黄河源河岸带植物根-土复合体抗蚀特性研究
2019-11-23朱海丽谢彬山李本锋胡夏嵩李国荣
李 可,朱海丽,谢彬山,李本锋,胡夏嵩,李国荣
(1.甘肃省交通规划勘察设计院股份有限公司,甘肃兰州730030;2.青海大学地质工程系,青海西宁810016;3.青藏高原北缘新生代资源环境重点实验室,青海西宁810016)
黄河源区地处青藏高原东北部,区内发育着众多自由弯曲河流,高寒草甸、高寒草原化草甸植被群落发育良好,覆盖度较高[1]。2011—2017年对黄河源区河流地貌野外调查表明,弯曲河流在季节性冻融及水流冲蚀作用下,凹岸崩塌现象较为严重,河湾每年产生0.5~1.0 m的横向迁移,崩落至近岸边的根-土复合体在水流浸泡及冲刷作用下缓慢崩解、分散,能够在一定时间内保护坡脚免受水流冲刷,从而减缓河岸的崩塌。因此,定量化对比分析黄河源区高寒草甸根-土复合体在冻融循环作用前后的抗水蚀特性,对于定量分析弯曲河流横向演变过程、崩岸速率等具有重要意义。
土壤抗侵蚀性可分为土壤抗冲性和土壤抗蚀性[2-4],其中土壤抗蚀性是指土壤抵抗水的分散和悬浮的能力[5],是评定土壤抵抗侵蚀能力的重要参数,与土壤的理化性质密切相关。国内外学者开展的相关试验研究表明:土体在水中的变化有缓慢崩解、指数崩解、完全崩解三个阶段[6-7];土壤抗蚀性不能由单一指标来评价[8];土壤生物学指标以及土壤内在的物理性质即结皮强度、有机质含量、孔隙状况、水稳性团聚体含量、土壤颗粒组成等,对土壤抗蚀性的影响较为明显[9-12];植被有效根密度及在土壤剖面中的盘绕能够提高土壤的透水性,并增强土壤的抗冲性[13-16];高寒草甸植被多为密丛性根系植物,根径≤1 mm的须根较多[17],因此对提高土体抗蚀性具有重要作用;季节性冻融侵蚀是黄河源区常见的一种土壤侵蚀类型[18];长期的冻融作用,使黄河源区河岸植被完整性遭到破坏,河岸冻胀裂缝发育,在水流的冲刷下河岸土体易坍塌[19];土体结构在冻融作用下受到很大的破坏,土的孔隙度减小,但渗透性有所增强,在经过30次冻融循环后逐渐趋于稳定[20-23];随着冻融循环次数的增加,黏土的黏聚力逐渐下降[24-25]。关于冻融作用的研究,主要集中在土体的物理特性和力学参数等方面,而对冻融作用影响根-土复合体抗水蚀的相关研究较少。因此,笔者以黄河源区高寒草甸区河岸根-土复合体为研究对象,通过原位取样和室内试验,对比分析试样冻融循环前后的抗水蚀指数,定量研究冻融作用下黄河源区高寒草甸根-土复合体的抗水蚀性能的变化规律,深入揭示不同植被类型和盖度、不同冻融循环次数对根-土复合体抗蚀性的影响,以期为黄河源区河岸保护等提供参考。
