基因组挖掘技术及其在真菌中的研究进展
2019-11-21徐杰黄建忠李力
徐杰 黄建忠 李力
(福建师范大学生命科学院,福州 350108)
与传统的以“模型导向”科研方式不同,基因组学研究以其高度的系统性,以数据为导向,开创了全新的研发方法。20世纪以来高通量基因测序技术的建立直接推动了生物信息学的发展[1],微生物基因组测序的成本不断降低,周期缩短,越来越多的生物遗传信息被公布,公共数据库积累了大量的基因组序列数据,后基因组时代来临[2]。在此基础上,人们通过生物信息学软件对不同基因结构进行识别鉴定、功能分析、相互关系和可能的表达调控机制,发现了大量未知的可能与合成新型次级代谢产物相关的基因簇,这为直接以基因为导向替代传统的以生物活性为导向,寻找潜在药物提供了可能[3]。
生物体次级代谢产物是抗生素的主要来源,其频繁的使用会加速细菌适应性进化而产生免疫能力。研究证实,在对某一药物产生抗性的同时,细菌对其结构类似物或衍生型药物的抗药性也会得到提高[4]。自 1979 年 Campbell[5]从 40000 份土壤样品发现新型抗生素阿维菌素后至今的40年内,尚未找到其它有重要应用价值的活性化合物,以生物活性为导向的旧有药物筛选方式陷入瓶颈。挖掘药物的关键在于避免已知化合物的过度复筛,能否发现新药物很大程度上取决于挖掘过程中在概率上的盲目程度。用科学理论指导开发方向,可降低成本和周期,避免已知化合物的重复研究。微生物中负责编码次级代谢产物合成的酶基因通常成簇存在,通过NCBI中基础局部比对搜索工具(BLAST)搜索相关酶类,并通过基因预测算法,如Augustus发现未知的生物合成基因簇,通过信息学工具可预测出产物结构以及可能含有的新药物作用靶点[6-8],这为新药开发提供了新方向。
次级代谢产物是一种具有多样化结构的小分子,它在微生物体内不是为了满足机体基本生长需要而产生,而是作为一种“武器”来对抗复杂的生态环境并保证在与其它微生物的竞争中处于优势地位。然而编码此类化合物的基因往往只在特定环境下才表达,常规培养条件下处于沉默状态或表达量低,难以分离纯化和投入工业生产。统计数据显示微生物中表达并分离出对应代谢产物的合成基因只占总数的10%左右,微生物资源在新药研发领域的潜力远远没有被开发出来,还需要不断地进行探索[9]。基因组挖掘是指通过高通量测序技术对生物信息学预测的可能编码高价值天然产物的基因进行测序,了解基因表达翻译的控制机制并通过一系列手段将其激活表达,分离纯化出相应目的产物后解析结构并进行生物活性测试的技术。与传统方法相比它将整个过程简化为编码次级代谢产物基因序列数据在培养基上的变现及分离柱上的纯化,让整个流程处于分子生物学的指导[10],研究主要包含以下方向。
