海南黎母山坡垒种群结构及伴生群落特征研究
2019-11-12符明期方燕山桂慧颖方发之
符明期,方燕山,桂慧颖,方发之
(1.海南省黎母山林场,海南琼中 572934;2.海南省林业科学研究所,海南海口 571100)
坡垒(Hopea hainanensis)为龙脑香科坡垒属植物[1],是热带沟谷雨林代表树种和特有种,原产于海南,越南北部也有分布。坡垒是中国珍贵用材树种之一,其木材优良,质地硬重,耐水浸渍,耐日晒,经久耐用,为中国特类木材[2]。正因其优质材性,被过度采伐加工,导致其种群数量不断减少,已经从热带雨林的代表树种转变为偶见种。目前坡垒在中国仅少量自然分布于海南中部及西南山区的原始林和次生林中,已处于濒危状态,被列为国家Ⅰ级保护植物,IUCN评估定为“濒危灭绝”等级[3],应立即采取有效措施进行保护。
种群结构是种群的基本特征,决定种群的动态变化和发展方向,而群落特征则代表着物种的对某一生境的需求及其生态关系[4]。目前国内对坡垒的研究主要集中在其地理分布格局[5]、幼苗分布与母树关系[6]、苗木分级[7]、引种实验[8]和扦插育苗[9]等方面,关于坡垒种群生态学的研究鲜有报道。该文采用样方调查法,研究坡垒野外实生种群及其伴生群落特征,了解坡垒种群动态和生存现状,为构建坡垒适宜生境及人工促进种群恢复提供基础资料,并探索导致坡垒濒危的可能原因,提出相应保护策略。
1 材料与方法
1.1 研究区概况
黎母山自然保护区(19°07′22″~19°14′03″N,109°39′05″~109°48′31″E) 位于琼中、白沙两县境内,是中国珍稀原始雨林热带保护区之一。其主峰黎母岭海拔1411.7m,一般山峰海拔600~1000m,低山海拔300~600m。属热带季风气候,终年温暖湿润,雨量充沛。区内年平均气温23.1℃,年平均降雨量为2343.1mm,年平均相对湿度84%,年日照时数为1870.6h,年辐射总量109.37 kcal/cm2,平均年蒸发量为1617.8mm[10]。土壤类型主要为砖红壤、赤红壤和山地黄壤。砖红壤主要分布在保护区的东北部及南部边缘400m以下的丘陵地带,赤红壤主要分布于保护区海拔400~750m的低山,保护区海拔750m以上的山地主要是山地黄壤[11]。
1.2 调查方法
根据黎母山坡垒分布的现状,对黎母山保护区沿着一定的海拔梯度和不同管护点或监测点,对坡垒物种进行样方调查。再选择立地条件基本一致,且具有代表性的坡垒林地进行样方设置(20m×20m),共设置4个样地,四个样地分别代表了四个不同的伴生群落,样地1为白颜树+割舌树+东方肖榄群落(群落1);样地2为血桐树+公孙锥+鸭脚木群落(群落2);样地3为贡甲+青梅+景烈樟群落(群落3);样地4为鸭脚木+赤楠蒲桃+白颜树群落(群落4),具体情况如表1。分别记录各样地的海拔、坡位、坡向等生境指标,将每个样地再划分成5m×5 m的小样方,对小样方内所有乔木、灌木、藤本,草本进行调查。其中乔木、灌木调查的内容是物种、胸径、高度、冠幅;藤本的调查内容是物种,胸径;草本的调查内容是物种、株数、高度、盖度。
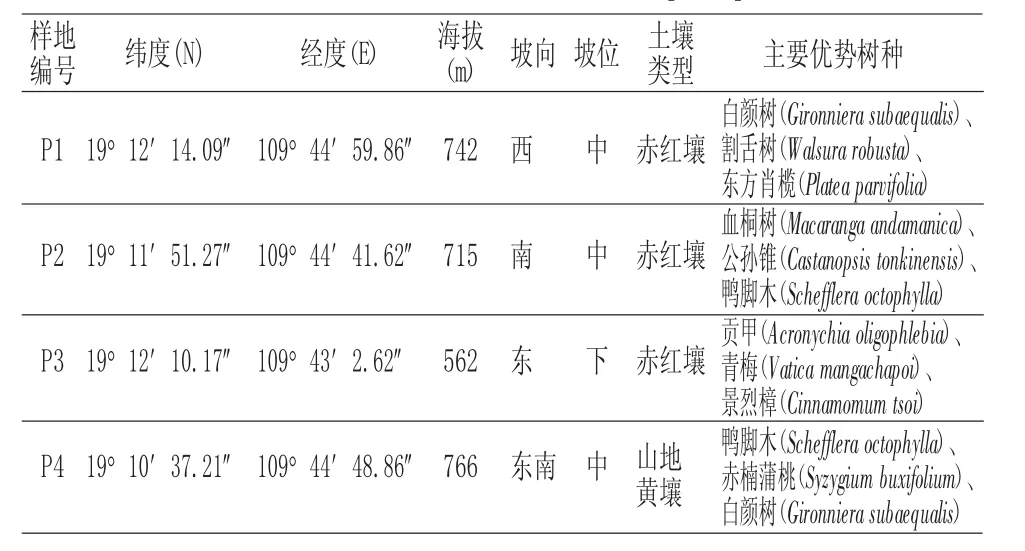
表1 样地基本情况Tab.1 Basic situation of Sample plot
1.3 数据计算
各生态学数据统计计算参照薛建辉[12]的《森林生态学》相关内容,具体如下:
(1)种群密度:
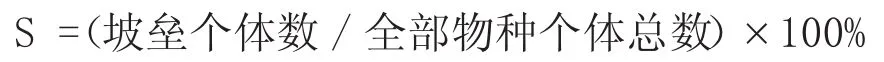
(2)重要值:

(3)Simpson指数:
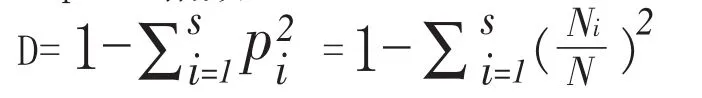
(4)Shannon-wiener指数:
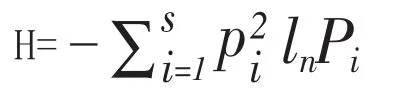
(5)Pielou指数:

式中Pi为第i个物种所占的比例,S为物种总数,Ni为第i个物种的个体数目,N为群落中所有种的个体总数,H’为群落的实测多样性,H’max为物种S的完全均匀群落多样性。
2 结果与分析
2.1 坡垒种群数量及结构特征
在试验设置的总面积为1600m2的调查样地中,共发现坡垒12株,种群密度为1.92%。树高最小仅2.5m,最大达28m,平均树高9.7m。胸径最小为1.8cm,最大为 49cm,平均胸径为 13.3cm。群落 1、群落2及群落3各发现坡垒1株,群落4发现9株。
种群结构对群落结构具有直接影响,还能客观地体现出群落的发展趋势。由于在实际工作中,个体年龄常难于确定,所以一般采用空间代替时间的方法,即以胸径级代替年龄进行分析。该文也采用此方法,参照杨小波等[13]对无翼坡垒种群的划分标准,将所有坡垒按胸径级划分结果如表2。黎母山坡垒种群径级结构呈现不连续的逆J型,小径级Ⅰ、Ⅱ占全部个体数的75%,成熟的大径级较少,从种群发展趋势看为增长型,呈上升趋势。

表2 黎母山坡垒种群年龄结构分析Tab.2 Population age structure of Hopea hainanensis in Limu Mountain
2.2 坡垒伴生群落主要种重要值以及坡垒在群落中的地位
四个样地内共发现各类植物131种,隶属于53科91属。其中乔木层共26科41属55种,灌木层共32科51属64种,藤草本层11科12属12种。
群落1共有各类植物27科36属42种。乔木层主要以东方肖榄(Platea parvifolia)、鸭脚木(Scheffleraoctophylla)及罗浮柿(Diospyros momrrisiana)、白颜树(Gironniera subaequalis)、割舌树(Walsura robusta)、大萼木姜(Litsea baviensis)等的一些植物构成;灌木层以海南暗罗(Polyalthia laui)、药用狗牙花 (Ervatamia officinalis)、贡甲(Acronychia oligophlebia)、厚边木犀(Osmanthus marginatus) 等构成;草本层以海南蒟(Piper hainanense)、单叶新月蕨(Pronephrium simplex)、铁线蕨(Adiantum capillus-veneris)等构成。群落1主要科为樟科5属6种,其次为壳斗科2属4种。主要种有白颜树(Gironniera subaequalis)、割舌树(Walsura robusta)、东方肖榄(Platea parvifolia)和大萼木姜(Litsea baviensis),重要值分别为 9.52、7.95、7.41 和 5.98。
群落2共有各类植物30科42属50种。乔木层主要以青梅 (Vatica mangachapoi)、公孙锥(Castanopsis tonkinensis)、红果樫木(Dysoxylum binectariferum)、白颜树(Gironniera subaequalis)等的一些植物构成;灌木层以药用狗牙花(Ervatamia officinalis)、罗伞(Brassaiopsis glomerulata)、山小橘(Glycosmispentaphylla)、柏拉木(Blastus cochinchinensis)等构成;草本层以双盖蕨(Diplaziumdonianum)、藤竹 (Panicum incomtum)、鱼尾葵(Caryota ochlandra)等构成。群落2主要科为樟科5属6种,其次为壳斗科2属4种。主要种为血桐树(Macarangaandamanica)、公孙锥(Castanopsis tonkinensis)、鸭脚木(Schefflera octophylla)和药用狗牙花(Ervatamia officinalis),重要值分别为7.71、7.58、7.35 和 7.34。