1 研究区自然概况
研究区位于青海省东南部黄南藏族自治州河南县以南约 50 km 处,其地理坐标为北纬 34°26′—35°2′、东经101°29′—101°35′。 该区域海拔3 400~4 200 m,属高原亚寒带湿润气候区,冷季漫长多大风,暖季短暂,气候湿润多雨,干湿季节分明。研究区年均气温在-4℃以下,最低气温为-31.89℃,属季节性冻土区,年平均冻土深度为137.87 cm,季节冻土冻结时间年平均为149 d。多年平均降水量为560 mm,多集中在5—9月。研究区河岸带植被主要为高寒草甸,其中华扁穗草、线叶嵩草、西藏嵩草和垂穗披碱草等草本植物分布较广且生长良好,为主要优势草种,灌木种以金露梅、银露梅和西藏沙棘为多见[1]。
2 试验研究方法
2.1 取样与试验概况
2016年6月25—28日在研究区河岸带选取优势植物华扁穗草、金露梅大量分布的区域,选定3种不同植被覆盖度样地作为试验取样点,此外选取1个植被覆盖度小于5%的秃斑地作为裸露地取样点,4个取样点分别为裸露地(1号)、覆盖度为60%的华扁穗草地(2号)、覆盖度为90%的华扁穗草地(3号)、覆盖度为90%的金露梅+杂草地(4号)。用铁锹挖取直径为30 cm、地表以下深度为25 cm的圆柱形土样,放入取样盆中带回实验室。依据地下根量变化的特点,对取样盆中的土样按表层往下0~5、6~10、11~15 cm 三个取样深度用削土刀切取长×宽×高为10 cm×4 cm×5 cm的小块试样,按冻融循环次数为0、3、6、9、12次分别进行重复取样,将取得的小块试样装入自封袋并贴好标签后放入BD/BC-148JC型冰箱中进行冻结。依据2001—2015年10月—次年4月的气温变化情况,设定冻结温度为-18℃、融解温度为10℃。将抗水蚀试验的试样放入预设温度为-18℃的冰箱冻结24 h,然后置于预设温度为10℃的LRHS-150-III型恒温恒湿培养箱融解24 h,此过程为一次冻融循环,按照此方法和试验设置完成0、3、6、9、12次冻融循环。在挖取土样过程中,按0~5、6~10、11~15 cm 三个深度用环刀原状制取根-土复合体,测定其天然密度、根面积比、根长密度、根体积比、含水率和有机质含量,其中有机质测定采用重铬酸钾容量法。试样基本物理性质指标测定结果见表1。
