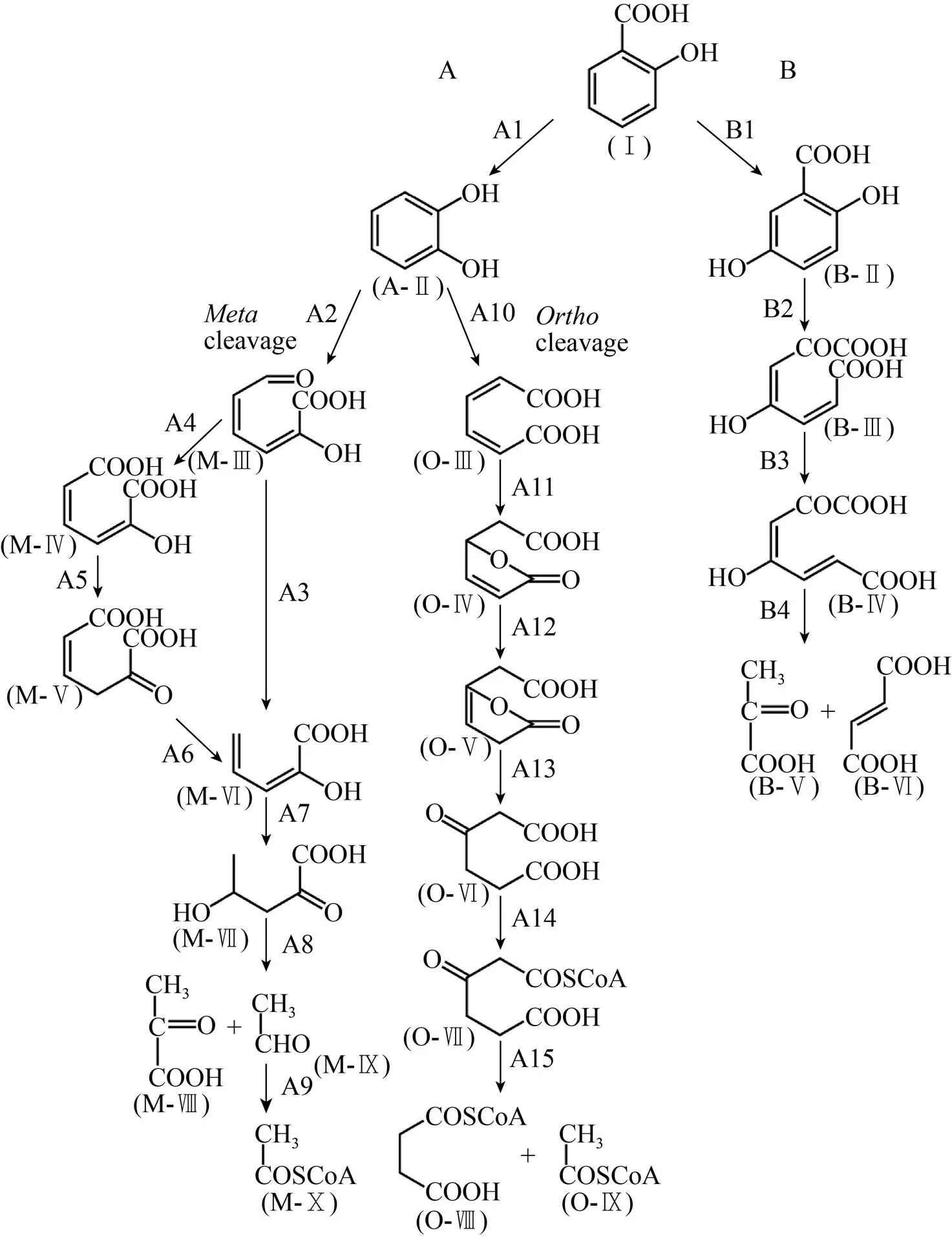
图3 通过邻苯二酚 (A) 和2,5-二羟基苯甲酸 (B)的萘的下游代谢途径
菲 (Phenanthrene) 由3个苯环组成,常用作研究致癌PAHs代谢的模式化合物。在过去的60年中,已经报道了许多革兰氏阴性和阳性细菌降解菲的研究[20-23],菲的主要代谢途径如图1B所示。参与萘转化为水杨酸的酶可以通过类似的分解代谢反应将菲降解为1-羟基-2-萘甲酸 (图1A,化合物 B-Ⅶ),其主要通过两个途径进行进一步代谢。一种分解代谢途径涉及1-羟基-2-萘甲酸的羟基化,形成1,2-二羟基萘 (图4A,化合物A-Ⅱ),然后进入萘降解途径。在另一种代谢途径中,1-羟基-2-萘甲酸的苯环被1-羟基-2-萘甲酸双加氧酶直接裂解 (图4B)。后通过酶促反应生成邻苯二甲酸酯 (图4B,化合物B-Ⅳ),其通过原儿茶酸酯 (图4B,化合物 B-Ⅴ)进一步代谢为TCA循环中间体。
相似地,蒽 (Anthracene) 的主要分解代谢途径的第一步是通过在1,2位的初始氧化作用,蒽转化为1,2-二羟基蒽 (图1C,化合物C-Ⅲ)。然后通过1,2-二羟基蒽的间位裂解途径,生成2-羟基-3-萘甲酸 (图1C,化合物C-Ⅶ),2-羟基-3-萘甲酸酯紧接着转化为2,3-二羟基萘,其进一步代谢为水杨酸盐和邻苯二酚。

图4 通过1,2-二羟基萘 (A) 和邻苯二甲酸酯 (B)的菲的下游代谢途径
