康禾自然保护区森林苔藓植物丰富度对地形因子的响应
2019-11-30杨学成吴卓翎周庆徐明锋苏志尧
杨学成,吴卓翎,周庆,徐明锋,苏志尧
华南农业大学林学与风景园林学院,广东 广州 510642
苔藓植物是苔类(liverworts)、藓类(mosses)和角苔类(hornworts)3个植物类群的统称,全世界苔藓植物超过20000种,其物种数仅次于种子植物(Pharoet al.,2007)。在森林生态系统中,苔藓植物不仅是生物多样性的重要组成部分,在维持森林生态系统功能、涵养水源等方面也起着关键作用(Gough et al.,2006;Soudzilovskaiaet al.,2013;田维莉等,2011)。此外,由于苔藓植物具有结构简单、表面积大,缺乏维管组织和真正的根系等独特的生物学特性,表现出对生境变化的高度敏感性,被认为是反映生境特征和森林群落特征及其动态变化的良好指示生物(Bagella,2014)。但相对于维管植物而言,苔藓植物相关的生态学研究还相对缺乏(陈云等,2017)。
揭示环境因子与物种分布和丰富度格局之间的关系是群落生态学研究的经典主题。一般认为生境越复杂,物种丰富度和多样性也越高(李粉霞等,2006;张元明等,2003)。地形因子是植物群落生境异质性的重要来源,因此植物群落的分布及多样性与基于地形划分的生境类型之间存在密切的关系(Chuyonget al.,2011;Takyuet al.,2002)。在山地和丘陵地区,地形因子往往对局部生境温湿度有良好的指示作用,影响光照、水分和土壤的发育,进而影响不同地形下植物群落组成和多样性格局(McDonaldet al.,1996)。多数研究表明,海拔及坡度、坡向等地形因子对大多数维管植物的分布有显著的影响(Homeier et al.,2010;Kebedeet al.,2013;徐远杰等,2017),而苔藓植物分布与地形因子的关联仅有少数文献报道(Henriqueset al.,2016)。雷波等(2004)发现在人工油松林中,坡向对苔藓植物群落组成、多样性差异具有直接的影响;陈云等(2017)则认为苔藓植物群落的物种组成和多样性主要受海拔的影响。然而,相比海拔、坡度和坡向,凹凸度更能反映局部区域的微地形生境,而生长在多样化基质上个体微小的苔藓植物对微生境的响应更为敏感(王挺杨等,2015),因此凹凸度作为重要的地形因子也影响着苔藓植物的分布。目前苔藓植物分布与海拔关联的研究相对较多(Koeniget al.,2015;Songet al.,2015),而关于坡向、坡度和凹凸度等地形因子对苔藓植物丰富度的影响,相关的研究甚少。为此,本文以康禾自然保护区森林苔藓植物群落为研究对象,通过对样地植物和地形因子进行调查与分析,拟探讨以下问题,(1)苔藓植物群落组成和多样性在根据不同地形因子划分的生境类型中是否存在显著差异?(2)苔类和藓类植物分布及丰富度对坡向、坡度和凹凸度等地形因子的响应分别有何规律?(3)对苔藓植物丰富度影响最大的是什么地形因子?