将完成冻融循环次数的试样放入托盘中自然风干,然后依次过5、2 mm的筛,留取直径2~5 mm的土颗粒90粒作为试样,分三批均匀置于自制简易水蚀装置中浸泡,该装置[5]包括托盘(20 cm×20 cm×3 cm) 、网架(15 cm×15 cm ×2 cm)、烧杯(1 000 mL)。 观察土颗粒在静水中的崩解现象,当土颗粒表面出现大的裂缝,并在轻微触碰下碎裂时,认为已完全崩解,每隔1 min记录崩解土颗粒个数,连续观察记录10 min,并记录最终的崩解土颗粒个数。依次完成冻融前后不同植被覆盖度、不同取样深度和不同冻融次数的根-土复合体的崩解试验,每组试验重复3次。
2.2 数据处理
土体抗蚀指数指在规定时间内未崩解土颗粒个数占总颗粒数的比例[10],计算公式为

式中:S为土体抗蚀指数,%;ω为土粒总数,个;φ为崩解土粒数,个。
采用Excel进行数据处理和分析,采用SPSS21.0对数据进行单因素方差分析。试验数据的分析、统计于2017年4—5月完成。

表1 试样基本物理性质指标测定结果
3 试验结果分析
将挑选出的土颗粒缓慢放入水中后,土颗粒表面的微小分散土粒会立即融入水中,使土颗粒周围水体浑浊,少量土颗粒偶尔会有气泡逸出,接着土颗粒的松散表面开始崩解。试验刚开始时土颗粒崩解速度较快,随后逐渐减慢。随着试验的进行,土壤结构逐渐被破坏,土颗粒体积膨胀,表面慢慢出现裂缝,接着土颗粒快速崩解,但很快崩解速度又减缓,直至完全崩解。也有少部分土颗粒崩解所需时间短、速度快,在整个崩解过程中崩解速度变化较小。试验结果统计见表2。
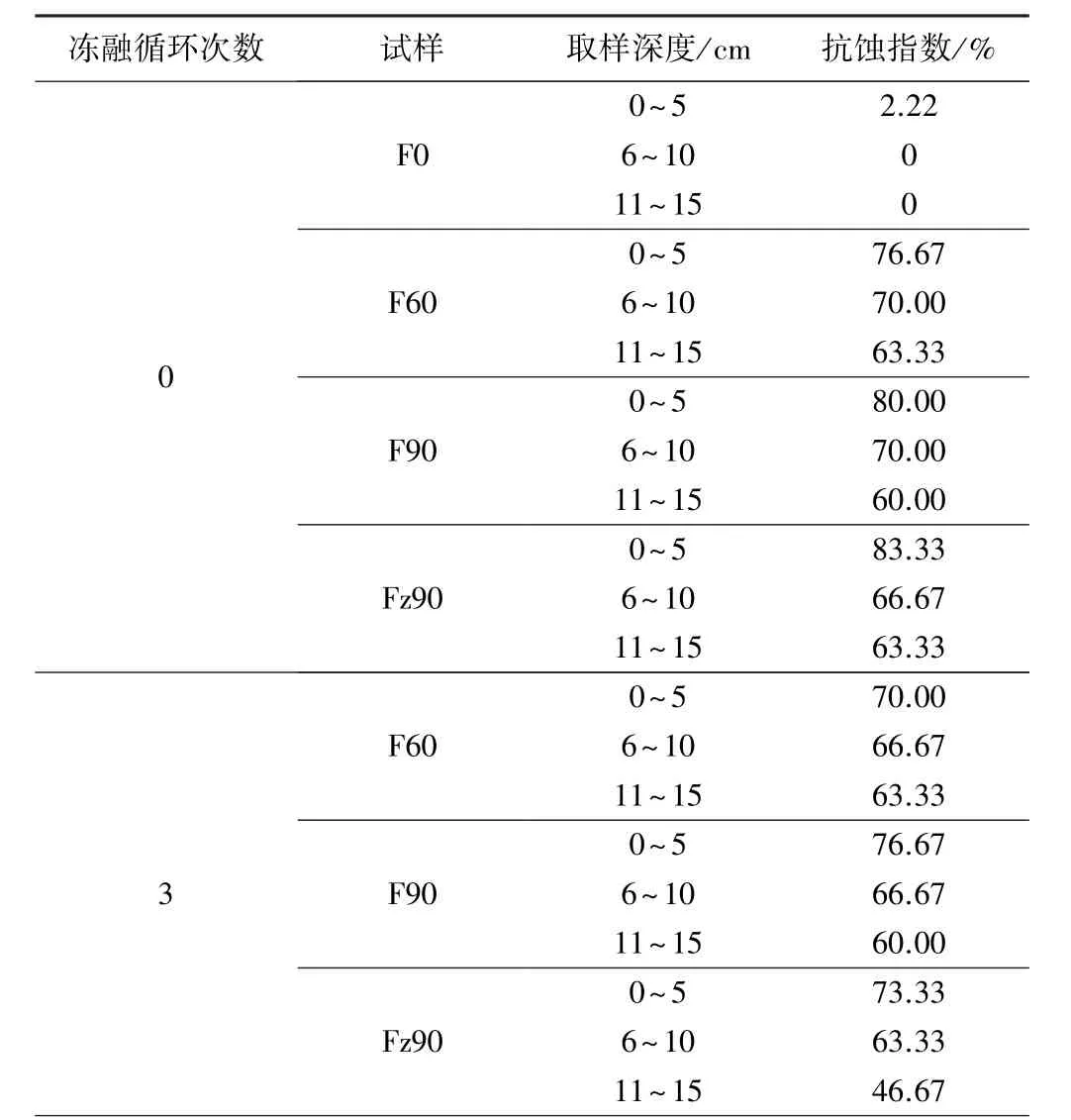
表2 土体抗蚀试验结果统计

续表2
3.1 土体抗蚀指数与植被种类及覆盖度的关系
通过试验发现:研究区未经冻融循环的裸露地(1号取样点)0~5 cm土层3组土颗粒在试验开始8 min左右时有2组土颗粒全部崩解,剩下的1组在浸泡10 min后未崩解土颗粒仅剩2个,试样整体抗蚀指数仅为2.22%,而6~15 cm土层的抗蚀指数为0%。含根土体的抗蚀指数均远大于不含根的,植被覆盖度为90%的金露梅+杂草地(4号取样点)0~5 cm土层土体抗蚀指数最大,达到83.33%;覆盖度为90%的华扁穗草地(3号取样点)0~5 cm土层土体抗蚀指数次之,为80.00%;覆盖度为60%的华扁穗草地(2号取样点)0~5 cm土层土体抗蚀指数为76.67%,略小于覆盖度较高的2种样地的。植被覆盖度影响土体抗蚀指数的根本原因在于,不同植被覆盖度高寒草甸土壤中根系含量不同。