1 基因组挖掘
1.1 传统基因组学挖掘
基因组学挖掘天然产物的关键在于寻找到有价值的合成基因,酶作为生物体内代谢活动的催化剂在次级代谢产物合成途径中发挥了重要作用,催化同一类型反应的酶系在分子结构上具有保守性,核心往往有一个高度重复的氨基酸序列,直接以负责目的代谢产物的合成酶作为靶点,系统进化树分析含有关联序列的酶基因,设计交联探针或PCR引物即可定向选择具有合成潜力的基因。最典型的就是目前研究比较深入的聚酮化合物合酶(Polyketidesynthase,PKS)和非核糖体肽合酶(Nonribsomal peptide synthetases,NRPS), 其 分子结构是由多个结构单元组成,线性排列的负责相应结构单元的模块化合成酶基因组成了完整的生物合成基因簇。Sun等[11]对海洋黏细菌Haliangium ochraceum SMP-2进行基因组挖掘,发现了一种聚酮化合物-非核糖体肽杂合化合物Haliamide 1(图1),通过核磁共振和HR-MS对结构进行表征和喂养实验发现其生物合成的结构单元为苯甲氨酸盐、丙氨酸和丙氨酸盐,从而建立了卤胺合成的生物模型,活性检测发现对Hela-S3细胞具有毒性。

图1 Haliamide 1结构
1.2 比较基因组学挖掘
比较基因组学是基于基因组图谱和测序技术,对已知基因和基因组结构进行比较,了解基因功能、表达机理、物种进化的科学。在研究目的蛋白基因的同时,关注完整的基因簇或基因邻域,分析不同分子间相关性来挖掘未知酶系和其体内的生物合成机制,促进新化合物的发现。将已知基因序列的功能基因簇通过NCBI数据库与其它已测序的生物遗传图谱进行局部共线性比对验证是否具有类似的合成路径,挖掘已知物种未发现的合成潜力。通过将编码目标修饰蛋白的抗药性基因与未表征次级代谢合成基因簇孤儿BGCs相关联,基因组定向挖掘的过程中,可预测路径特异性小分子的生物学功能。Tang等[12]对86个盐孢菌基因组的全基因组查询,获得与天然产物BGCs共定位的管家基因,筛选出一个可能具有脂肪酸合成酶抗性基因的PKS-NRPS杂合BGC(TLM),从而推测出硫代四酸类抗生素的合成途径。
