浑善达克沙地榆树疏林植被和土壤的空间分异特征
2019-11-30张志永朱媛君时忠杰杨晓晖
张志永 ,朱媛君,时忠杰,杨晓晖*
1. 中国林业科学研究院林业科技信息研究所,北京 100091;2. 中国林业科学研究院荒漠化研究所,北京 100091
植被是地球陆地生态系统的主要组成部分(Parmesan et al.,2003)。作为土壤、大气和水体的重要连接体,植被是物质和能量循环的关键环节(陈效逑等,2009)。植被分布受水热条件的影响在大尺度上往往呈现明显的经度地带性和纬度地带性(于永奇,2015)。土壤作为一个不断发展变化的动态生命系统,是包括物理、化学、生物等多种组分的综合体,并且各组分之间相互影响、相互作用,进行着多种复杂的生态过程(Karlen et al.,1997)。土壤作为陆地生态系统的物质基础,是诸多生态过程的载体(康冰等,2010)。土壤的理化性质反映了土壤紧实程度、通透性以及养分状况,不仅作用于土壤的成土过程,同时对土壤动物的活动、植物根系的延伸、养分的转运等过程产生影响(曹国栋等,2013;王文等,2013)。
浑善达克沙地位于中国北方地区的农牧交错带上,长期以来,在气候变化、过度放牧、采伐开垦等因素的影响下,生态退化严重,属于内蒙古中东部的生态敏感区域(曹红芳等,2018)。榆树(Ulmus pumila)疏林是本区域内的主要天然植被类型,也是沙地植被演替的顶级群落(马媛等,2017;Zhang et al.,2019)。榆树疏林在维持浑善达克沙地的生态稳定性方面发挥着重要作用,多年来,学者对浑善达克沙地榆树疏林的种群结构、幼苗更新、空间分布格局、植被生产力、碳氮储量以及土壤微生物等方面均进行了诸多研究(赵云杰等,2014;唐蛟等,2014;赵玮等,2016;马媛等,2017;张志永等,2018)。但是,在整个榆树疏林生态系统尺度上,探讨植被、土壤空间分布特征以及与水热条件之间关系的研究还比较缺乏。
了解植被和土壤的空间分布特征以及与水热条件的关系是理解整个群落物质和能量循环的基础(Ferreira et al.,2007;Rossatto et al.,2014)。因此,本研究在借鉴其他地域疏林草地相关研究经验的基础上,重点了解浑善达克沙地榆树疏林植被的空间分布特征,同时探讨影响植被分布的环境因子。研究成果将有助于深化对榆树疏林基础结构的认识,丰富该区域内的植物共生理论,为研究整个系统的生态过程和生态功能奠定基础,同时,也为该区域的草地恢复工作提供理论依据。
1 研究区概况
研究区位于浑善达克沙地东南部中榆树疏林的主要分布区域(115°29′—116°58′E,42°29′—43°11′N)。该区域属于温带半干旱大陆性季风气候,夏季炎热干燥,冬季寒冷多风,年均温度1.8 ℃左右,年均降水量在250—540 mm之间,且在季节间分布不均,主要集中在夏季的 7—8月间(胡日娜等,2019)。全年盛行西风,年均风速在 3.5—5.5 m·s-1,主要土壤类型是栗钙型风沙土(姚雪玲等,2019)。