截至目前,已经研究报道的可降解萘、蒽、菲的细菌超过200种,早在60年代就有研究,之后陆续在假单胞菌属中发现很多代谢萘、蒽、菲的基因,例如铜绿假单胞菌中的基因簇[28],恶臭假单胞菌中的基因簇[24-26],sp中的基因簇[31]等,为萘、蒽、菲及更复杂的多环芳烃的降解研究提供了重要元器件和实践基础。
在基因簇中,是编码萘、蒽、菲初始氧化作用的双加氧酶的操纵子,分别编码代谢途径中的脱氢酶,二羟基萘、蒽、菲双加氧酶,异构酶,水合酶醛缩酶,以及水杨醛脱氢酶,最终代谢为水杨酸。在基因簇中,是编码萘、蒽、菲初始氧化作用的双加氧酶的操纵子,分别编码途径中剩余的酶。在基因簇中,是编码萘、蒽、菲初始氧化作用的双加氧酶的操纵子,分别编码途径中剩余的酶。具体的基因编码情况如表2所示。
表3总结了包括以上3个降解菌株的更多降解PAHs菌株的生长底物特性以及参与其上游分解代谢途径的酶对萘、菲和蒽的转化能力。对于列出的大多数细菌,都可以利用萘和菲作为底物生长,然而,通过邻苯二甲酸酯途径的菲降解细菌 (例如,菌株AFK2和KP7) 不能以萘为唯一碳源进行生长。
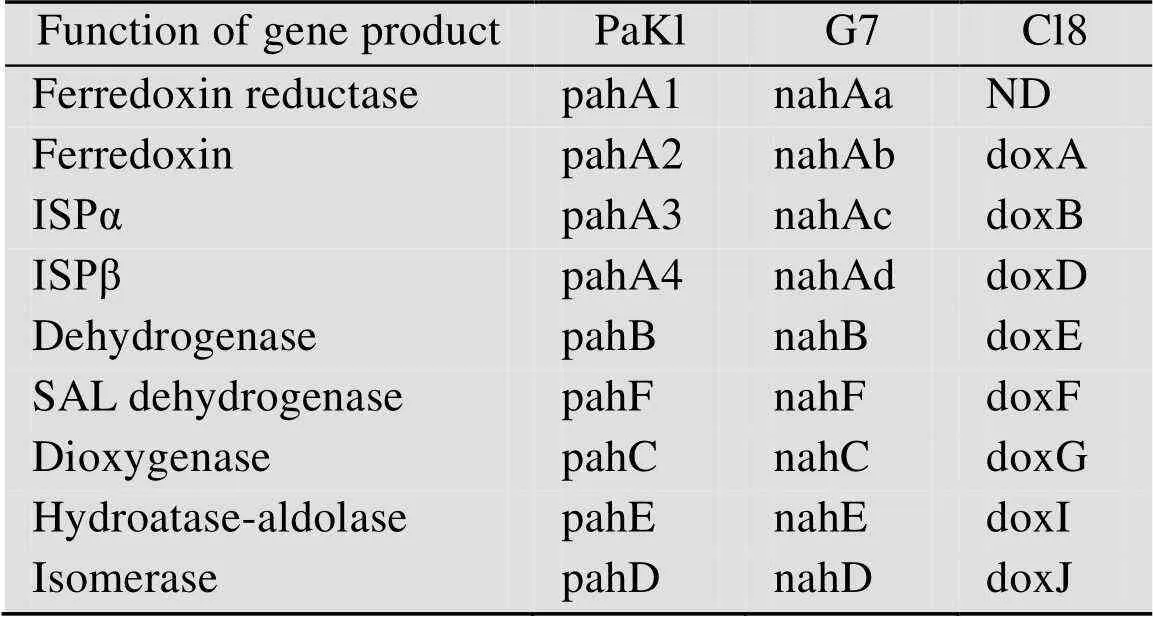
表2 假单胞菌中萘、蒽、菲上游降解基因簇
ND: indicates that the gene encoding the enzyme has not been detected.