绘制未冻融试样土颗粒崩解个数与时间的关系曲线(图1)可以看出:试样土颗粒崩解个数随着样地植被覆盖度的提高呈现减少的趋势;祼露地试样土颗粒崩解个数在试验开始后4 min内迅速增加,之后增速减缓并在6 min后趋于稳定;覆盖度为90%的华扁穗草地土颗粒崩解量在7 min时达到峰值;而覆盖度为60%的华扁穗草地与覆盖度为90%的金露梅+杂草地试样土颗粒崩解个数与时间基本呈递增关系,且覆盖度为60%的华扁穗草地试样崩解速率相对较快,表明莎草科植物华扁穗草地密集的根系使根、土间的黏结性大大增强,从而增强试样的抗蚀性。经SPSS 21.0单因素方差检验(见表3)表明,研究区不同植被覆盖度样地土体抗蚀指数具有极显著差异性。通过对比表1中不同样地类型的根系参数可知,植被覆盖度为90%的华扁穗草地试样的根面积比、根长密度和根体积比与覆盖度为60%的华扁穗草地、覆盖度为90%的金露梅+杂草地、裸露地3种试样的相比,平均高出0.30、2.40、0.76 个百分点,较多的根系能够更有效地胶结土体,因此植被覆盖度为90%的华扁穗草地表现出较强的抗蚀能力。

图1 不同植被覆盖度试样土颗粒崩解个数与时间的关系

表3 土体抗蚀指数的单因素方差检验结果
3.2 土体抗蚀指数与有机质含量的关系
由不同植被种类及覆盖度的未冻融试样水蚀试验及有机质含量测定结果(见表4)可知,4种样地不同深度土壤的有机质含量不同,有机质含量随深度增加总体呈递减趋势,0~5 cm表层土壤有机质含量较高,其中:植被覆盖度为90%的金露梅+杂草地表层试样有机质含量最高,为91.73 g/kg;裸露地表层试样有机质含量最低,为17.47 g/kg。随着有机质含量的降低,抗蚀指数呈减小的趋势。由表1可知,随着深度的增加,11~15 cm土层试样根面积比、根长密度、根体积比与0~5 cm表层试样相比,平均分别下降了0.15、1.79、0.54个百分点,由于表层植物根系发育,土壤中有机质含量相对较高,能凝聚、胶结更多的水稳性团聚体[15],且活性根系对温度的传导具有一定的阻隔效应,土颗粒之间冷热传导速度减缓,因此表层土壤的抗蚀性大于下层土壤的。