木本植物主要有榆树、小叶锦鸡儿(Caragana microphylla)、耧斗菜叶绣线菊(Spiraea aquilegifolia)、楔叶茶藨子(Ribes diacanthum)、小红柳(Salix microstachya)、小黄柳(Salix flavida)等。草本植物主要有羊草(Leymus chinensis)、冰草(Agropyron cristatum)、灰绿藜(Chenopodium glaucum)、狗尾草(Setaria viridis)、叉分蓼(Polygonum divaricatum)、委陵菜(Potentilla chinensis)、冷蒿(Artemisia frigida)、赖草(Leymus secalinus)等(张志永等,2017;闫德仁等,2018;姚雪玲等,2019)。
2 研究方法
2.1 研究区域
以正蓝旗的桑根达来镇为中心,将浑善达克沙地榆树疏林划分为东部、南部、西部和北部4个区域,对榆树疏林植被、土壤的空间分异特征进行研究。在每个区域分别设立6个样方,共24个样方,每个样方的空间位置如图1所示。
2.2 样地设置
每个样方采用改进的 Whittaker取样法进行样方设置。改进的 Whittaker取样法具有多尺度性,在干旱、半干旱等空间异质性较大的生态系统具有较高的实用价值,可以较好地反映沙地中斑块状分布的植被资源信息(Stohlgren et al.,1998;喻泓等,2009)。
根据该方法,首先设置面积为1000 m2(20×50 m)的长方形大样方,之后在大样方的中央设置 1个100 m2(5 m×20 m)的样方,大样方的左下角和右上角分别设置1个10 m2(5 m×2 m)的样方,最后在中心样方的4个边分别设置1个1 m2(2×0.5 m)的小样方,在大样方的边缘设置6个1 m2(2×0.5 m)的小样方(长边2个,宽边1个),这6个小样方分别距离大样方边缘 0.5 m,以降低边缘效应对调查结果的影响(梁继业等,2007)。每个改进的Whittaker样方中,面积为 1000、100、10、1 m2的样方数分别为1、1、2、10个。具体设置如图2所示。
2.3 植被调查和取样方法
于2015年8月,调查并记录每个1 m2小样方中草本植物的名称、高度、株/丛数及其总盖度等信息,同时收获草本植物的地上部分,并迅速带回实验室在105 ℃下杀青处理30 min,然后在65 ℃下烘干至恒重。对于10、100和1000 m2分别只记录前一级未出现的植物种类,并对1000 m2大样地内出现的所有木本植物进行调查,利用塔尺测量乔木的树高(精确到0.1 m),用胸径尺测量乔木的胸径(树干1.3 m处)(精确到0.1 cm),用皮尺分东西、南北2个方向测量乔木的冠幅(精确到0.1 m),利用钢卷尺测量灌木(包括无胸径且已经木质化的榆树)的高度(精确到0.1 m),用皮尺分东西、南北2个方向测量冠幅(精确到0.1 m)。

图1 以桑根达来镇为中心的4个区域样方分布图Fig. 1 Distribution map of sampling plots in the four areas with Sanggendalai Town as the center