表3 PAHs降解细菌的基因定位、生长底物特异性和酶底物特异性
a. Listed PAH-degrading bacteria; b. “n” means “not yet reported”; c. Nap, Phe and Ant represent naphthalene, phenanthrene and anthracene, respectively; “+” and “-” mean “growth” and “non-growth”, respectively; d. “+” means that a clone carrying a metabolic pathway enzyme-encoding gene can convert naphthalene to salicylic acid, convert phenanthrene to 1-hydroxy-2-naphthoic acid, or convert hydrazine to 2-hydroxy-3-naphthoic acid. “(+)” means that at least the initial double oxidative decomposition of each PAH compound was observed.
2 芴的降解
芴 (Fluorene) 是美国环保局确定的16种优先处理的污染物之一,存在于汽车废气、玉米须以及煤焦油的高沸点组分中,有类似于萘的特征性芳香气味。2017年10月27日世界卫生组织国际癌症研究机构公布的致癌物清单中,芴被列入3类致癌物。经过多年的研究,已经发现了不少可以利用芴作为唯一底物的菌株[10,42-45],芴的3种主要代谢途径如图6所示。其中两种途径分别是通过1,2位 (图5A)和3,4-位 (图5B) 的双氧化作用开始的,生成相应的顺式-二氢二醇 (图4,化合物A-Ⅱ,化合物B-Ⅱ),紧接着发生脱氢反应,然后进行间位裂解。在醛缩酶反应和环状产物脱羧反应后,得到的茚酮是Baeyer-Villiger反应的底物,经反应后分别得到芳香内酯3-异色酮和3,4-二氢香豆素 (化合物A-Ⅵ和B-Ⅵ)。3,4-二氢香豆素可通过酶促水解产生3-(2-羟基苯基) 丙酸 (化合物B-Ⅶ),其可进一步代谢为水杨酸。第三种途径 (图5C) 通过在C-9位置的单氧化作用引发,得到9-芴醇,然后通过脱氢反应生成9-芴酮。9-芴酮在角二氧化作用下生成1,1a-二羟基-1-氢-9-芴酮 (化合物C-Ⅳ),接下来发生五元环的裂解生成2,3-二羟基联苯的2′-羧基衍生物,其通过类似于联苯降解的反应进行分解代谢,导致邻苯二甲酸的形成,然后通过3,4-二羟基苯甲酸的产生代谢邻苯二甲酸。

