籼型杂交水稻恢复系粒长及粒重的分子改良
2019-11-14苏相文高方远任鄄胜吕建群陆贤军任光俊
苏相文, 高方远, 任鄄胜, 吕建群, 陆贤军, 任光俊
(四川省农业科学院作物研究所, 四川 成都 610066)
【研究意义】目前,水稻育种的主要任务之一是在高产的基础上进一步提高稻米品质[1-2]。水稻品质性状包括外观品质、加工品质和蒸煮食味品质,其中外观品质是最直观的性状,决定消费者的购买欲望,细长型的稻米受到多数消费者喜爱[3]。水稻籽粒的增长有利于粒重的增加[4]。因此,增加水稻粒长和长宽比能同时改良水稻外观品质和产量性状。分子标记辅助选择(Molecular Marker-assisted Selection,MAS)可以大大提高育种效率、缩短育种周期,定向改良受体品种的目标性状,是现代分子育种的重要手段。利用MAS技术选择粒长基因,培育新的长粒品种,对水稻生产具有重要意义。【前人研究进展】近年来,随着分子生物学及生物信息学的发展,越来越多的水稻粒长基因被定位[5-9]。目前,已经被克隆的粒长基因有qGL3[10]和gs3[11]。Zhang等[10]指出qGL3编码一个含有2个Kelch功能域的蛋白磷酸酶OsPPKL1,在水稻粒长调控中发挥负调节子的作用。Fan等[11]的结果表明gs3是调控水稻粒长的主效QTL,其等位基因gs3增加粒长[11]。在MAS育种应用上,杨梯丰等[12]、李扬等[13]和谢坤[14]进行了报道,他们分别利用gs3基因功能性分子标记筛选出了新的长粒株系。【本研究切入点】有关水稻粒长改良的研究主要是以导入gs3基因为主,且报道较少。【拟解决的问题】以籼稻中间材料R855302作为长粒gs3基因的供体亲本,以农艺性状优良的抗稻瘟病、短粒恢复系R33947为受体亲本,利用gs3基因的特异分子标记开展MAS育种研究,以期育成长粒、千粒重较高的新恢复系,为进一步培育优质、高产兼顾的杂交水稻新品种奠定基础。
1 材料与方法
1.1 试验材料
受体亲本为R33947,其株叶型较优、稻瘟病抗性强,但籽粒短小、千粒重较低,配制的杂交组合产量优势不突出。供体亲本为长粒型中间材料R855302,其来源是川香29B/成恢3203//成恢3203///成恢3203。这两个材料均为本团队自主育成。
1.2 gs3基因内分子标记辅助选择及RICE6K育种芯片分析
采用CTAB 法[15]提取水稻幼嫩叶片总DNA。gs3基因内标记RGS1 (F:5’-TCCACCTGCAGA TTTCTTCC-3’;R:5’-GCTGGTCTTGCACATCTCTCT-3’)[16]由北京迪纳兴科生物科技有限公司合成。 PCR反应体系和反应程序参照李浩杰等(2004)方法[17]。采用RICE6K育种芯片(中国种子集团有限公司)进行遗传背景分析。该芯片包含5102个SNP标记和InDel标记,其中4500个标记能被稳定检出。这些标记分布在水稻12条染色体上,平均标记距离为1 Mb[18]。
1.3 目的基因与表型选择
用R33947与R855302杂交,并用R33947回交1次。BC1F1单株用分子标记鉴定gs3基因型,选择该位点为杂合的单株,收获自交种子。在BC1F2群体中选择长粒型单株。以后各世代结合分子标记辅助选择和表型观察,选择农艺性状优良、田间稻瘟病抗性强和低垩白的长粒型优良单株或株系。从BC1F5代开始,进行配合力测定,观察产量杂种优势。对中选株系进行考种和遗传背景分析,并与优质香稻不育系川康606测交,培育米质优良和产量配合力高的新恢复系。选育流程见图1。