1 研究区概况
康禾自然保护区位于广东省东南部(23°44′—23°53′N,115°04′—115°09′E),属低山陵地带,总体呈东北—西南走向,海拔700 m以上山峰有4座,最高点海拔白石岗 839.7 m。该区处于亚热带季风气候带,年均气温 20.3—21.1 ℃,年均日照时数1810.2—2056.9 h,年均降水量2142.6 mm,年均湿度为78%。主要土壤类型为山地黄壤、红壤和赤红壤等。保护区内植被类型丰富,以亚热带常绿阔叶林为主,优势科为山毛榉科(Fagaceae)及山茶科(Theaceae)(He et al.,2017)。
2 材料与方法
2.1 苔藓植物与地形因子调查
在保护区内选择苔藓植物分布丰富的区域设置6 hm2固定样地,按水平距离分为150个20 m×20 m的样方。以6 hm2固定样地的西南角为原点,采用 Nikon DTM-310全站仪测量并记录每个样方四角顶点的水平距离和高程数据,据此计算坡度、坡向和凹凸度等地形因子的数值。在每个样方中设置5个1 m×1 m的小样方,分别位于样方中心点及对角线1/4、3/4处,调查并记录小样方内苔藓植物的种名、盖度。
每个 20 m×20 m 样方坡向范围为 19.7°—353.2°,坡度范围为 7.9°—45.8°,平均海拔范围为209.1—371.9 m。在北半球由于西南坡的太阳辐射最强,而东北坡的太阳辐射最弱,形成了与坡向相关的热量梯度(McCune et al.,2002)。根据热量从较冷的坡向到较暖的坡向逐渐递增,把样地的坡向可分为3级:阴坡(东坡 68°—112°、东南坡113°—157°、西北坡 293°—337°和北坡 338°—22°),半阳坡(南坡 158°—202°、西坡 248°—292°)、阳坡(西南坡 203°—247°)。坡度由缓到陡可以分为:1级坡度<20°,2级坡度为 20°—30°,3级坡度为 30°—40°,4级坡度>40°。凹凸度的计算采用样方的平均海拔减去与该样方相邻的 8个样方海拔的平均值,而处于样地边缘的样方凹凸度为样方中心的海拔减去4个顶点海拔的平均值(Laiet al.,2010),凹凸度按数值大小划分为3级:1级凹凸度<-1;2级凹凸度为-1—1;3级凹凸度>1。
2.2 数据统计分析
以样地苔类植物和藓类植物的丰富度(即物种数)分别建立数据集(Dataset)。根据地形因子的计算与划分,分别构建与苔类植物和藓类植物丰富度相对应的3级坡向分组变量、4级坡度分组变量、3级凹凸度分组变量的分析数据集。采用Kruskal-Wallis非参数方差分析检验检验苔藓植物丰富度在不同地形分组变量间是否存在显著差异,而对于苔藓植物与地形因子的关联则采用Pearsons相关性分析。用多响应置换过程(Multi-Response Permutation Procedures,MRPP)分析苔藓植物群落组成在不同的地形梯度间是否存在显著差异。方差分析和相关性分析均在Statistica 8.0中运行,MRPP在多元统计分析软件PC-ORD 6.0中运行。
3 结果与分析
3.1 坡向与苔藓植物分布的关联