图5 芴的分解代谢途径
尽管已经分离和表征了许多降解芴的细菌,但是关于芴的分解代谢中涉及的特定酶,特别是编码这些酶的基因,还需要更深入的研究。本文以地杆菌sp为例,对芴降解的基因簇加以介绍。该基因簇由、、、和组成,其中编码芳环双加氧酶,均编码短链的还原脱氢酶,编码二羟基双加氧酶,编码meta-cleavage水解酶,在以上酶系的作用下将芴分解为邻苯二甲酸 (图6)。
近期,笔者实验室同仁从上海老港垃圾填埋场分离到一株以芴为唯一碳源的SMT-1菌株[46],经16S rDNA基因序列分析鉴定为假单胞菌。该菌株在含0.4 mm芴的无机盐培养基中,以30 ℃、pH 7.0和200 r/min条件下生长和降解活性最高。此外,菌株SMT-1在连续6 d内降解不同多环芳烃的比较结果,芴为85%,菲为48.4%,二苯并呋喃 (DBF)为47.5%,荧蒽为29.1%。这些降解特性表明,菌株SMT-1在芴降解中具有良好的降解效果,并有一定的底物适应范围。基因组信息的初步分析揭示了该菌株中可能存在参与芳香化合物代谢的重要基因。

图6 芴的分解代谢基因簇
二苯并呋喃的降解也是环境治理领域的热点问题之一。本实验室Fawad Ali等[47]分离鉴定了一株具有高DBF降解活性的铜绿假单胞菌FA-HZ1。该菌株降解中间产物的鉴定表明,其通过利用横向脱氧和间裂途径降解DBF (图7)。一种新型双加氧酶——HZ6359可将DBF转化为1,2-二羟基-1,2-二氢二苯并呋喃。铜绿假单胞菌对DBF的降解作用在以往的研究中尚未见报道,该研究首次明确了铜绿假单胞菌对DBF的降解能力。此外,菌株FA-HZ1还可以利用DBF以外的其他碳源,其针对多底物的广泛降解活性对环境生物修复具有重要意义。
与降解途径相比,低分子量PAHs的分解代谢途径的调控机制的研究也至关重要。研究者就调节过程中的分子伴侣、触发途径表达的信号以及激活和抑制表达的确切机制等进行了深入的研究,很快发现了大量多样的调节系统用于介导分解代谢途径的表达[48]。例如LysR型转录调节因子家族 (LTTR)[49-50],该家族包含不同菌属的100多个成员。其中CatR控制恶臭假单胞菌中邻苯二酚代谢的表达[51],TfdR (和/或其相同的双TfdS) 也调节编码氯代邻苯二酚代谢的操纵子 (即基因)。NahR蛋白是萘降解调控中的主要调节因子,通过控制萘对水杨酸和丙酮酸代谢所需的操纵子和操纵子的表达起作用,编码水杨酸酶,两个操纵子都存在于质粒NAH7上[52-53]。经过20多年的研究,我们理解了不同调控蛋白在介导转录过程中的行为,但对于调节蛋白和效应化合物之间的相互作用、结合位点、信号传导等方面的内容还知之甚少,仍需要进一步探索。