群落3共有各类植物25科40属47种。乔木层主要以青梅 (Vatica mangachapoi)、景烈樟(Cinnamomum tsoi)、蝶斗青冈(Cyclobalanopsis disciformis)和贡甲(Acronychia oligophlebia)等的一些植物构成;灌木层以粗毛野桐(Mallotus hookerianus)、毛荔枝(Nephelium topengii)、海南破布叶(Microcos chungii)等构成;草本层以穗花轴榈(Licuala fordiana)、单叶新月蕨(Pronephrium simplex)、双盖蕨(Diplazium donianum)、卷柏(Selaginella tamariscina)等构成。群落3主要科为樟科7属11种,其次为桃金娘科1属4种。主要种为贡 甲 (Acronychiaoligophlebia)、青梅(Vatica mangachapoi)、景烈樟(Cinnamomum tsoi)和粗毛野桐(Mallotus hookerianus),重要值分别为15.96、9.61、4.13 和 3.94。
群落4植物种类最为丰富,共有各类植物26科45属60种。乔木层主要以鸭脚木(Schefflera octophylla)、白颜树(Gironniera subaequalis)、托盘青冈(Cyclobalanopsis patelliformis)和黄杞(Engelhardia roxburghiana)等的一些植物构成;灌木层以粗毛野桐(Mallotus hookerianus)、假鹊肾(Streblus indicus)、双瓣木犀(Osmanthus didymopetalus)、毛荔枝(Nephelium topengii)等构成;草本层以穗花轴榈(Licuala fordiana)、藤槐(Bowringia callicarpa)、双盖蕨 (Diplaziumdonianum)、卷柏(Selaginella tamariscina)等构成。群落4主要科为樟科7属10种,其次为桑科3属5种。主要种为鸭脚木(Scheffleraoctophylla)、赤楠蒲桃(Syzygium buxifolium)、白颜树(Gironniera subaequalis)和厚壳桂(Cryptocarya chinensis),重要值分别为 7.15、6.99、6.91 和 6.11。
坡垒在群落1中重要值为5.23,排名第五位;群落2中重要值为2.84,排名第12位;群落3中重要值为1.96,排名第13位;群落4中重要值为2.81,排名第12位。重要值大小整体表现为:群落1>群落2>群落4>群落3。坡垒幼苗较为集中的群落4重要值较低,表明坡垒幼苗在郁闭度较高的热带雨林林下生长不占优势。

表3 主要植物种类及重要值Tab.3 Major plant species and their important values
2.3 坡垒伴生群落生物多样性
四个群落的多样性指数如表4所列。由表4可以看出,各个群落的Simpson多样性指数数值相近,整体表现为群落4最大,群落1次之,群落2最小;而Shannon-wiener指数则呈现完全不同的变化,群落1Shannon-wiener指数最大,显著大于其他三个群落,其余群落表现为:群落4>群落3>群落2;Pielou均匀度指数在群落间的变化差异与Shannon-wiener指数相似,Pielou均匀度指数大小在四个群落中整体表现为:群落1>群落4>群落3>群落2。从三个多样性指数综合分析可发现,群落1和4的各项多样性指数均大于群落2和3。结合坡垒在各个群落中的分布数来看,多样性指数大的群落坡垒分布较多,多样性指数小的群落坡垒分布少。