样地中共调查记录到苔藓植物47种(含3变种),隶属于25科36属;其中苔类有7科11属15种;藓类有18科25属32种。在阴坡、半阳坡、阳坡3个坡向的苔藓植物群落中分别出现9种、14种和 13种苔类植物,分别占样地苔类总物种数的60.0%、93.3%和 86.7%;出现藓类植物分别有 16种、26种和 29种,分别占样地藓类总物种数的50.0%、81.3%和90.6%。3个坡向的苔类植物群落均以细指苔(Kurzia gonyotricha)、双齿裂萼苔(Chiloscyphus latifolius)为优势种,藓类植物群落均以东亚拟鳞叶藓(Pseudotaxiphyllum pohliaecarpum)为优势种。
对不同坡向梯度下苔藓植物丰富度进行Kruskal-Wallis非参数方差分析,结果表明(图1a,图 1c),苔类植物和藓类植物的丰富度在不同坡向分组间均不存在显著差异(P>0.05),说明坡向对苔类和藓类植物丰富度不存在显著的梯度效应。苔类和藓类植物物种丰富度在阴坡的四分位距离最大,说明在阴坡的苔类和藓类植物丰富度分布相对于其他两个坡向较分散,在半阳坡的物种丰富度分布变化范围处于中间水平,阳坡的苔类和藓类植物丰富度变化范围最小。在阳坡的苔类和藓类植物均出现了较多离群值(图1a,图1c)。
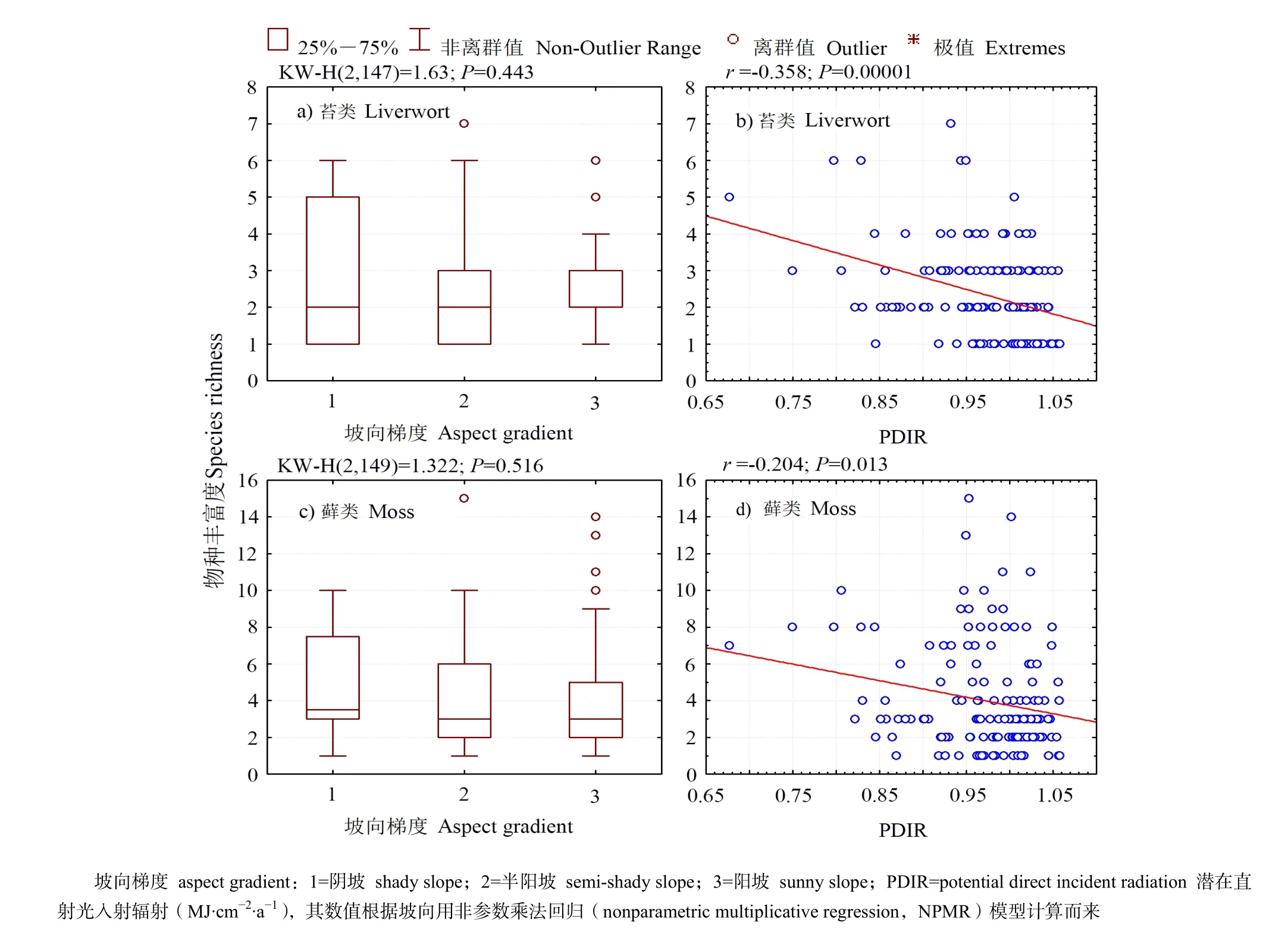
图1 苔藓植物丰富度与坡向的生境关联Fig. 1 Habitat association between bryophyte richness and aspect
相关性分析表明(图1b,图1d),与坡向分组变量不同,基于坡向的潜在直射光入射辐射(PDIR)与苔类植物丰富度存在显著的负线性相关(P<0.01),说明从阴坡到半阳坡和阳坡,随着所接收到的热量增强,苔类植物的物种丰富度显著下降。藓类植物丰富度对PDIR的响应与苔类植物一致,但从相关系数及P值大小来看,苔类植物对连续变量坡向的响应相对藓类植物更为敏感。
进一步对不同坡向苔类和藓类植物群落组成进行的多响应置换程序(MRPP)分析,结果表明3个坡向级的苔类和藓类植物群落组成总体上均不存在显著差异,两两比较的结果也表明各坡向分组间苔类和藓类植物群落组成不存在显著差异(表1,表2)。
3.2 坡度与苔藓植物分布的关联
按照从小到大的4个坡度级分组中(图2),苔藓植物群落分别有苔类植物4种、11种、15种和8种、各占样地中苔类植物总数的 26.7%、73.3%、100%和53.3%;藓类植物各有12种、28种、27种和 15种,分别占样地苔类植物总数的 37.5%、87.5%、84.4%和46.9%。苔藓植物群落在不同坡度的物种组成表现不同。其中,苔类植物在1级坡度的群落组成表现出以细指苔为优势种;2级坡度级苔类植物群落组成以双齿裂萼苔和细指苔为共优势种,变异多褶苔(Spruceanthus polymorphus)和三裂鞭苔(Bazzania tridens)为次优种;3级坡度苔类植物群落以细指苔为优势种,双齿裂萼苔为次优势种;4级坡度级苔类植物群落则表现出以细指苔为优势种,双齿裂萼苔和三裂鞭苔为次优势种的物种组成特征。藓类植物群落在1级坡度表现为以东亚拟鳞叶藓和粗肋凤尾藓(Fissidens laxus)为共优势种,以锦藓(Sematophyllum subpinnatum)为次优种;2级坡度以东亚拟鳞叶藓为优势种,粗肋凤尾藓和细叶小羽藓(Haplocladium microphyllum)为次优种;3级和4级坡度分组的藓类植物群落均表现为以东亚拟鳞叶藓为优势种,粗肋凤尾藓为次优势种的物种组成特征。