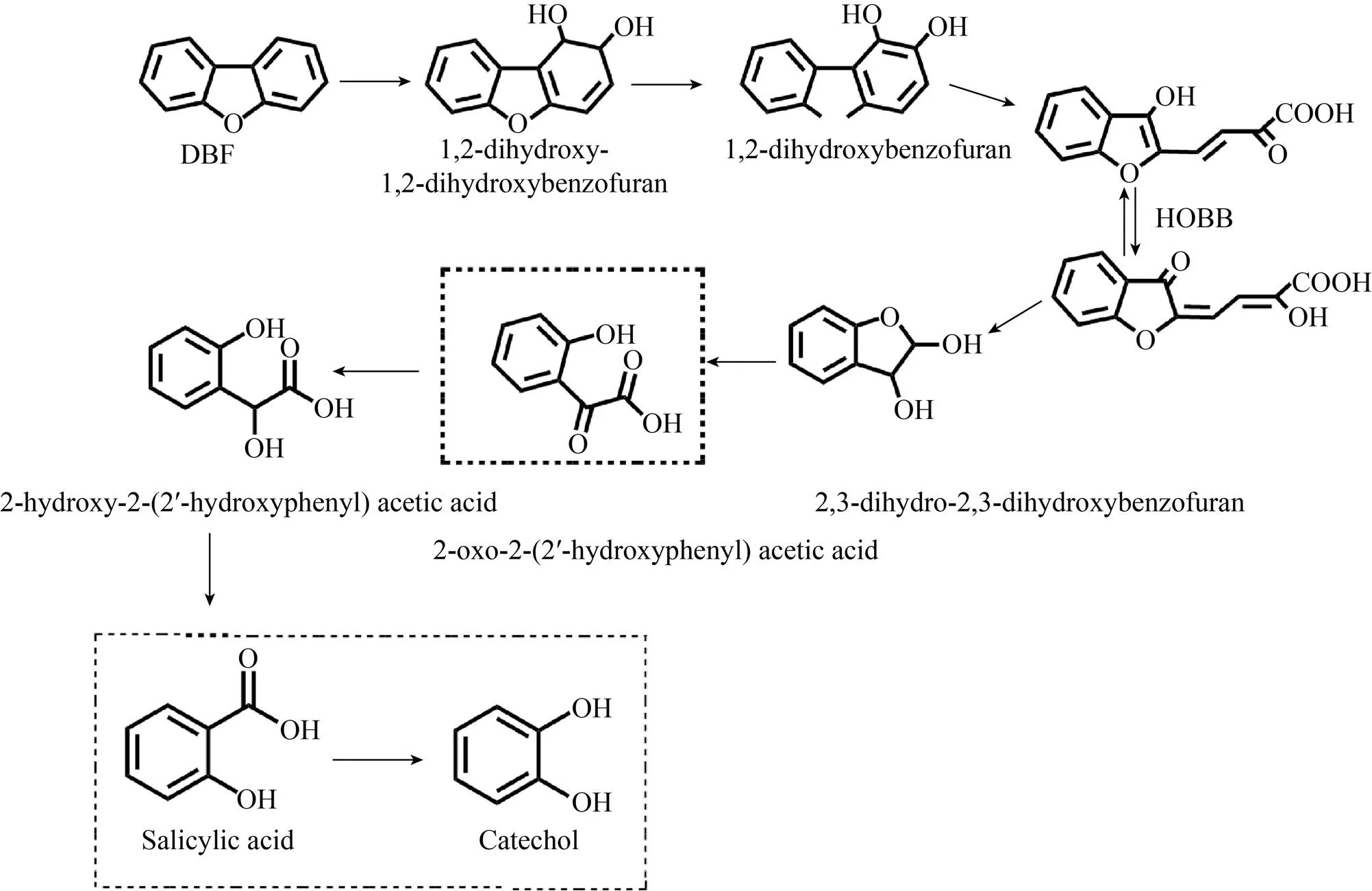
图7 推测出的菌株FA-HZ1代谢DBF的降解途径
3 总结与展望
为了探求更高效的PAHs降解途径,减轻其对环境的危害,生态友好地进行生物修复,需要在分子水平更深入地了解细菌降解代谢PAHs的全过程。经过几十年的深入研究,已报道的可降解二环或三环组成的PAHs的分解代谢基因的数量已经大大增加。然而,在LMW PAHs中,芴和氘代苊等的降解涉及的分解代谢基因还需要更积极地探索,可降解由4个及以上苯环组成的PAHs基因的获取还很有限,由于PAHs的毒性和致癌性,更突显了这些研究的急迫性和重要性。同时,低分子量PAHs关键代谢酶的研究可以为实现改造人工酶进而实现高分子量PAHs降解提供理论基础。因此,不仅要深入了解这些基因和关键酶的功能,还要对PAHs的代谢或共代谢途径有全面的认识,基于此可构建用于监测污染环境中降解物的基因探针,进一步促进生物修复的发展。此外,考虑到环境中污染物的复杂程度,我们需要进一步研究除萘、菲、蒽、芴以外的PAHs的降解。通常认为,高环PAHs开环降解过程会进入低环途径,因此,低分子量PAHs的降解途径对最终修复环境中所有PAHs的污染至关重要。最近,基于PCR的方法,已经报道了许多新的可部分分解代谢PAHs的基因,这些结果表明环境中PAHs降解细菌的多样性,并且可能仍有许多未鉴定出的PAHs降解细菌,包括不可培养的细菌。积累这些信息对于监测PAHs污染土壤中不同PAHs分解代谢基因型的有效性是必要的,并有助于PAHs的有效生物修复。这是我们相关研究的最终目标,相信这一目标将在广大科研工作者的共同努力下实现。
[1] Dat ND, Chang MB. Review on characteristics of PAHs in atmosphere, anthropogenic sources and control technologies. Sci Total Environ, 2017, 609: 682–693.
[2] Wang CY, Wang YD, Herath HMSK. Polycyclic aromatic hydrocarbons (PAHs) in biochar - their formation, occurrence and analysis: a review. Organ Geochem, 2017, 114: 1–11.