1.3 宏基因组学
宏基因组是指环境中所有微生物遗传物质的总和,包含可培养和未可培养微生物遗传物质基因,通过构建宏基因组文库,绕过培养难点直接在基因层次研究未可培养微生物的合成潜力,可能会发现新的耐药基因和抗生素机制,完善药物靶点,以及鉴定在不同生长环境下对细菌、真菌生长起重要作用的基因,从而丰富基因库来源[13]。操作包括从环境中提取样品DNA,高通量测序后选择合适的载体拼接修饰,导入宿主细胞激活表达,实现对基因组网络的功能验证与开发利用。Levin等[14]通过结合基因组学和生物学知识,将合成具有保守氨基酸结构的蛋白质基因和正常人体的消化道菌群数据进行对比,分析了甘氨酰自由基酶(GRE)家族,以甘氨酸为中心用自由基进行化学反应,确定了一个新的在生产L-脯氨酸通路上起重要作用的反式 4 羟基脯氨酸脱水酶,它在所有肠道微生物宏基因组样本中都能发现,从而揭示了反式 4 羟基 L 脯氨酸的常规代谢途径。
2 沉默基因的表达
通过基因组挖掘的方法能够准确发现大量与合成次级代谢产物相关的基因簇,然而多数基因在常规培养条件下处于沉默状态,必须在特定生理环境下或外来细菌病毒入侵引起免疫应答机制时才会激活。这些基因由于常规条件下不表达,因而更有可能在人为控制下激活产生骨架新颖、生物活性高的次级代谢产物,如何选用适当的方法激活这些“沉默代谢途径”是继基因组挖掘后的重要研究内容,也是最终获得潜在药物资源的关键。现阶段激活沉默基因的策略如下。
2.1 改变培养条件、共培养和化学诱导
通过改善培养基组分、温度、pH、光照条件、气压、氧浓度、微量元素,或与合适病原菌、竞争菌共培养,提高生存压力刺激沉默基因表达。某些能够改变微生物表观遗传途径的小分子物质,如DNA甲基转移酶抑制剂、组蛋白去乙酰化抑制剂能够影响DNA甲基化、组蛋白修饰和RNA干扰等从而影响微生物对于次级代谢产物的合成调控,诱导沉默基因表达。Bode等[15]研究小组通过改变培养条件从6个微生物中分离出超过25个结构类型的100多个化合物;Scherlach等[16]发现Aspergillusnidulans这种产生在水稻培养基但不产生在其它介质中的新型喹诺林类生物碱;Shwab等[17]发现编码黑曲霉组蛋白去乙酰化酶(HDAC)的hdaA缺失,导致两个端粒近端基因簇的转录激活,随后相应的分子(毒素和抗生素)水平增加。
2.2 调控基因改造