1.4 田间种植和性状考察
1.4.1 田间种植 恢复系材料,每个小区种植4行,每行10株,单本插,间距16.7 cm×23.3 cm。记载生育期和整齐度。成熟时,结合分子标记鉴定结果和表型,选择优良单株或株系。在BC1F7世代,每个小区3次重复,各5行,每行10株,单本插,间距16.7 cm×23.3 cm,以R33947为对照。
测交组合,每个小区种植8行,每行10株,单本插,间距16.7 cm×26.7 cm,以F优498为对照。
1.4.2 性状考察 对改良的长粒型株系和短粒型受体恢复系R33947进行性状考察,包括:株高 (Plant height, PH)、单株有效穗数 (Number of panicles per plant, NP)、每穗实粒数 (Number of filled grains per panicle, NFG)、每穗总粒数 (Total number of spikelets per panicle, TNS)、结实率 (Seed setting ratio, SSR)、千粒重 (1000-grain weight, TGW)、单株产量 (Grain yield per plant, GY)、粒长 (Grain length, GL)、粒宽 (Grain width, GW)、长宽比 (Ratio of length and width, L/W)、垩白粒率 (Chalky grain rate, CGR)、垩白度 (Chalkiness degree, CD)。
对测交组合测定产量杂种优势。2018年川康优637(川康606A/成恢637)参加长江上游科企联合体区域试验,有关试验方案和数据的收集按照区试要求进行。
1.5 数据收集和分析
考种数据的收集和整理使用WPS表格,数据分析使用SPSS 19.0。
2 结果与分析
2.1 长粒型改良系的培育
2012年冬季,海南种植R33947与R855302的BC1F1群体,在苗期用gs3基因的功能标记RGS1进行分析,共有21个单株为杂合基因型。2013年夏季,在成都种植21个BC1F2群体,田间收获籽粒较长、株型较好的单株,并调查各单株的垩白粒率和垩白度,共保留72个低垩白单株。从BC1F3代起,结合分子标记辅助选择、田间稻瘟病抗性鉴定和农艺性状观察,选择长粒型优良单株;至BC1F5代,中选99个株系。在配合力测定的基础上,于2015年获得4个性状稳定的长粒型优良株系,分别命名为RGL1、RGL2、RGL3和RGL4,其gs3基因型与长粒供体亲本R855302相同(图2),籽粒长度显著大于短粒受体亲本R33947(图3)。

图1 长粒型恢复系成恢637的选育流程Fig.1 Breeding procedure of the restorer line Chenghui 637 with long grain
2.2 改良系与亲本的籽粒和垩白性状比较
4个改良系的粒长为10.15~10.39 mm,平均比短粒亲本R33947增加粒长25.61 %,达极显著水平,与供体亲本R855302差异不显著;粒宽为2.69~2.80 mm,显著小于亲本R333947的3.08 mm;长宽比为3.81~3.86,平均比受体亲本R33947增加43.89 %,达极显著水平;4个株系之间在粒宽、粒长和长宽比性状上的差异不显著(表1)。