[3] Kuppusamy S, Thavamani P, Venkateswarlu K, et al. Remediation approaches for polycyclic aromatic hydrocarbons (PAHs) contaminated soils: Technological constraints, emerging trends and future directions. Chemosphere, 2017, 168: 944–968.
[4] Keith L, Telliard W. ES&T special report: priority pollutants: I-a perspective view. Environ Sci Technol, 1979, 13(4): 416–423.
[5] Cerniglia CE. Biodegradation of polycyclic aromatic hydrocarbons. Biodegradation, 1992, 3(2/3): 351–368.
[6] Shuttleworth KL, Cerniglia CE. Environmental aspects of PAH biodegradation. Appl Environ Microbiol, 1995, 54(1/3): 291–302.
[7] Gibson DT, Parales RE. Aromatic hydrocarbon dioxygenases in environmental biotechnology. Curr Opin Biotechnol, 2000, 11(3): 236–243.
[8] Kanaly RA, Harayama S. Biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by bacteria. J Bacteriol, 2000, 182(8): 2059–2067.
[9] Pannu JK, Singh A, Ward OP. Influence of peanut oil on microbial degradation of polycyclic aromatic hydrocarbons. Can J Microbiol, 2003, 49(8): 508–513.
[10] Moody JD, Freeman JP, Doerge DR, et al. Degradation of phenanthrene and anthracene by cell suspensions ofsp. strain PYR-1. Appl Environ Microbiol, 2000, 67(4): 1476–1483.
[11] Grifoll M, Casellas M, Bayona JM, et al. Isolation and characterization of a fluorine-degrading bacterium: identification of ring oxidation and ring fission products. Appl Environ Microbiol, 1992, 58(9): 2910–2917.
[12] Grifoll M, Selifonov SA, Chapman PJ. Evidence for a novel pathway in the degradation of fluorene bysp. strain F274. Appl Environ Microbiol, 1994, 60(7): 2438–2449.
[13] Grifoll M, Selifonov SA, Gatlin CV, et al. Actions of a versatile fluorene-degrading bacterial isolate on polycyclic aromatic compounds. Appl Environ Microbiol, 1995, 61(10): 3711–3723.
[14] Davies JI, Evans WC. Oxidative metabolism of naphthalene by soil: the ring-fission mechanism. Biochem J, 1964, 91(2): 251–261.
[15] Resnick SM, Lee K, Gibson GT. Diverse reactions catalyzed by naphthalene dioxygenase fromsp. strain NCIB 9816. J Ind Microbiol, 1996, 17(5/6): 438–457.
[16] Kauppi B, Lee K, Carredano E, et al. Structure of an aromatic-ring-hydroxylating dioxygenase-naphthalene 1, 2-dioxygenase. Structure, 1998, 6(5): 571–586.
[17] Parales RE, Parales JV, Gibson DT. Aspartate 205 in the catalytic domain of naphthalene dioxygenase is essential for activity. J Bacteriol, 1999, 181(6): 1831–1837.
[18] Carredano E, Karlsson A, Kauppi B, et al. Substrate binding site of naphthalene 1,2-dioxygenase: functional implications of indole binding. J Mol Biol, 2000, 296(2): 701–712.
[19] Parales RE, Lee K, Resnick SM, et al. Substrate specificity of naphthalene dioxygenase: effect of specific amino acids at the active site of the enzyme. J Bacteriol, 2000, 182(6): 1641–1649.
[20] Blumer M. Polycyclic aromatic compounds in nature. Sci Am, 1976, 234(3): 34–45.
[21] IARC (International Agency for Research on Cancer). IARC monographs on the evaluation of carcinogenic risks to humans//Polynuclear Aromatic Compounds. Part 1. Chemical, Environmental and Experimental Data. Lyon, France: World Health Organization, 1983, 419–430.
[22] Pelkonen O, Nebert DW. Metabolism of polycyclic aromatic hydrocarbons: etiologic role in carcinogenesis. Pharmacol Rev, 1982, 34(2): 189–222.
[23] Gibson DT, Subramanian V. Microbial degradation of aromatic hydrocarbons//Gibson DT, Ed. Microbial Degradation of Organic Compounds. New York: Dekker, 1984: 181–252.
[24] Menn FM, Applegate BM, Sayler GS. NAH plasmid-mediated catabolism of anthracene and phenanthrene to naphthoic acid. Appl Environ Microbiol, 1993, 59(6): 1938–1942.