表2 藓类植物群落组成随坡向变化的多响应置换程序分析Table 2 Multi-response permutation procedures (MRPP)for the variations in species composition of mosses community across aspect gradients
不同坡度级分组变量对苔类植物和藓类植物丰富度的影响均不存在显著差异(图2,P>0.05)。从图2a来看,苔类植物丰富度在2级坡度的分布相对于其他3个坡度级较分散,丰富度变化范围最大。在2级坡度和3级坡度分别出现了1个和2个样方的丰富度相对于其他样方的丰富度大得多。而藓类植物也是在2级坡度的物种丰富度分布变化范围最大(图 2c)。相关性分析结果表明苔类植物和坡度存在弱线性正相关,藓类植物和坡度存在弱线性的负相关,但相关性均不显著(图2b,图2d)。说明在样地中,坡度不是决定苔藓植物多样性的主要地形因子。
MRPP分析结果表明,苔类植物群落组成在不同坡度下总体存在显著差异(P<10-6),除了3级和4级坡度的苔类植物群落间不存在显著差异,其它坡度分组间均存在显著差异(P<0.05)。且3级和2级坡度间的T值最小,说明2级和3级坡度的分组界限最明显(表3)。而藓类植物群落组成在不同坡度下总体不存在显著差异,且两两比较的结果表明除了2级和3级坡度的藓类植物群落间存在显著差异外,其它坡度分组间均不存在显著差异(表4)。

图2 苔藓植物丰富度与坡度的生境关联Fig. 2 Habitat association between bryophyte richness and slope steepness

表3 苔类植物群落组成随坡度变化的多响应置换程序分析Table 3 Multi-response permutation procedures (MRPP)for the variationsin species composition of liverwort community across slope gradients
3.3 地表凹凸度与苔藓植物分布的关联
在 3个凹凸度分组中,苔类植物分别出现 11种、12种和8种,占样地苔类植物总数的73.3%、80.0%和53.3%;藓类植物各有28种、22种和22种,分别占样地中藓类植物总数的 87.5%、68.8%和68.8%。其中,苔类植物群落组成在1级凹凸度的优势种为细指苔和双齿裂萼苔;2级凹凸度以细指苔为优势种,双齿裂萼苔为次优种;3级凹凸度的苔类植物群落表现为以细指苔为优势种,双齿裂萼苔为次优势种的物种组成特征。藓类植物在不同凹凸度级的群落组成均以东亚拟鳞叶藓为优势种,次优种在不同凹凸度级存在差异。1级凹凸度藓类植物群落次优种为细叶小羽藓和粗肋凤尾藓;2级凹凸度次优种仅有粗肋凤尾藓;而3级凹凸度藓类植物群落次优种为细叶小羽藓。