“M”为DL2000,“1”和“6”为短粒对照R33947,“7”为长粒对照R855302, “2”、“3”、“4”、“5”为改良系RGL1、RGL2、RGL3和RGL4‘M’ is DL2000; ‘1’ and ‘6’ are R33947 with shorter grain; ‘7’ is R855302 with longer grain; ‘2’, ‘3’,‘4’ and ’5’ indicated the improved lines named as RGL1, RGL2, RGL3 and RGL4, respectively图2 利用gs3基因内标记RGS1对4个改良系和亲本的PCR扩增结果Fig.2 PCR analysis of gs3 with marker RGS1 in four improved lines and the parental lines
4个改良系的垩白粒率极显著低于供体亲本R855302。RGL1、RGL3和RGL4的垩白粒率较低,与受体亲本R33947没有显著差异。其中,RGL4的垩白粒率较低,为1.63 %。在垩白度方面,4个改良系极显著低于供体亲本R855302,然而高于受体亲本R33947。其中,RGL4的垩白度较低,与受体亲本R33947的差异不显著;另外3个改良系的垩白度有所增加,与受体亲本R33947的差异达显著水平,其中RGL3较高,达1.58,且与RGL1和RGL2的差异达显著水平(表1)。
2.3 改良系与亲本的主要产量性状比较
经多重比较(表2)发现,RGL1、RGL2和RGL3的株高较高,它们与RGL4和双亲的差异达显著水平。RGL1的单株有效穗数较多,RGL4个改良系的单株有效穗数较少,它们之间的差异大极显著水平,且与其他2个改良系和双亲有显著差异。4个改良系的千粒重为26.05~27.04 g,平均比比供体亲本R855302低12.56 %,但是比短粒亲本R33947增加26.27 %,差异达极显著水平。 在每穗总粒数和每穗实粒数方面,4个改良系都显著低于受体亲本R33947,显著高于供体亲本R855302,且它们之间有显著差异。RGL1的每穗总粒数和每穗实粒数较低,且与其它3个改良系的差异达极显著水平。4个改良系的结实率都显著高于受体亲本R33947,显著低于供体亲本R855302,且改良系之间的差异也达到显著水平。从单株产量来看,RGL1、RGL2和RGL3的单株产量显著高于受体亲本R33947,其中RGL2的最高,达34.27 g,比短粒亲本R33947增产30.58 %;RGL3为29.79 g,比对照增产14.44 %;RGL4的单株产量最低,为24.45 g,减产6.67 %。
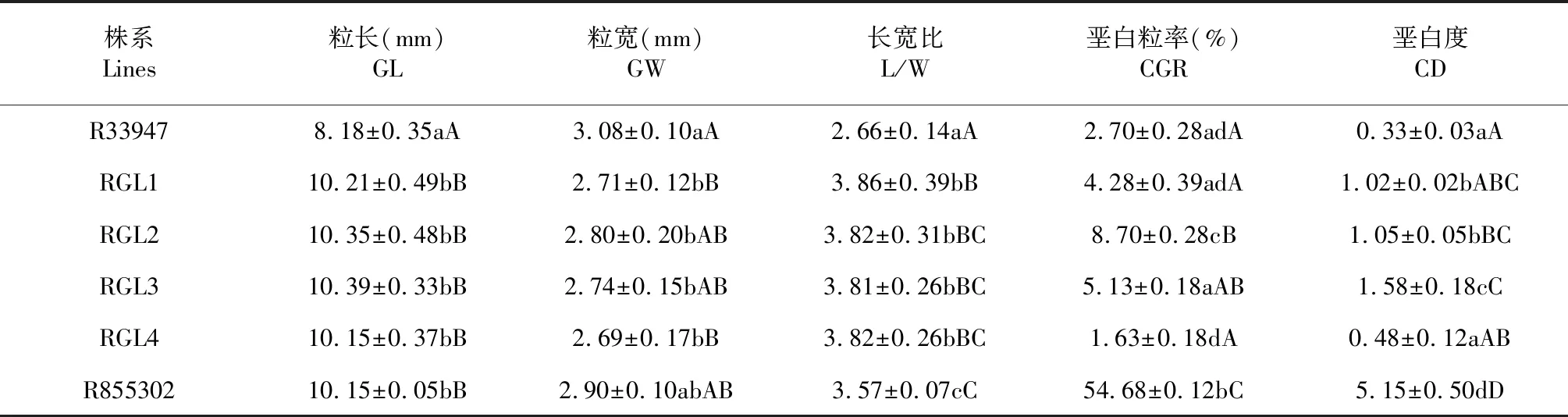
表1 改良系和亲本的粒形和垩白性状
注:同列不同小写字母表示差异显著(P<0.05) ;同列不同大写字母表示差异极显著(P<0.01) 。下同。
Note: Different lower case letters in each column indicated significant differences (P<0.05). Different capital letters indicated extremely significant differences (P<0.01).The same as below.