图2 改进的Whittaker取样法设计图示Fig. 2 The graphical representation of modified-Whittaker sampling plot
同时,在每个样方中,分别在榆树下、灌木丛下以及开阔草地中进行土壤取样,每种类型3个重复。取样时,按0—10、10—20、20—40 cm 3个层次进行。每层土样各自混合均匀后,装于自封袋中,共获得648个样品。
2.4 分析方法
2.4.1 草本群落指标的测定方法
采用针刺法测定各小样方内草本群落的盖度(陈芙蓉等,2013)。采用烘干法测定草本地上生物量(方楷等,2012;干珠扎布等,2015)。
2.4.2 草本群落多样性指数的选择
本研究中,均选择4种α多样性指数(Margalef丰富度指数、Simpson多样性指数、Shannon-Weiner多样性指数、Pielou均匀度指数)作为草本群落物种多样性的评价指标,具体计算公式如下(孔凡洲等,2012)。
Margalef丰富度指数:

Simpson多样性指数:
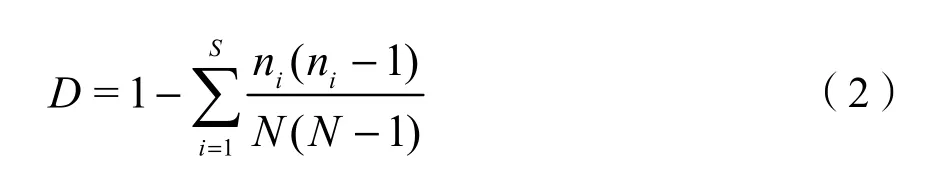
Shannon-Weiner多样性指数:

Pielou均匀度指数:

式中,S为物种数目;N为所有物种的个体数之和;ni为第i个种个体数量。
2.4.3 土壤理化性质的分析方法
野外获取的土壤样品,在室内自然风干后,过2 mm筛子,采用激光粒度分析仪测定土壤的粒径组成。过100目筛子,采用重铬酸钾氧化-外加热法测定土壤有机碳含量,半微量凯氏法测定土壤全氮,HF-HClO4-HNO3消解-ICP法测定土壤全磷(鲍士旦,2000;陈宇轩等,2019;蔡文良等,2019)。
2.5 数据处理
运用Excel 2010软件和Origin 8软件进行数据处理和图表绘制。运用SPSS 18.0软件中的回归分析和Pearson相关系数分析不同因子之间的相关性。
3 结果与分析
3.1 水热条件的空间分异特征
本研究温度和降水量数据(2000—2014年)来源于中国气象科学数据共享服务网。根据本研究区所处的分布范围(图1),选择克什克腾旗和正蓝旗2个气象站点分析东部和南部区域的水热状况,选择正镶白旗和锡林浩特市和 2个气象站点分析西部和北部区域的水热状况。4个区域内的年均温度和年降水量状况如表1所示。由表1可知,4个区域内的年均温度和年降水量具有一定的差异性,但均不显著。东部区域的年均温度最高(3.44 ℃),南部的年均温度最低(3.05 ℃),北部和西部居中(分别为3.28 ℃和 3.15 ℃)。东部区域的年降水量最高(361.39 mm),北部的年降水量最低(289.40 mm),南部和西部居中(分别为343.01 mm和341.01 mm)。

表1 不同区域温度和降水量状况的空间分异特征Table 1 The spatial differentiation of temperature and precipitation in four areas