次级代谢产物合成基因簇是复杂的,它不只是单纯含有合成化合物的基因,同时还包含大量参与基因表达,控制代谢途径的调节基因。通过设计方案对调节基因进行敲除、阻遏、或使其过表达,都会影响次级代谢产物的合成效率。选择性过表达正调控因子和降低负调控因子表达减少反馈抑制可促进基因的激活和代谢产物在发酵液中的积累[18]。Laureti等[19]过表达Streptomyces ambofaciens中的特异性调节基因Sam R0484激活了一个沉默的聚酮合酶的次级代谢基因簇表达,最终得到了新型的具有抗肿瘤活性的化合物Stambomycins。Molloy等[20]通过灭活推测的TetR转录抑制因子 arPRII等手段对编码化合物基因进行鉴定,质谱和核磁共振光谱对产物进行结构表征证实其是新的生物碱类化合物,生物活性测试发现对于黏菌的生长和菌落分化具有负面作用,其中一些抗菌活性较弱。
2.3 核糖体工程
与微生物在营养缺陷时的应激机制相似,核糖体工程就是通过以不同的微生物抗性突变为靶标,高效获得次级代谢产物合成能力得到提高的突变株的推理育种新方法。核糖体不仅是微生物体内蛋白质合成位点,还可以与药物结合抑制蛋白质合成从而调控次级代谢产物的合成途径,如链霉素和氯霉素可分别与核糖体30S亚基和核糖体50S亚基结合。在微生物的稳定生长期,与合成次级代谢产物相关基因的大量表达取决于此时的核糖体功能。以生物体中核糖体蛋白、RNA聚合酶RNAP、转录因子为作用靶点,通过在核糖体蛋白编码基因上引入定点修饰和突变,改造核糖体结构来改变相关催化酶的生物学性质,可能会激活相关沉默的抗生素合成基因,影响代谢途径[21-23]。如 Dong等[24]通过核糖体技术使海洋真菌杂色曲霉菌ZBY3获得耐新霉素抗性后发现6种新型抗肿瘤物质;Yi等[25]通过同样手段在紫草青霉G59中分离到一种新的环戊酮硫化物。
2.4 异源表达与强启动子替换
异源表达一般指异源蛋白表达,对于具有复杂调控机制或不可培养微生物的基因,将目的合成基因簇整合插入到质粒、黏粒、人工染色体等载体后在异源宿主内表达。该方法可绕过自身复杂的调控机制,减少原有产物含量积累造成的反馈抑制和对特定表达环境的需求,对生长缓慢的菌种可明显降低生产周期。目前用于挖掘新化合物的异源表达流程通常为:大肠杆菌扩增载体、酵母细胞对目的片段进行拼接、曲霉作为宿主细胞对目的基因进行表达[26]。