表4 各群落多样性指数Tab.4 Diversity index of each community
3 结论与讨论
3.1 结论
(1)从径级结构看,坡垒种群的发展呈上升趋势,但其种群密度低,导致其种群的增长缓慢且不稳定,易受外界胁迫影响。
(2)4个坡垒伴生群落内共发现维管束植物131种,隶属于53科91属。樟科、壳斗科等为优势科,白颜树(Gironniera subaequalis)、血桐树(Macaranga andamanica)等在伴生群落中具有较大重要值。坡垒幼苗较为集中的群落4重要值较低,在郁闭度较高的热带雨林林下生长不占优势。
(3)多样性指数大的群落坡垒分布较多,多样性指数小的群落坡垒分布少。
(4)低海拔区受人为干扰严重,高海拔区生长较为缓慢,林下自然更新困难,是导致坡垒濒危的重要原因。
3.2 讨论
种群结构是种群的基本特征,既可反映种群内不同大小个体数量的分布和配置状况,也能反映种群的数量动态及其未来发展与演变趋势[14]。该次调查发现黎母山坡垒种群径级结构呈现不连续的逆J型,小径级Ⅰ、Ⅱ占到全部个体数的75%,成熟的大径级较少,从种群发展趋势看为增长型,呈上升趋势。结合种群密度分析,每100m2内约有2株坡垒幼树生长,易受到外界不定因素影响,因此整体种群虽有上升趋势,但增长缓慢且不稳定。相关研究也表明低密度种相对于群落的主要种来说,更容易受到各种胁迫的影响而在群落中绝灭,从而使群落的物种多样性降低[15]。
植物群落是在一定地理区域内不同植物种群在相同环境下形成的组合体,对群落物种组成进行分析,是认识群落区系特征和生物多样性的基础[16]。该次调查结果表明,4个坡垒伴生群落内共发现维管束植物131种,隶属于53科91属。其中乔木层共26科41属55种;灌木层共32科51属64种;藤草本层仅11科12属12种。就种属丰富度而言,以樟科(8属19种)、壳斗科(3属9种)、茜草科(6属 7种)和桑科(3属7种)为优势科。从种水平来看,白颜树(Gironniera subaequalis)、血桐树(Macaranga andamanica)、贡甲(Acronychia oligophlebia)、鸭脚木(Schefflera octophylla)等占据群落主要优势,具有较大重要值。但坡垒幼苗较为集中的群落4重要值较低,表明坡垒幼苗在郁闭度较高的热带雨林林下生长不占优势。这与坡垒幼苗自身特性有关,研究发现坡垒幼苗虽为耐阴性,但随着苗龄的增加需光性逐渐增强[17],郁闭度高的热带雨林为坡垒幼苗提供光的能力有限,且坡垒属于慢生树种,因此未能在整个群落中占据重要生态位。
生物多样性是指地球上的生物(动物、植物和微生物)在所有形式、层次和联合体中生命的多样化,包括遗传多样性、物种多样性,、生态系统多样性和景观多样性[18]。生物多样性对维持生境稳定,保持土壤肥力,保证水质,调节小气候等有重要意义。对坡垒伴生群落生物多样性的分析也证实这一结论,调查发现,多样性指数大的群落坡垒分布较多,多样性指数小的群落坡垒分布少。
此外调查中还发现,在低海拔区受人为干扰严重,坡垒自然分布较少,高海拔区虽有幼树分布,但生长较为缓慢,林下自然更新困难,是导致坡垒濒危的重要原因。因此建议及时采取针对性保护策略以保存坡垒珍稀种质资源,具体方法如下:(1)就地保护,在坡垒分布区内实施全封闭封育管理,减少人为干扰,使坡垒群落自行演替;(2)迁地保护,采集坡垒果实与种子,选择气候适宜的区域建立种质资源圃,扩大其居群规模;(3)野外回归,通过迁地保护繁殖的坡垒幼苗,在类似生境的区域开展回归种植,建立种质资源保护区,开展生物特征特性、生长状况监测,丰富坡垒资源的野外居群规模;(4)加强基础研究及开发利用,加大科研投入,对坡垒生物特性、生态特性、繁育技术、遗传特性、育种潜力、开发价值等方面进行研究,为坡垒种质资源保护与利用提供技术支持。