3.2 榆树种群的空间分异特征
4个区域内榆树的种群密度、胸径、树高和冠幅情况如表2所示。由表2可知,在4个区域间,榆树的种群密度、胸径、树高、冠幅具有一定的差异性。东部区域的种群密度最大(270.00 plant·hm-2),南部的种群密度最小(116.67 plant·hm-2),北部和西部居中(分别为 198.33 plant·hm-2和 141.67 plant·hm-2)。南部区域的胸径最大(26.32 cm),东部区域的胸径最小(16.52 cm),西部和北部居中(分别为24.03 cm和20.97 cm)。南部区域的树高最大(6.82 m),东部区域的树高最小(4.92 m),西部和北部居中(分别为6.58 m和5.75 m)。南部区域的冠幅最大(7.06 m),东部区域的冠幅最小(4.61 m),西部和北部居中(分别为6.44 m和5.77 m)。
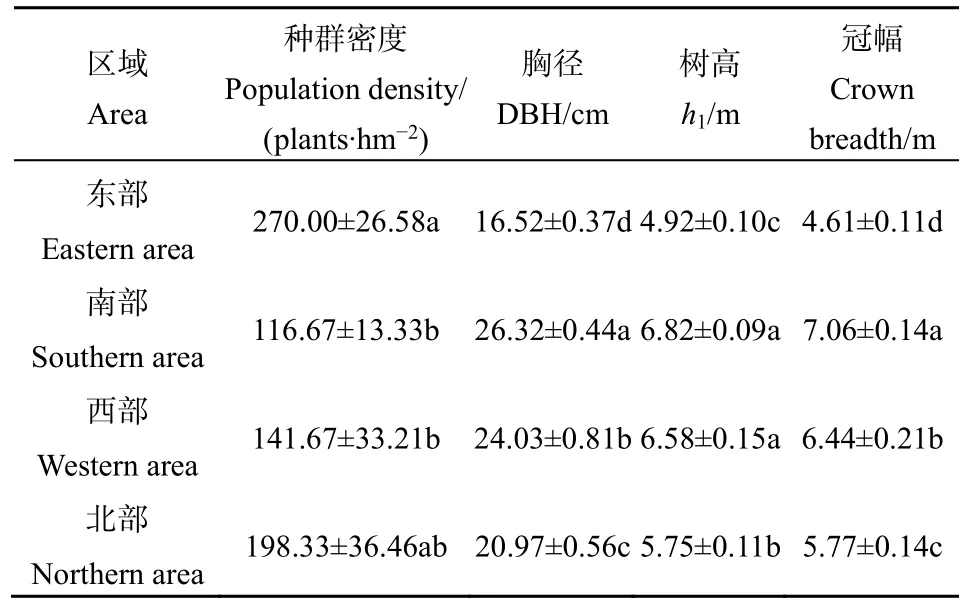
表2 不同区域榆树种群的空间分异特征Table 2 The spatial differentiation of U. pumila L. population in four areas
3.3 灌木群落的空间分异特征
4个区域内灌木群落的物种密度、高度和冠幅情况如表3所示。由表3可知,在4个区域间,灌木群落的物种密度、高度和冠幅具有一定的差异性。西部区域的物种密度最大(415.00 plant·hm-2),北部区域的物种密度最小(238.33 plant·hm-2),南部和东部居中(分别为321.67 plant·hm-2和273.12 plant·hm-2)。北部区域的高度最大(1.14 m),西部区域的高度最小(0.75 m),南部和东部居中(分别为1.08 m和0.93 m)。南部区域的冠幅最大(1.24 m),西部区域的冠幅最小(0.86 m),北部和东部居中(分别为1.21 m和1.12 m)。
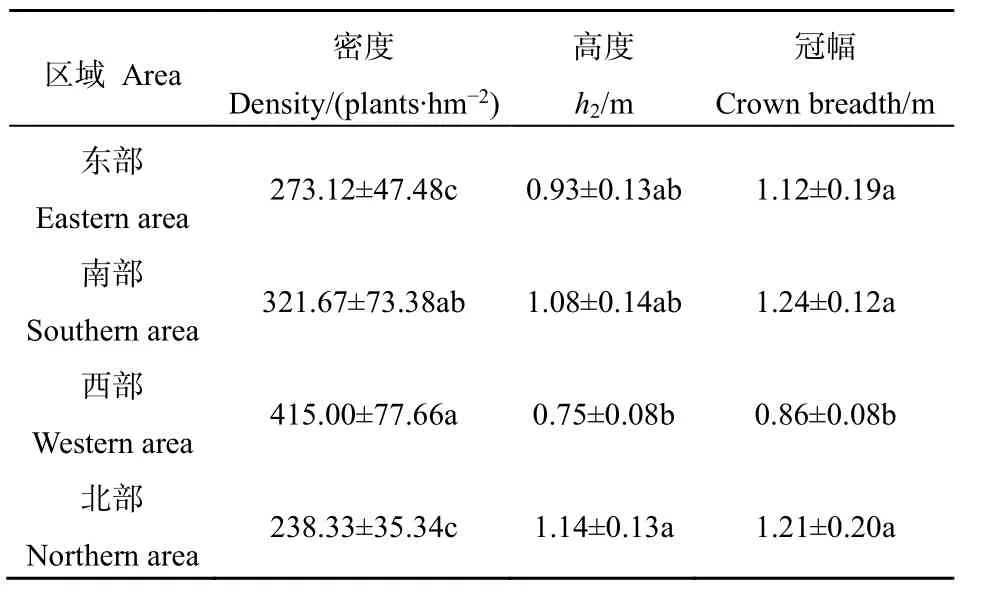
表3 不同区域灌木群落的空间分异特征Table 3 The spatial differentiation of shrub community in four areas
3.4 草本群落的空间分异特征
3.4.1 草本群落结构指标的空间分异特征
4个区域内草本群落的物种数、地上生物量、盖度和高度情况如表4所示。由表4可知,在4个区域间,草本群落的物种数、地上生物量、盖度和高度具有一定的差异性。北部区域的物种数最多(16种),西部的物种数最少(13种),东部和南部居中(分别为16种和14种)。南部区域的地上生物量最大(212.15 g·m-2),东部的地上生物量最小(108.45 g·m-2),西部和北部居中(分别为 178.11 g·m-2和 169.53 g·m-2)。西部区域的草本盖度最大(69.92%),东部区域的草本盖度最小(56.97%),北部和南部居中(分别为68.31%和63.69%);南部区域的高度最大(18.68 cm),东部区域的高度最小(12.35 cm),西部和北部居中(分别为18.63 cm和18.37 cm)。