[25] Yen KM, Serdar CM. Genetics of naphthalene catabolism in. Crit Rev Microbiol, 1988, 15(3): 247–268.
[26] Simon MJ, Osslund TD, Saunders R, et al. Sequences of genes encoding naphthalene dioxygenase instrains G7 and NCIB 9816-4. Gene, 1993, 127(1): 31–37.
[27] Yang Y, Chen RF, Shiaris MP. Metabolism of naphthalene, fluorene, and phenanthrene: preliminary characterization of a cloned gene cluster fromNCIB 9816. J Bacteriol, 1994, 176(8): 2158–2164.
[28] Takizawa N, Iida T, Sawada T, et al. Nucleotide sequences and characterization of genes encoding naphthalene upper pathway ofPaK1 andOUS82. J Bacteriol Bioeng, 1999, 87(6): 723–731.
[29] Kiyohara H, Torigoe S, Kaida N, et al. Cloning and characterization of a chromosomal gene cluster, pah, that encodes the upper pathway for phenanthrene and naphthalene utilization byOUS82. J Bacteriol, 1994, 176(8): 2439–2443.
[30] Takizawa N, Kaida N, Torigoe S, et al. Identification and characterization of genes encoding polycyclic aromatic hydrocarbon dioxygenase and polycyclic aromatic hydrocarbon dihydrodiol dehydrogenase inOUS82. J Bacteriol, 1994, 176(8): 2444–2449.
[31] Denome SA, Stanley DC, Olson ES, et al. Metabolism of dibenzothiophene and naphthalene instrains: complete DNA sequence of an upper naphthalene catabolic pathway. J Bacteriol, 1993, 175(21): 6890–6901.
[32] Takizawa N, Iida T, Sawada T, et al. Nucleotide sequences and characterization of genes encoding naphthalene upper pathway ofPaK1 andOUS82. J Bacteriol Bioeng, 1999, 87(6): 723–731.
[33] Bosch R, García-Valdés E, Moore ERB. Genetic characterization and evolutionary implications of a chromosomally encoded naphthalene-degradation upper pathway fromAN10. Gene, 1999, 236(1): 149–157.
[34] Goyal AK, Zylstra GJ. Molecular cloning of novel genes for polycyclic aromatic hydrocarbon degradation fromGZ39. Appl Environ Microbiol, 1996, 62(1): 230–236.
[35] Zylstra GJ, Kim E, Goyal AK. Comparative molecular analysis of genes for polycyclic aromatic hydrocarbon degradation//Setlow JK, ed. Genetic Engineering. Boston, MA: Springer, 1997: 257–269.
[36] Goyal AK, Zylstra GJ. Genetics of naphthalene and phenanthrene degradation by. J Ind Microbiol Biotechnol, 1997, 19(5/6): 401–407.
[37] Meyer S, Moser R, Neef A, et al. Differential detection of key enzymes of polyaromatic- hydrocarbon-degrading bacteria using PCR and gene probes. Microbiology, 1999, 145(7): 1731–1741.
[38] Moser R, Stahl U. Insights into the genetic diversity of initial dioxygenases from PAH-degrading bacteria. Appl Environ Microbiol, 2001, 55(5): 609–618.
[39] Fuenmayor SL, Wild M, Boyes AL, et al. A gene cluster encoding steps in conversion of naphthalene to gentisate ins sp. strain U2. J Bacteriol, 1998, 180(9): 2522–2530.
[40] Zhou NY, Fuenmayor SL, Williams PA. nag genes of(formerly) sp. strain U2 encoding enzymes for gentisate catabolism. J Bacteriol, 2001, 183(2): 700–708.
[41] Laurie AD, Lloyd-Jones G. The phn genes ofsp. strain RP007 constitute a divergent gene cluster for polycyclic aromatic hydrocarbon catabolism. J Bacteriol, 1999, 181(2): 531–540.
[42] Kiyohara H, Nagao K, Kouno K, et al. Phenanthrene-degrading phenotype ofAFK2. Appl Environ Microbiol, 1982, 43(2): 458–461.
[43] Casellas M, Grifoll M, Bayona JM, et al. New metabolites in the degradation of fluorene bysp. strain F101. Appl Environ Microbiol, 1997, 63(3): 819–826.