土体含水量随土层深度的增加逐渐降低,4种样地下层试样含水率平均减小6.18个百分点,在冻融循环过程中,表层土体中较多的水分通过冻融作用对土体中团聚体的破碎效应强于下层土体,且表层土壤孔隙与空气接触,在冻结过程中水分相变增加的体积和气体收缩的体积接近时,团聚体之间或内部的压力达到平衡,减缓团聚体破碎[26],从而使表层土壤的抗蚀指数大于下层土壤的。
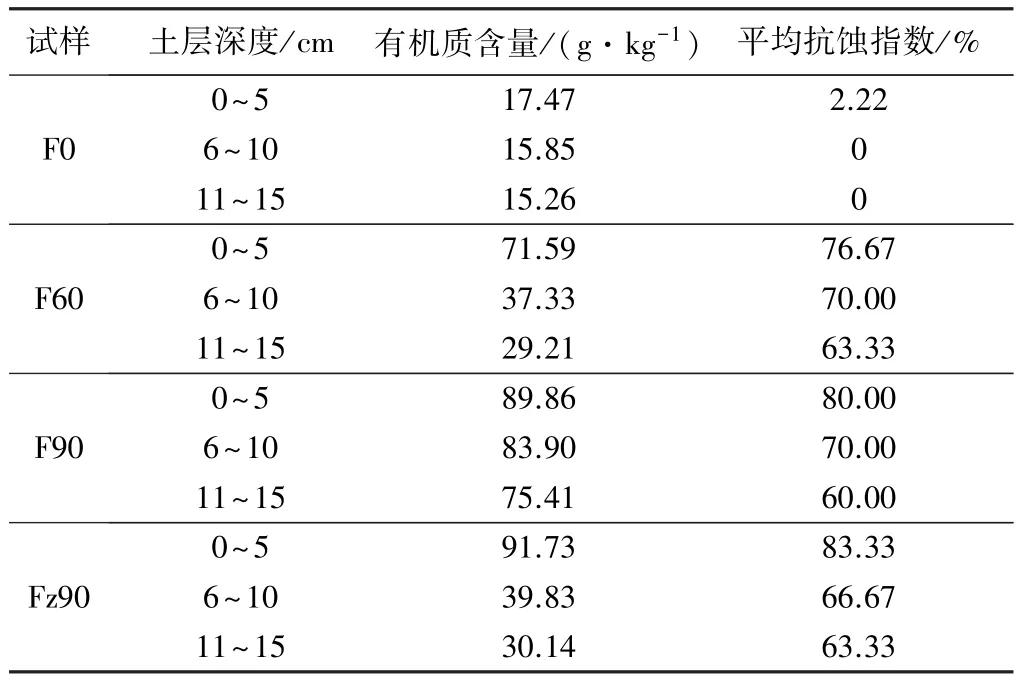
表4 土体抗蚀指数与有机质含量统计
3.3 土体抗蚀指数与冻融循环次数的关系
绘制土体抗蚀指数与冻融次数关系曲线(见图2)可知,随着冻融次数的增加土体抗蚀指数总体呈减小的趋势。由图2(a)可以看出,覆盖度为60%的华扁穗草地0~5 cm土层和6~10 cm土层试样的抗蚀指数随着冻融循环次数的增加趋于线性减小的趋势,11~15 cm土层试样的抗蚀指数在冻融9次时减小幅度达29.42%;由图2(b)可以看出,覆盖度为90%的华扁穗草地0~5 cm土层和6~10 cm土层试样的抗蚀指数在冻融9次时降至最低,而11~15 cm土层试样的抗蚀指数在冻融6次时开始减小、在冻融12次时降至最低,整体上看冻融循环对覆盖度为90%的华扁穗草地上层土体抗蚀指数影响相对较小,在冻融循环12次后土体抗蚀指数减小幅度仅为8.33%;由图2(c)可以看出,覆盖度为90%的金露梅+杂草地不同深度土层试样的抗蚀指数随着冻融循环次数的增加总体呈减小的趋势,且深度越大试样抗蚀指数减小的幅度越大。试样经多次冻融循环后,土壤有机质含量随着温度的变化以指数形式下降[27],土壤中的水稳性团聚体相应减少,因此试样的抗蚀指数随着冻融循环次数的增加总体呈减小趋势。

图2 土体抗蚀指数与冻融循环次数的关系
土壤在反复冻融循环作用下,自身的物理性质、生物学指标均会发生变化,势必影响土壤的抗蚀指数[24]。由不同覆盖度、相同深度土体抗蚀指数与冻融循环次数的关系曲线比较可知,不同覆盖度、相同深度试样的抗蚀指数整体随着冻融循环次数的增加而减小,其中:深度为11~15 cm试样的抗蚀指数减小幅度较大,覆盖度为60%的华扁穗草地试样经过12次冻融循环后,抗蚀指数下降幅度最大,为44.21%,覆盖度为90%的金露梅+杂草地试样抗蚀指数下降幅度为42.1%、覆盖度为90%的华扁穗草地抗蚀指数下降幅度为38.88%。