基于线粒体基因nd5对长江上游黑尾近红鲌遗传多样性分析
2019-11-02严太明王雄延张松培何佳洋
严太明,王雄延,罗 杰,李 松,张 骞,何 亮,张松培,何佳洋,何 智
(四川农业大学动物科技学院,四川 成都 611130)
黑尾近红鲌(Ancherythroculter nigrocauda),隶属于鲤科(Cyprinidae)、鲌亚科(Cultrinae)、近红鲌属(Ancherythroculter),是长江上游特有鱼类,主要分布于四川和重庆境内的长江干流及其支流[1-3]。近年来,由于过度捕捞、水利水电工程修建、水体污染和其他人为因素的影响,黑尾近红鲌种群数量急剧下降[4-6]。基于灭绝风险指数(ERI)和优先保护程度分析发现,急需对长江上游黑尾近红鲌采取保护措施[7-8]。
遗传多样性研究结果能为制定科学有效的物种保护策略提供重要信息[9-10]。基于线粒体基因cytb,Liu H.Z.等[5,11]发现龙溪河、习水河和木洞河黑尾近红鲌群体遗传多样性水平较高,而其遗传多样性水平有明显的下降。人工大坝的修建改变了支流群体与干流群体间的基因交流方式,栖息地环境的破坏、过度捕捞等导致的种群数量快速下降,可能是物种遗传多样性在短期内显著降低的主要原因[12]。因此,为评价既有保护措施的有效性和实施有针对性的保护策略,有必要对黑尾近红鲌的遗传资源现状进行再评估。
本研究采集了长江干流合江段、支流龙溪河和濑溪河野生黑尾近红鲌样本,对其线粒体NADH脱氢氧化酶亚基5(NADH dehydrogenase subunit 5,nd5)序列的扩增和分析,评估了其遗传多样性和遗传结构现状,旨在明确支流和干流种群以及支流种群间是否存在遗传分化,以期为黑尾近红鲌野生资源的保护和合理开发利用提供科学依据。
1 材料与方法
1.1 样品采集
89 个野生黑尾近红鲌样本于2016年7月—2017年11月分别采自长江干流合江段(HJ),长江一级支流龙溪河(LOR)以及沱江(长江的一级支流)一级支流濑溪河(LAR)(图1),采集地信息见表1。对其进行形态学鉴定后,取胸鳍保存于无水乙醇中,备用。
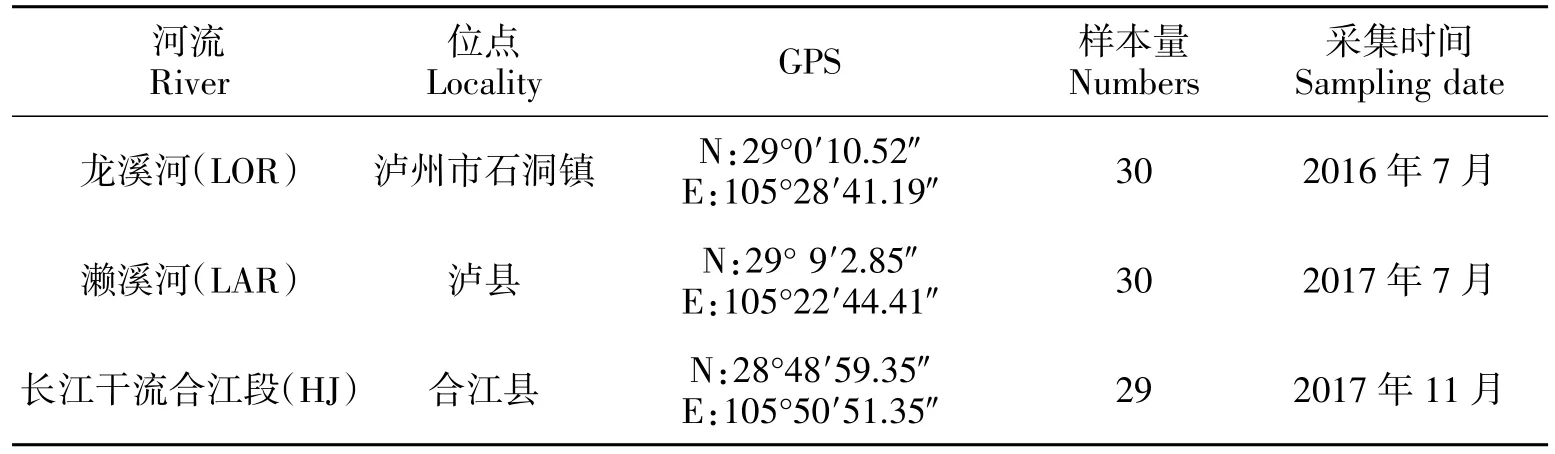
表1 黑尾近红样本基本信息Table 1 Sample information of A.nigrocauda populations
1.2 DNA提取
取胸鳍组织约100 mg,加入600 μL 组织裂解液(0.01 M Tris-HCl,pH 8.0;0.1 M EDTA;5 g/L SDS)匀浆后加入 10 μL 蛋白酶 K(10 mg/mL),56 ℃消化5 h 后,酚-氯仿法抽提基因组 DNA[13]。用 30 μL 1×TE Buffer 溶解乙醇沉淀后的DNA 样品,-20 ℃保存,备用。
1.3 引物设计和PCR扩增
根据NCBI 报道的黑尾近红鲌线粒体基因组全序列(GenBank 登录号:KC513573),利用 Primer Premier 5.0 设计 nd5 序列引物(F:GTTCATCCATTGGTCTTAGG,R:CTCGTTGAATGTCGCTTGTA),并送成都擎科梓熙生物技术有限公司合成。
PCR 反应体系为 40.0 μL,其中包括 2×Taq PCR Master Mix(成都擎科梓熙生物技术有限公司)20.0 μL,上、下游引物各 2.0 μL(10 mM),模板 DNA 4.0 μL,加双蒸水至总体积 40.0 μL。PCR 反应条件为94 ℃预变性3 min;然后35 个循环,每一个循环包括 94 ℃变性 30 s,55 ℃退火 30 s,72 ℃延伸 2 min,最后 72 ℃延伸 10 min,4 ℃保存。PCR 扩增产物经1.5%琼脂糖凝胶电泳检测后,送至成都擎科梓熙生物技术有限公司完成目的片段回收及测序。