表4 藓类植物群落组成随坡度变化的多响应置换程序分析Table 4 Multi-response permutation procedures (MRPP) for the variations in species composition of mosses community across slope gradients

图3 苔藓植物丰富度与凹凸度的生境关联Fig. 3 Habitat association between bryophyte richness and convexity
非参数方差分析结果表明(图3a,图3c),不同的凹凸度分组对苔类植物和藓类植物丰富度均存在显著影响(P<0.001),说明样地凹凸度对苔类和藓类植物丰富度的影响表现出显著的梯度效应。且苔类植物和藓类植物丰富度在不同的凹凸度分组间的变化均一致,表现为在1级凹凸度的丰富度分布较分散,变化范围最大,且物种丰富度最高,其次为2级凹凸度,3级凹凸度的物种丰富度最低。从相关性分析来看(图3b,图3d),苔类植物和藓类植物均与作为连续变量的凹凸度存在极显著的负线性相关关系(P<0.0001),说明地表凹凸度越小,苔类植物和藓类植物的丰富度越高。
MRPP分析结果表明,3个凹凸度分组下的苔类植物群落组成总体上存在极显著差异,从两两比较的结果来看,各凹凸度分组间均存在显著差异(表5)。其中3级凹凸度和1级凹凸度间的T值最小,且A值最大,说明苔类植物丰富度在凹凸度分级的1级和3级组界限最强,组内一致性最高。藓类植物群落组成在不同凹凸度分组下总体以及两两比较间均存在显著差异(表6),与苔类植物一样,藓类植物丰富度在凹凸度分级的1级和3级的分组界限最强,组内一致性最高。
表5 苔类植物群落组成随凹凸度变化的多响应置换程序分析Table 5 Multi-response permutation procedures (MRPP)for the variations in species composition of liverwort community across convexity gradients
4 讨论
物种丰富度是生物多样性的一个重要指标,其在自然界的分布状况受到地形因子的影响(Magurran et al.,2011)。在康禾自然保护区森林苔藓植物群落中,总体上地形对苔藓植物的分布模式有重要影响,苔藓植物的丰富度与基于坡向的潜在直射光入射辐射(PDIR)及凹凸度存在显著的线性相关,但坡度和坡向两个地形因子对苔藓植物的丰富度变化并没有表现出显著的梯度效应,这与陈云等(2017)的研究结果一致。而凹凸度则不仅与苔藓植物丰富度表现出显著的线性相关,而且有显著的强梯度效应。

表6 藓类植物群落组成随凹凸度变化的多响应置换程序分析Table 6 Multi-response permutation procedures (MRPP)for the variations in species composition of mosses community across convexity gradients