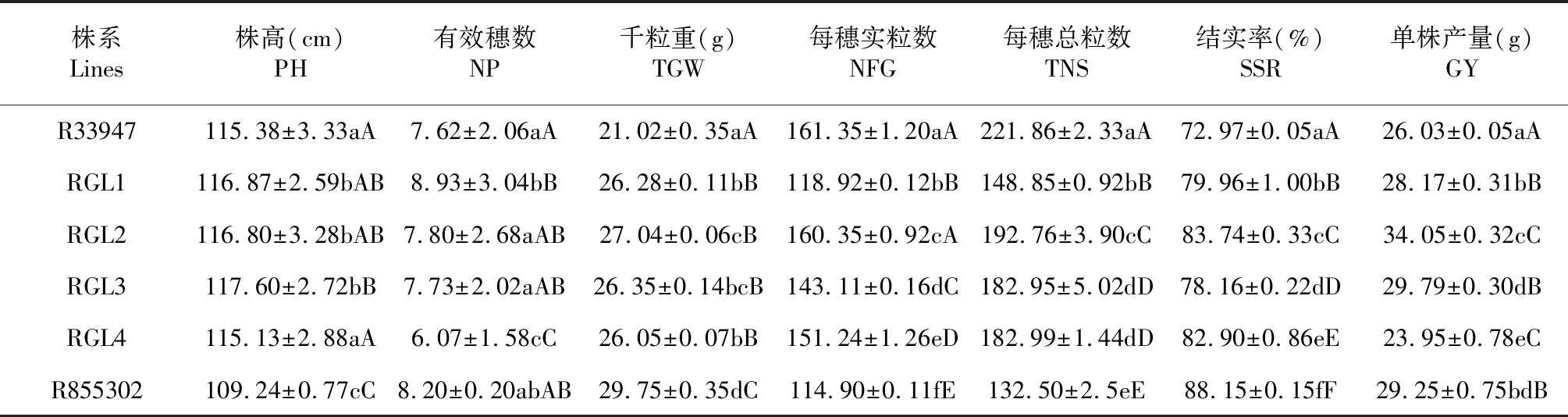
表2 改良系和亲本的主要产量性状表现
2.4 长粒改良系的育种芯片分析结果
中国种子集团有限公司生命科学技术中心提供的RICE6K育种芯片数据显示,4个改良系的分析结果相同,它们之间没有差异位点。这可能是因为在BC1F5世代来自于同一个株系。育种芯片共检测到4402个SNP位点,其中与短粒亲本R33947带型一致的标记数是3704个,占检测位点数的84.14 %。改良系有695个位点来自于与长粒亲本R855302。

图3 改良系与亲本的粒型比较Fig.3 Comparison of grain type among the improved lines and parents
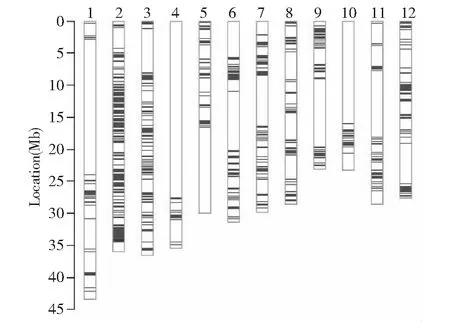
线条表示SNP位点为纯合供体亲本R855302片段,白色区域表示SNP位点为纯合受体亲本R33947片段The lines represents the SNP loci with homozygous genotypes as the donor parent R855302, the white boxes indicate the loci with the same genotypes as the recurrent parent R33947图4 长粒型改良系育种芯片分析Fig.4 Analysis of four long-grain improved lines using RICE6K array
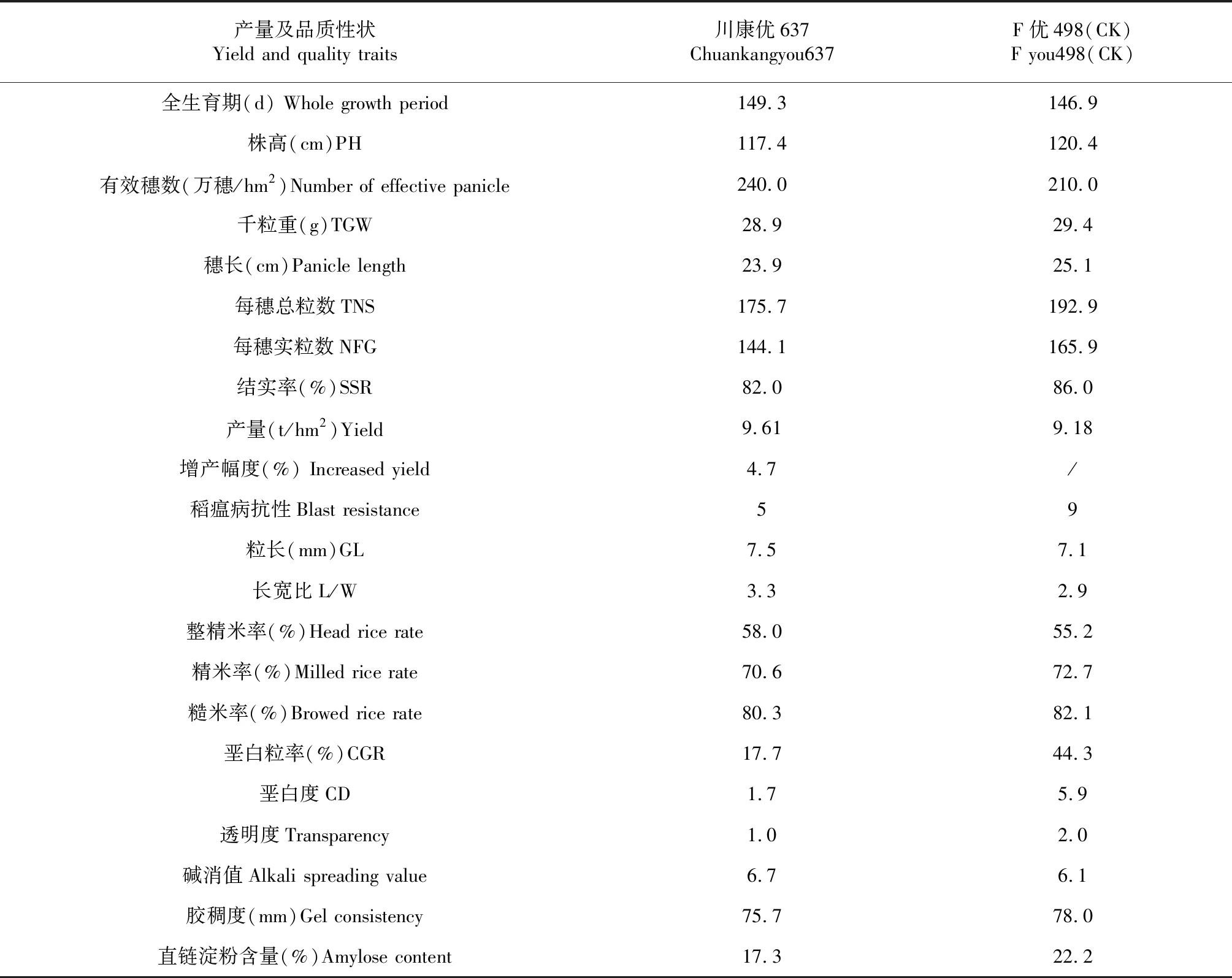
表3 川康优637和F优498的产量及品质性状比较
它们主要集中在第2、3、6、7、8和12染色体上(图4)。
2.5 优良杂交组合川康优637的产量及品质测试