3.4.2 草本群落物种多样性的空间分异特征
4个区域内草本群落的物种多样性指数如表 5所示。由表5可知,在4个区域间,草本群落的物种多样性指数具有一定的差异性,但大多数未达到显著性水平。北部区域的Margalef指数最大(1.34),南部区域的Margalef指数最小(0.99),西部和东部居中(分别为 1.12和 1.11)。南部区域的 Simpson指数最大(0.64),西部区域的 Simpson指数最小(0.57),东部和北部居中(分别为 0.60和 0.58)。南部区域的Shannon指数最大(1.32),西部区域的Shannon指数最小(1.22),东部和北部居中(均为1.26)。南部区域的Pielou均匀度指数最大(0.69),北部区域的Pielou均匀度指数最小(0.56),东部和西部居中(分别为0.62和0.57)。

表4 不同区域草本群落物种数、地上生物量、盖度和高度的空间分异特征Table 4 The spatial differentiation of species, aboveground biomass,herbage coverage and height in four areas
3.5 土壤粒径组成的空间分异特征
4个区域内土壤粒径组成的空间分异特征如表6所示。由表6可知,在4个区域间,土壤粒径组成的粘粒、粉粒、砂粒含量具有一定的差异性,仅有东部区域的粉粒和砂粒含量与其他区域呈现显著性差异。西部区域的粘粒质量分数最高(0.27%),东部区域的粘粒质量分数最低(0.19%),南部和北部居中(分别为0.26%和0.25%)。西部区域的粉粒质量分数最高(7.78%),东部区域的粉粒质量分数最低(3.35%),北部和南部居中(分别为7.06%和6.11%)。东部区域的砂粒质量分数最高(96.45%),西部区域的砂粒质量分数最低(91.95%),南部和北部居中(分别为93.62%和92.70%)。
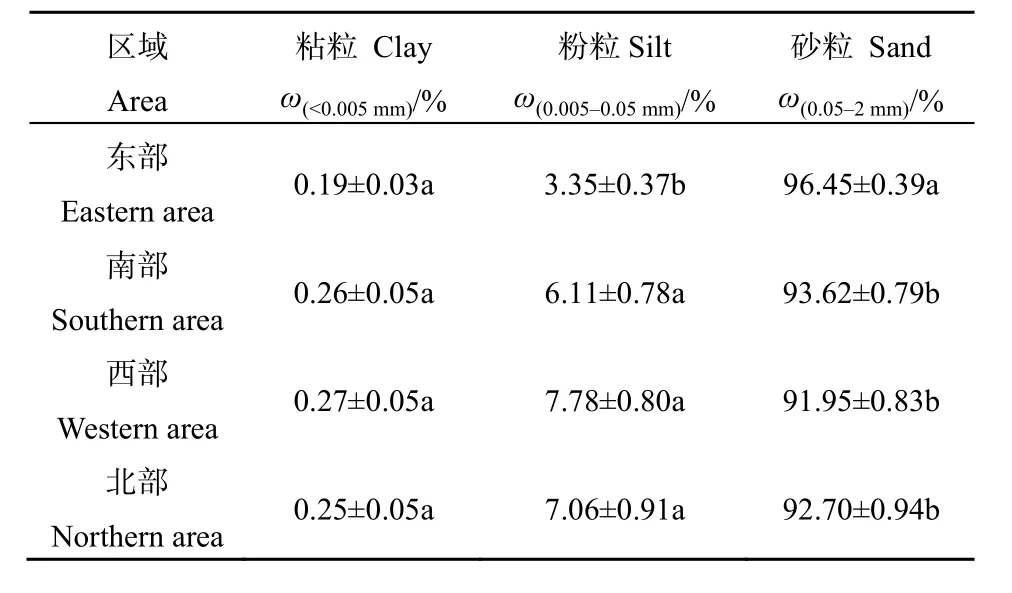
表6 不同区域土壤粒径组成的空间分异特征Table 6 The spatial differentiation of soil particle size in four areas
3.6 土壤化学性质的空间分异特征
4个区域内土壤化学性质的空间分异特征如表7所示。由表7可知,在4个区域间,土壤有机碳、全氮、全磷含量具有一定的差异性。北部区域的有机碳含量最高(15.12 g·kg-1),东部区域的有机碳质量分数最低(7.58 g·kg-1),西部和南部居中(分别为 11.55 g·kg-1和 9.54 g·kg-1)。南部区域的全氮质量分数最高(4.37 g·kg-1),东部区域的全氮质量分数最低(0.74 g·kg-1),北部和西部居中(分别为1.46 g·kg-1和 1.24 g·kg-1)。北部区域的全磷质量分数最高(0.33 g·kg-1),东部区域的全磷质量分数最低(0.19 g·kg-1),西部和南部居中(分别为 0.30g·kg-1和 0.27 g·kg-1)。