地形因子对植物群落的影响,实质上是通过控制局部生境的水热条件及土壤养分的再分配,进而对植物群落分布和多样性格局造成影响(Tateno et al.,2003)。已有研究表明,坡度和坡向是影响植物群落分布最重要的地形因子,如区余端等(2011)通过研究地形因子对粤北山地森林不同生长型地表植物分布的影响,发现坡向对蕨类、木质藤本、草本和灌木分布格局的影响均显著,而坡度仅对灌木分布影响显著;Kebede et al.(2013)发现森林群落树种组成和丰富度与海拔和坡向有关。在本研究中,基于坡向的PDIR与苔藓植物丰富度存在显著的负相关关系,但坡度无论对苔类还是藓类植物的分布均不存在显著影响。苔藓植物是一类以孢子繁殖的植物,其个体矮小,结构简单,不具真正的根和维管组织,其独特的生物学性质在一定程度上决定了其对地形因子的响应与维管植物存在一定的差异(Gensel,2008;Oliver et al.,2005)。
在北半球,南坡和西南坡接受到的太阳辐射更强烈,在一天中接受的太阳辐射持续时间最长,而东北坡接收的太阳辐射最弱,因此形成了与坡向相关的热量梯度(Heet al.,2017;McCuneet al.,2002)。而苔藓植物丰富度与基于坡向的PDIR呈显著的负相关,且苔类的相关性更强。从阴坡到半阳坡再到阳坡,热量变高,苔藓植物的丰富度下降,这与苔类植物对生境偏好有关;较冷湿的生境,更适合大多数苔类植物的生长,苔类和藓类植物的多样性更高。
本项研究中,坡度对苔类和藓类植物的分布和多样性均不存在显著的影响。一般来说,坡度决定了土层的厚度。在陡坡,土层较薄,而缓坡的土层较厚(Wanget al.,2014)。坡度和坡向共同决定了土壤的养分和水分。苔藓植物丰富度在不同坡度间不存在显著差异,说明苔藓植物对土层厚度并不敏感。可能的原因是因为苔藓植物不具真正的根,不像大多维管植物可以依靠根系从土壤中吸收水分和养分,而是依靠其巨大的体表面积吸收养分,因此对坡度及土层厚度的响应并不敏感。
此外,在本研究中发现,地表凹凸度对苔类和藓类分布和丰富度均存在显著影响,并且凹凸度是影响藓类植物最主要地形因子。在以往的研究中,往往忽略了凹凸度对植物群落分布的影响。凹凸度是表示水平方向的地形因子,主要通过控制土壤水分存留时间来影响土壤的含水量(郭屹立等,2016)。凹凸度越小越利于水分的存留,生境中的土壤含水量越高,以及存留的凋落物也越多;相反,地表凸的地方较干旱,土壤裸露,养分含量也较低,但是采光相对较好。而苔藓是一类喜阴植物,在有性生殖时,必须借助于水,多数生在阴暗、潮湿的环境中,因此地表凹凸度的大小就能很好的指示苔藓植物丰富度的大小,地表凹凸度值越小,苔藓植物的多样性就越高。
5 结论
本项研究结果表明,苔类和藓类对地形因子的响应各不相同。与坡向相关的潜在直射光入射辐射(PDIR)对苔类植物丰富度的影响强于对藓类植物;而凹凸度对苔类和藓类植物的分布和丰富度变化均有显著的影响,且呈现很强的梯度效应;反映出苔类植物多样性对与坡向相关的热量变化较为敏感,而无论是苔类还是藓类植物均对与水分条件相关的凹凸度变化敏感。然而,苔藓植物的分布和丰富度格局往往是由多个环境因子的综合效应形成的,本研究专门研究了地形因子的影响,揭示了由地形因子引起的苔藓植物分布和丰富度变异,其他未解释的变异则可能还与苔藓植物所处森林的植被类型、结构、土壤性质以及外界干扰等因子有关,有待进一步的研究。