2016年成都,根据田间观察,RGL1和RGL3与优质香稻不育系川康606A的杂交组合表现较强的杂种优势。测产分析表明,杂交组合川康606A/RGL3的产量较高(数据略)。随后将长粒型株系RGL3定名为成恢637,并将该组合定名为川康优637。2017年,川康优637参加多点试验,表现优良。2018年参加长江上游科企联合体中籼迟熟组区域试验。结果表明,川康优637的全生育期比对照F优498长0.9 d,株高矮4.3 cm;有效穗数达240万穗/hm2,比对照高14.29 %。川康优637的产量达到9.61 t/hm2,比对照增产4.7 %(表3)。
川康优637米质优良。在外观品质性状方面,川康优637的粒长和长宽比分别为7.5 mm和3.3,分别比对照F优498增加5.63 %和13.79 %;垩白粒率和垩白度分别为17.7 %和1.7,比对照降低150.28 %和247.06 %;川康优637的透明度为1.0,比对照提高100 %。在加工品质性状方面,川康优637的糙米率和精米率分别是80.3 %和70.6 %,略低于对照F优498,但是整精米率比对照增加5.07 %。在食味品质性状方面,川康优637的碱消值为6.7,比对照高9.84 %;胶稠度略短于对照,直链淀粉含量为17.3 %,比对照低28.32 %。与部颁优质米标准对比,川康优637达到“优 2”等级。此外,川康优637在四川和贵州点穗颈瘟最高级为5级,而对照F优498为9级、重感稻瘟病。
综上所述,川康优637是一个达到部颁2级优质米、丰产性好、抗病性强的杂交水稻新品种。
3 讨 论
随着人们生活的改善,消费者对籼稻米质有了更高的要求。主要需求是稻米细长、透明度好、口感软硬适中,香气浓郁等。由于历史上粮食短缺的原因,我国水稻育种长期重视丰产性改良,而对优质育种关注不够。至今,杂交水稻给消费者的印象是产量很高,但品质一般。随着水稻分子技术的进步,水稻粒长、粒宽和香味等品质性状基因已先后被克隆[11, 20-22]。