表7 不同区域土壤化学性质的空间分异特征Table 7 The spatial differentiation of soil chemical properties in four areas

表5 不同区域草本群落物种多样性指数的空间分异特征Table 5 The spatial differentiation of diversity index in four areas
3.7 榆树种群与灌木、草本群落各指标间的相关性分析
将榆树种群特征分别与灌木、草本群落各结构指标之间进行Pearson相关性分析,结果如表8所示。由表8可知,灌木群落的密度分别与榆树的种群密度呈现显著的负相关关系(P<0.05),与榆树的胸径呈现显著的正相关关系(P<0.05),与树高呈现极显著的正相关关系(P<0.01),与冠幅呈现极显著的正相关关系(P<0.01)。灌木群落的高度、冠幅以及草本群落的物种数、地上生物量、盖度和高度均未与榆树种群各指标之间呈现出显著的相关关系。
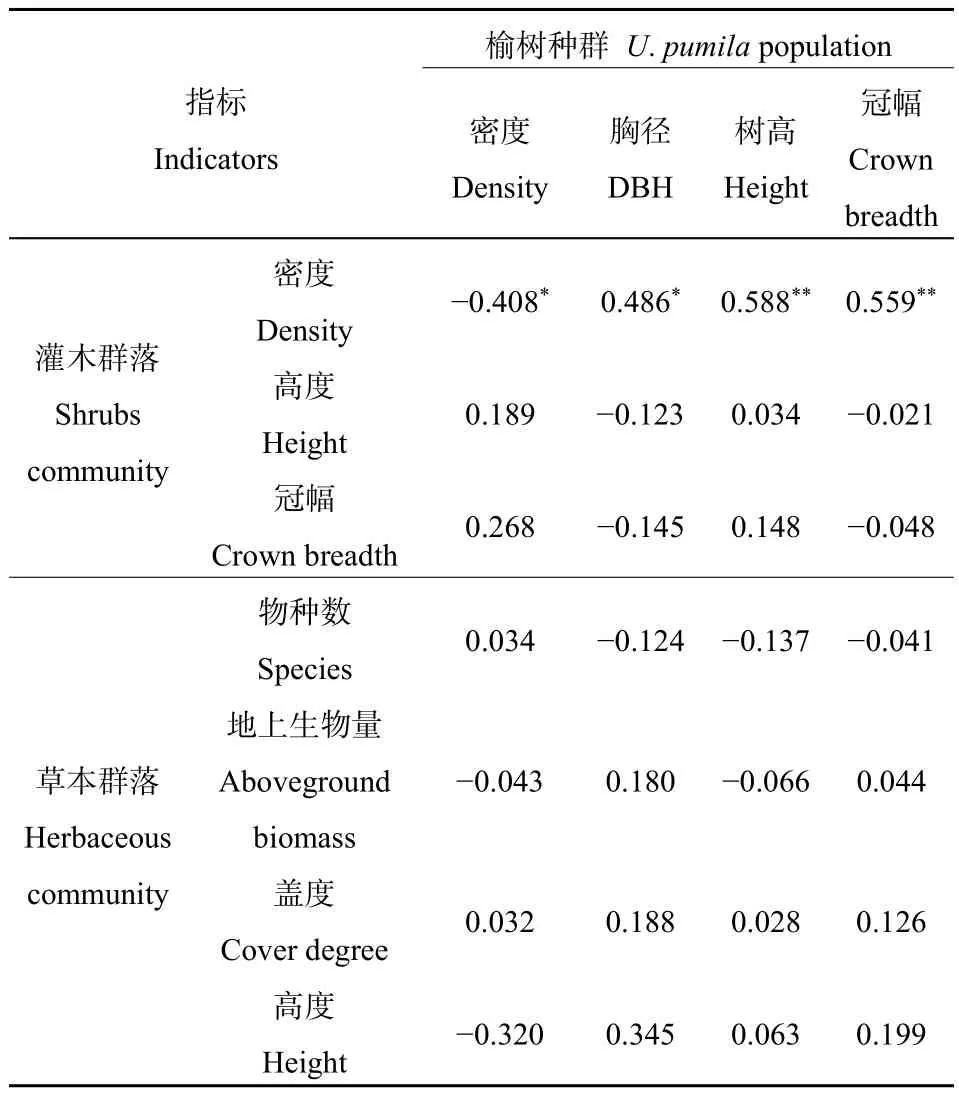
表8 榆树种群与灌木、草本群落各指标间的相关性分析Table 8 Correlative coefficient between U. pumila population indicatorsand shrubs community, herbaceous community
3.8 植被和土壤的相关性分析
将植被的各项指标与土壤理化性质进行Pearson相关性分析,结果如表9所示。由表9可知,榆树种群和灌木群落的各项指标与土壤理化性质之间未呈现出显著的相关关系,草本群落的部分指标与土壤理化性质之间呈现显著的相关关系。具体表现为:土壤粘粒含量分别与草本群落的物种数、地上生物量、盖度和高度呈现显著的正相关关系(P<0.05),土壤粉粒含量与草本群落的物种数呈现极显著的正相关关系(P<0.01),土壤砂粒含量与草本群落的物种数呈现极显著的负相关关系(P<0.01),土壤有机碳、全氮、全钾含量分别与草本群落的物种数呈现极显著的正相关关系(P<0.01),土壤有机碳、全氮、全钾含量分别与草本群落的地上生物量呈现显著的正相关关系(P<0.05)。
4 讨论
植被和土壤作为生物圈的底层,在空间上受到降水和温度的深刻影响。水热条件是影响植被和土壤空间分布的主要非生物因子(陈效逑等,2009)。温度和降水可以通过有效积温和可利用水分对植物的生长周期、生长速率等生理过程造成影响。一般而言,水热条件在大尺度上的空间异质性对植被和土壤的空间分布特征具有重要影响,水热条件的差异性往往会在空间上造成植被分布的空间异质性。