[44] Casellas M, Grifoll M, Sabaté J, et al. Isolation and characterization of a 9-fluorenone-degrading bacterial strain and its role in synergistic degradation of fluorene by a consortium. Can J Microbiol, 1998, 44(8): 734–742.
[45] Wattiau P, Bastiaens L, Van Herwijnen R, et al. Fluorene degradation bys sp. LB126 proceeds through protocatechuic acid: a genetic analysis. Res Microbiol, 2001, 152(10): 861–872.
[46] Wang RF, Wennerstrom D, Cao WW, et al. Cloning, expression, and characterization of the katG gene, encoding catalase-peroxidase, from the polycyclic aromatic hydrocarbon-degrading bacteriumsp. strain PYR-1. Appl Environ Microbiol, 2000, 66(10): 4300–4304.
[47] Desta M, Wang WW, Zhang LG, et al. Isolation, characterization, and genomic analysis ofsp. strain SMT-1, an efficient fluorene-degrading bacterium. Evol Bioinform, 2019, 15, doi: 1176934319843518.
[48] Ali F, Hu HY, Wang WW, et al. Characterization of a dibenzofuran-degrading strain of, FA-HZ1. Environ Pollut, 2019, 250: 262–273.
[49] Keasling JD. Gene-expression tools for the metabolic engineering of bacteria. Trends Biotechnol, 1999, 17(11): 452–460.
[50] Henikoff S, Haughn GW, Calvo JM, et al. A large family of bacterial activator proteins. Proc Natl Acad Sci USA, 1988, 85(18): 6602–6606.
[51] Schell MA. Molecular biology of the LysR family of transcriptional regulators. Annu Rev Microbiol, 1993, 47: 597–626.
[52] Rothmel RK, Aldrich TL, Houghton JE, et al. Nucleotide sequencing and characterization ofcatR: a positive regulator of the catBC operon is a member of the LysR family. J Bacteriol, 1990, 172(2): 922–931.
[53] Schell MA. Transcriptional control of the nah and sal hydrocarbon-degradation operons by the nahR gene product. Gene, 1985, 36(3): 301–309.
[54] Schell MA, Wender PE. Identification of the nahR gene product and nucleotide sequences required for its activation of the sal operon. J Bacteriol, 1986, 166(1): 9–14.
Progress in biodegradation of low molecular weight polycyclic aromatic hydrocarbons
Zikang Zhou1,2, Jie Cui1,2, Ping Xu1,2and Hongzhi Tang1,2
1 State Key Laboratory of Microbial Metabolism, and School of Life Sciences & Biotechnology, Shanghai Jiao Tong University, Shanghai 200240, China 2 Joint International Research Laboratory of Metabolic & Developmental Sciences, Shanghai Jiao Tong University, Shanghai 200240, China
Polycyclic aromatic hydrocarbons (PAHs) cause enormous environmental hazards that threaten human health. Bacterial degradation of PAHs has been extensively studied. Bacteria enhanced their biodegradability through multiple levels of regulatory analysis and adaptive evolution to produce diverse catabolic pathways. Based on recent developments, we address here the research progress in bioremediation technology to degrade low molecular weight polycyclic aromatic hydrocarbons.
PAHs, low molecular weight PAHs, bioremediation, fluorene
June17, 2019;
October11, 2019
Science and Technology Commission of Shanghai Municipality (No. 17JC1403300), The “Dawn” Program of Shanghai Education Commission (No. 17SG09), National Key Research and Development Program of China (No. 2018YFA0901200).
Hongzhi Tang. Tel: +86-21-34204066; E-mail: tangzhihong@sjtu.edu.cn
周子康, 崔洁, 许平, 等. 细菌降解低分子量多环芳烃的研究进展. 生物工程学报, 2019, 35(11): 2069–2080.
Zhou ZK, Cui J, Xu P, et al. Progress in biodegradation of low molecular weight polycyclic aromatic hydrocarbons. Chin J Biotech, 2019, 35(11): 2069–2080.
上海市2017年度“创新行动计划”基础研究重点项目 (No. 17JC1403300),上海市曙光计划 (No. 17SG09),国家重点研发计划 (No. 2018YFA0901200) 资助。
(本文责编 郝丽芳)