冻融次数超过9次后,试样抗蚀指数减小幅度变小并在冻融循环次数达到12次时,部分土颗粒抗蚀指数增大,原因是经过多次冻融循环之后土颗粒内部结构达到了新的平衡状态,试样的抗蚀性相应增强。冻融循环次数较少时,破碎的土颗粒阻挡土体裂缝的产生,使得土壤内部裂隙减少、阻止水分的渗入,随着冻融次数的增加,冻融作用的累加效果逐渐显现,土体中水分冻结体积增大,改变土壤颗粒的位置,使得土壤孔隙增大,融化的过程中又使得细小的土颗粒流失,土壤变得更加松散,抗蚀指数随之减小;但当冻融次数增加到9次后,试样抗蚀指数减小趋势减缓,说明当冻融达到一定次数后冻融作用对试样抗蚀性的影响不再显著。
不同深度、冻融循环次数试样抗蚀指数的单因素方差分析结果(见表5)表明,冻融循环次数对覆盖度为60%的华扁穗草地土体抗蚀指数的影响具有极显著性(P<0.01),对覆盖度为 90%的金露梅+杂草地土体抗蚀指数的影响具有显著性(P<0.05),对覆盖度为90%的华扁穗草地土体抗蚀指数的影响不显著(P=0.10)。

表5 不同深度、冻融循环次数试样抗蚀指数的单因素方差分析结果
4 结 论
(1)不同样地的植被种类和覆盖度、土体含根量、有机质含量等均不相同,对应试样的抗蚀指数也不相同;土体抗蚀指数随着植被覆盖度、有机质含量的提高而增大,随土层深度的增加而减小;冻融作用对覆盖度为60%的华扁穗草地与覆盖度为90%的金露梅+杂草地土体抗蚀指数影响显著,对覆盖度为90%的华扁穗草地土体抗蚀指数影响不显著。
(2)未经冻融情况下,植被覆盖度为90%的华扁穗草地试样的崩解速率缓慢,抗蚀指数比覆盖度为60%华扁穗草地与裸露地试样分别增大3.33%和77.78%,与覆盖度为90%的金露梅+杂草地试样的接近。冻融情况下,各样地土体抗蚀指数变化情况各不相同,在冻融循环达到9次之后,土体抗蚀指数减小幅度变缓,并有回升的趋势,冻融循环对覆盖度为90%的华扁穗草地土体抗蚀指数影响相对较小,在冻融循环12次后土体抗蚀指数减小幅度仅为8.33%。
(3)植被覆盖度为90%的华扁穗草地试样的根面积比、根长密度和根体积比与覆盖度为60%的华扁穗草地、覆盖度为90%的金露梅+杂草地、裸露地3种试样的相比,平均高出 0.30、2.40、0.76 个百分点,较多的根系能够更有效地胶结土体,提高土体的抗蚀能力。由此表明,黄河源区河岸发育的草甸植被根系能够有效地固结土体,在受水流作用崩落水中后,崩解速率较慢,能够延长对近岸的保护时间,同时地下发达的植被根系在冻融过程中能够发挥较好的温度阻隔效应。