启动子是位于起始位点上游能增加或降低基因转录速率,被RNA聚合酶识别、结合和开始转录的一段DNA序列,它含有RNA聚合酶特异性结合和转录起始所需的保守序列,本身不被转录。在沉默基因簇前添加或替换诱导型或组成型启动子,构建新的调控机制,通常可刺激基因激活提高表达量,利于最终工业生产,常见的有构巢曲霉A的持家启动子gpdA,黑曲霉作为宿主异源表达时的常用强启动子glaA等[27-30]。Wang等[31]发现在工程大肠杆菌中表达编码嗜热链球菌双功能谷胱甘肽合成酶基因GshF,分批发酵分离积累的GSH达到以往表达最高水平。黄开华等[32]利用蛹虫草自身强启动子,构建天蚕菌素抗菌肽过表达载体,介导农杆菌遗传转化系统,在蛹虫草中成功表达,活性测定和抑菌实验表明相较于野生型其生物学活性明显提高。Saha等[33]通过启动子工程和异源表达对海洋链霉菌SCSIO02999中ptm基因簇进行激活,发现6种新的多环四酸酯大环酰胺PTMS,其对人癌细胞株具有较强细胞毒活性,体内基因破坏实验和体外生化实验表明多循环形成一个还原环化级联,并证明PtmC是双功能环化酶,可催化伊卡毒素内五元环形成。
3 真菌基因组挖掘进展
来源于真菌的天然产物具有广泛的生物活性,然而其遗传调控途径相对细菌复杂,导致现阶段对其研究较少。随着高通量测序技术和生物信息学发展,对复杂微生物代谢通路进行分析的技术逐渐成熟,真菌作为寻找新颖骨架结构和独特作用机制药物的重要来源,其生物医药价值受到众多科学家的关注。真菌中次级代谢产物主要分为:萜类化合物、聚酮类化合物、生物碱、非核糖体肽、氨基酸衍生物、氨基酸/聚酮杂合体细胞松弛素等。目前已从真菌中获得的药物包括具有抑菌、抗肿瘤、抗炎和免疫调节功能的核苷酸类抗菌素虫草菌素;药用真菌灵芝中分离的抗氧化抗衰老、促进核酸蛋白质代谢的灵芝多糖以及最经典的青霉素(图2-A)和头孢菌素(图2-B)等,研究还发现红豆杉内生真菌会合成毒性较小的广谱抗肿瘤药物紫杉醇(图2-C)[34]。绝大多数真菌种类属于尚未认知状态,近几年对于真菌挖掘潜在药物又有了新进展[35]。
Shu等[36]从青霉Penicilin Herquei中分离到新的重氮杂环辛烷衍生物和新戊二酮衍生物,进行生物活性测试发现具有抗丙型肝炎病毒(Human antiheptitis C virus,HCV)能力,这是首次报道具有抗HCV能力的重氮环辛烷衍生物(3-A)。Chiba等[37]在筛选新的抗流感病毒时,从真菌FKI-7215发酵液中分离出草甘氨酸A(Herquline A)(图3-C),其对于流感病毒复制有剂量依赖性的抑制作用且对病毒神经氨酸酶没有抑制作用。