不少研究者利用功能标记,通过分子辅助选择方法改良水稻的品质性状,并获得很好的效果[12-14]。在粒长改良方面,主要利用粒长主效基因gs3。2010年,杨梯丰等[12]首次实现了利用gs3对华粳籼74谷粒长性状进行定向改良,获得了一批谷粒长达到 9.51~9.67 mm的长粒形品系。2015年,谢坤利用gs3和qSS7定向改良Ⅱ-32B的粒长,获得了21个粒长在9.11~10.87 mm的长粒株系[14]。本研究利用来自中间材料R855302的gs3基因对谷粒长性状进行定向改良,结合低垩白、优良株型和稻瘟病抗性选择,育成了4个优良的长粒株系。这些研究结果表明,gs3在不同遗传背景中对粒长的基因效应表现稳定,适用于分子设计育种。
Fan C.C.等(2006)克隆了gs3基因,并证明其对粒长是主效基因,同时对粒重有影响[11]。谢坤的研究指出,当长粒gs3和宽粒GW28组合时,稻谷的千粒重较高[14]。王建等(2017)分析了粒型基因GW2和gs3在不同背景下对粒重的影响。结果表明,不同遗传背景中GW2和gs3的基因效应有差异,但同一背景下GW2对籽粒重量的贡献大于gs3[23]。在本研究中,4个改良株系的粒长和千粒重都显著优于短粒亲本R33947。这说明gs3能同时改良粒长和粒重。由于穗数、结实率等性状受其它基因调控,加之本研究只用短粒亲本回交了一次,所以尽管粒重都显著增加,但RGL4株系的单株产量仍然低于短粒亲本R33947。这一结果说明,在自交后代中既要关注目标性状的改良,又要注重综合性状的选择,才能进一步提高水稻单株和群体的产量。
4 结 论
通过常规的回交选育、分子标记辅助选择和抗病鉴定,结合优良形态选择的方法,筛选获得4份gs3纯合的优质抗病改良系,为选育新的优质水稻恢复系提供种质资源。育成优良恢复系成恢637,其与优质香稻不育系川康606A配制的新组合“川康优637”具有品质优良、丰产性好、抗病性强等特点。