研究发现,榆树在东部区域具有更大的种群密度,这与年均温度呈现出一致的空间分布规律,而与年降水量并没有体现出良好的空间分布特征,表明在浑善达克沙地榆树疏林中,温度可能是影响榆树种群空间分布的重要环境因子,榆树自身具有发达的根系,可以有效地利用地下水,从而摆脱降水量对生长的限制,降水量对榆树种群空间分布的影响可能相对较弱。榆树的种群密度分别与胸径、树高、冠幅呈现相反的空间分布规律,随着榆树个体的生长,种群密度逐渐降低,推断榆树种群存在“自疏”现象。

表9 植被指标与土壤理化性质的相关性分析Table 9 Correlative coefficient between vegetation indicators and soil physicochemical properties
灌木的群落密度与榆树的种群密度之间呈现出显著的负相关关系(表8),表明在整个区域内,榆树与灌木在空间上存在不同的分布区域,在该区域内榆树的存在可能会对灌木的生长产生抑制作用。但是长期来看,如果气候干旱加剧,同时缺乏有效的恢复措施,榆树的种群更新未达到良性发展,榆树的数量则可能会减少,那么灌木的数量则可能出现增加的趋势,最终可能占据整个疏林草地系统。
在东部区域中,水热条件相对较好,但是土壤的沙粒含量则相对较高,土壤的养分含量以及草本群落的地上生物量、盖度和高度相对较低。关于这一点,分析认为存在2个方面的原因,一是草本群落和土壤理化性质可能受局部小气候的影响更大,在本文的研究尺度下,水热条件的影响更大地体现在对木本植物的作用上;二是东部区域的地形起伏较大,基本处于内蒙古高原中达里湖向西南一线的丘陵地带之上,相对于其他区域更为平缓的地形而言,东部区域表层土壤的基本条件较差,草本植物和土壤之间的良性促进作用难以有效开展。
相关性分析表明,土壤理化性质与草本群落的部分指标之间存在显著相关关系,草本群落与土壤理化性质的空间分布呈现出较好的一致性(表9),说明土壤理化性质可能是影响草本群落空间分布的主要作用因子。已有的研究表明,在干旱、半干旱环境中,土壤的空间分布常与草本群落的空间分布具有较强的相关性(Meglioli et al.,2017)。土壤可以通过水、热、气、肥等调节植物的生长,影响植物的生长(John et al.,2007),另一方面植物的生命活动(尤其是根系)又可以改善土壤结构,促进土壤的成土过程,进而影响土壤的理化性质(Sauer et al.,2006)。
由于本研究区域范围较小,该区域内未发现有效的气象站点,因此,研究采用了克什克腾旗和正蓝旗 2个气象站点分析东部和南部区域的水热状况,选择正镶白旗和锡林浩特市和2个气象站点分析西部和北部区域的水热状况。研究具有一定的局限性,难以准确地反映该区域内水热条件的空间分布状况,但仍具有一定的参考价值。榆树疏林生态系统中植被和土壤的空间分布与水热等气候因子的关系还需进一步的研究证实。
5 结论
榆树的种群密度呈现由东部向北部、西部和南部逐渐下降的变化趋势,并且存在“自疏”现象,灌木群落与榆树种群呈现相反的空间分布格局。在当前的气候条件下,温度是影响榆树种群空间分布的重要环境因子,土壤对草本植物空间分布的影响较大,而对本木植物(包括乔木和灌木)空间分布的影响较弱。