Tansakul等[38]从土壤真菌黑曲霉(Penicillium herqueiPSURSPG93)中分离出新的苯丙烯酮衍生物和五个已知的phenalenone衍生物,其一个phnalenone衍生物对非洲绿猴肾成纤维细胞无细胞毒性,但抗氧化能力弱。Arunpanichlert等[39]在海草源真菌Pestalotiopsp中分离出4种新的代谢产物,包括2种巯基萜类、1种异香豆素和1种苯酚,提出了一种多烯类化合物的生物合成途径并有较好的抗菌、抗疟和细胞毒活性。Klaiklay等[40]从红树林真菌Pestalotitisp中分离到4种新的联苯醚、3种新的色酮、1种新的黄酮、1种新的花椒酮、1种新的丁烯内酯-Pestrolide(图3-D)和11种已知化合物。光谱技术确定了它们的结构。Pestalolide对白色念珠菌和新隐球菌具有较好的抗真菌活性。Uchoa等[41]通过改变培养条件,从巴西海岸筛选的一株黑曲霉中发现具有新的氮化骨架的天然呋喃衍生物(图3-B),其对HCT-116细胞系具有毒活性。随着研究的深入,真菌作为具有重大潜在价值的自然资源宝库,在研究新型药物和开发药物先导化合物过程中将发挥重要作用。
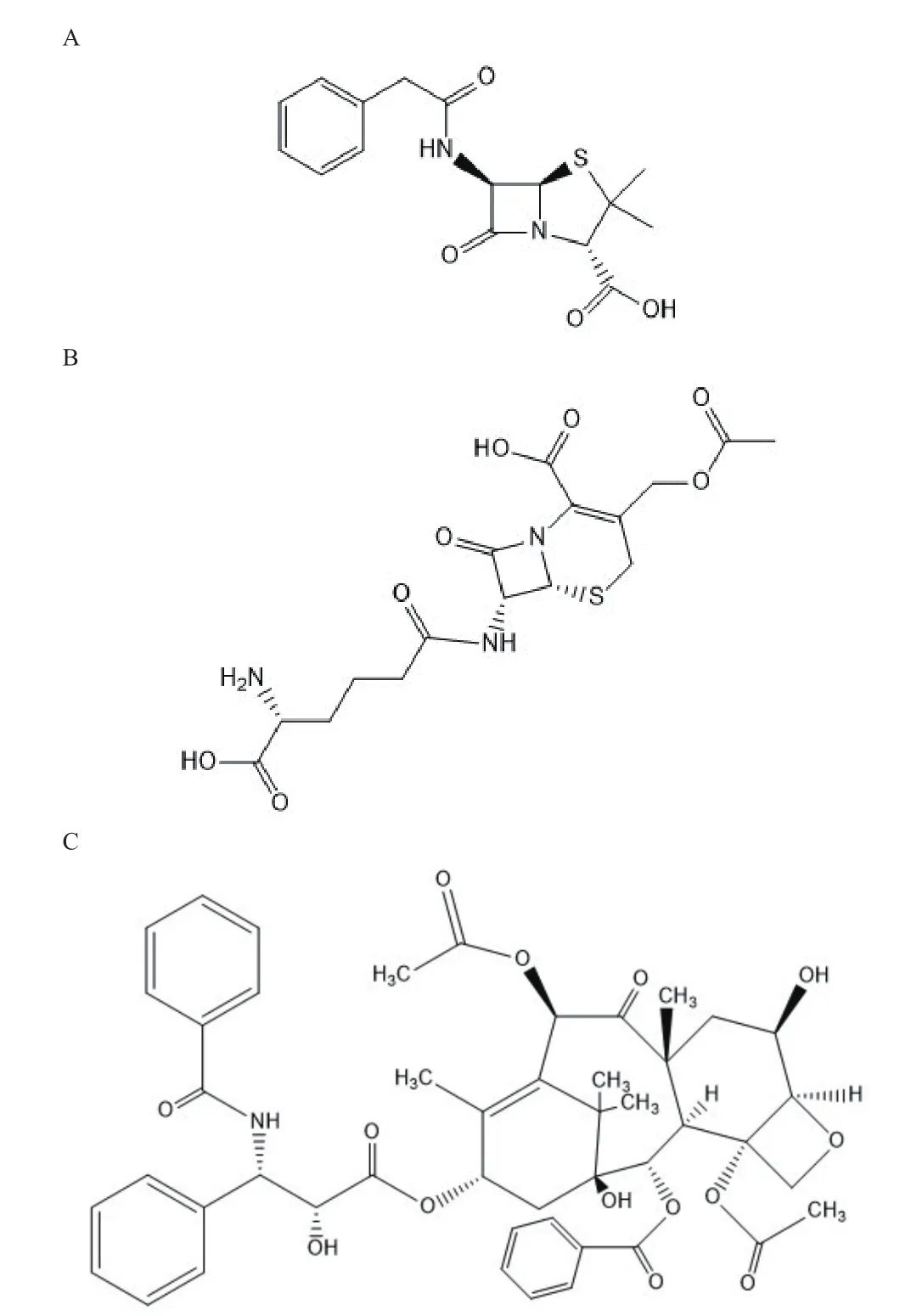
图2 青霉素(A)、头孢菌素C(B)、紫杉醇(C)结构
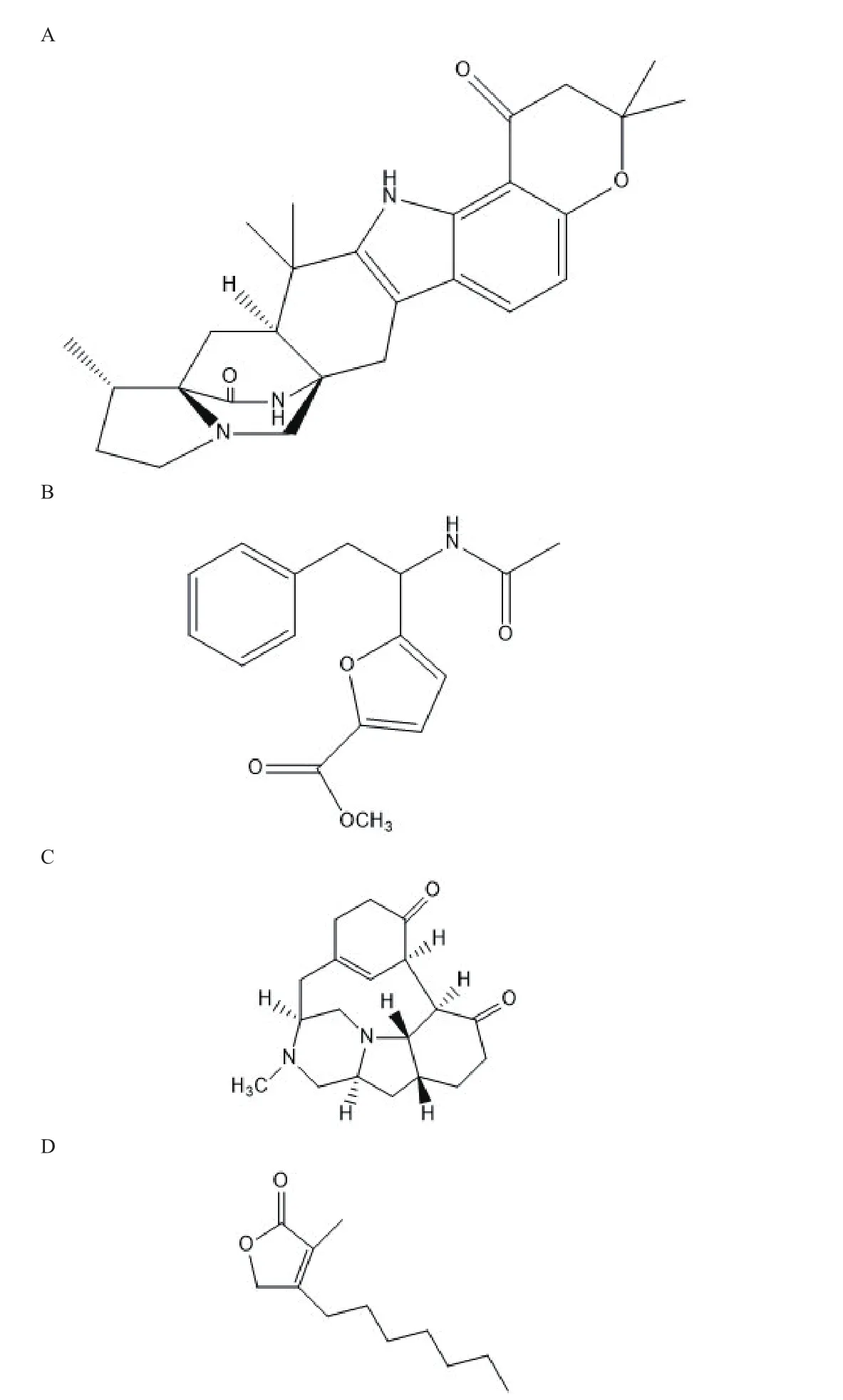
图 3 抗HCV(A)、抗HCT-116(B)、Herquline A(C)、Pestrolide(D)结构
4 展望
分子生物学技术的发展,使人们对于生物体遗传基因、代谢调控机制的了解愈加深刻,也越来越渴望通过基因水平定向寻找与实际需求相符的药物,尤其近几年计算机和数学等工具发展带来的强大基因图谱分析能力,更是为此提供强大的助力。随着大量微生物遗传图谱公布,潜在的合成基因簇数据不断积累,最终对于天然活性产物的挖掘将会得到井喷式发展,具有有效药用价值的化合物也会越来越多,真菌由于自身特性也必将扮演重要的角色。尽管现如今对于微生物资源利用才刚刚起步,尚没有完善的依托基因组挖掘技术开发药物的流程,但可以预计,通过诱导激活沉默基因簇充分挖掘生物合成潜力,将成为未来新药研发的重要手段,也会吸引更多国内外科研工作者的兴趣。