图1 黑尾近红鲌样点分布图Figure 1 The sampling locations of A.nigrocauda
1.4 数据分析
运用DNAstar 软件对所得序列进行校对和拼接,并在此基础上使用MEGA 7.0 进行碱基组成统计、群体间遗传距离的计算;使用DnaSP 5.0 软件计算各群体的单倍型数(n)、变异位点数、单倍型多样性(h)、核苷酸多样性(π)和基因流(Nm)等;Arlequin 3.5 软件计算群体间遗传分化系数(Fst),并进行群体AMOVA 分析。MrBayes 3.1.2 结合 Modeltest 3.7 构建贝叶斯系统进化树。
2 结果
2.1 核苷酸组成
获得黑尾近红鲌nd5 基因序列长度为1 668 bp。如表2所示,nd5 基因序列总体变异率为0.30%,其中,长江干流HJ 群体的变异率高于支流。89 条序列间存在2 个简约信息位点,3 个单一信息位点和2个插入/缺失位点,各群体碱基组成表现出明显的A+T 偏倚性(56.4%)。

表2 黑尾近红 鲌nd5 核苷酸序列特征Table 2 Sequence characteristics of nd5 in A.nigrocauda
2.2 遗传多样性分析
由表3可知,黑尾近红鲌整体呈现低单倍型多样性(h)和低核苷酸(π)的特点,其中,LAR 群体的单倍型多样性最高(0.662),HJ 群体的单倍型多样性最低(0.571);HJ 群体核苷酸多样性最高(0.000 69),LOR 群体最低(0.000 45)。如图2所示,基于 nd5 序列共检测到6 个单倍型,HJ 群体所特有的单倍型数目最多,Hap-1 为3 个地理群体共享单倍型,Hap-2为LAR 群体与HJ 群体共享,其他单倍型为单个群体所特有。
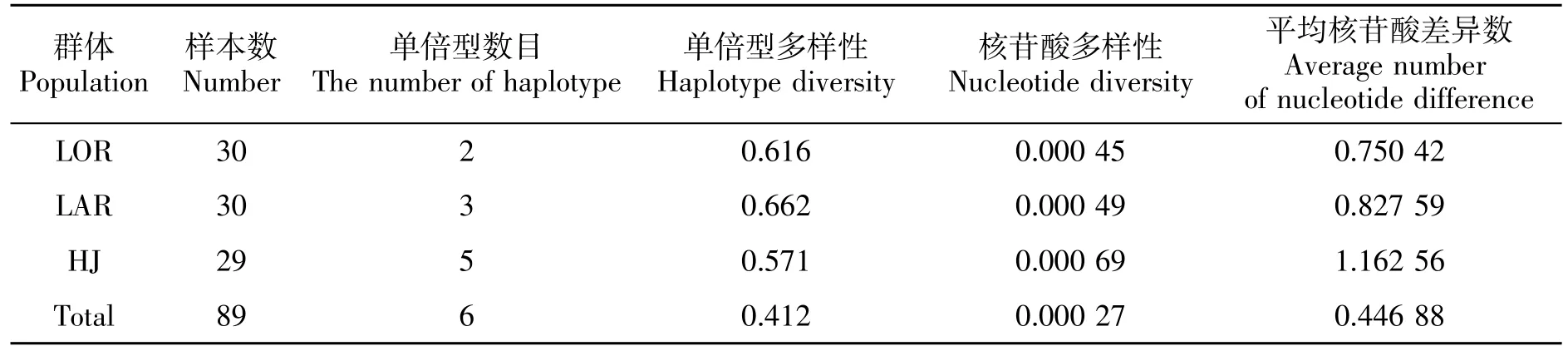
表3 黑尾近红 鲌nd5 基因遗传多样性参数Table 3 Genetic diversity indices of nd5 in A.nigrocauda populations

图2 黑尾近红鲌nd5 单倍型分布图Figure 2 Haplotypes distribution in A.nigrocauda populations based on mitochondrial nd5 sequences.
2.3 群体遗传结构分析
遗传分化系数的结果显示,黑尾近红鲌两两群体间的 Fst值(0.055 54~0.448 28)差异显著(P<0.01,表4)。LOR-LAR 群体间的 Fst值最大,LOR-HJ 群体间的 Fst值最小(表4),这表明 LOR 群体与 HJ 群体间的遗传关系较近。LOR 群体与HJ 群体间的基因流较大(Nm>1),LAR 群体与 LOR 和 HJ 群体间的基因流较小(Nm<1),这表明 LOR 群体和 HJ 群体存在明显的基因交流。分子变异方差分析(AMOVA)表明,分子变异主要来自群体内部(76.12%,表5)。
基于nd5 基因序列构建的系统发育进化树显示,尽管有少数黑尾近红鲌个体的聚类与他们的地理来源不一致,但大部分个体仍是按照他们的地理来源聚类(图3)。
3 讨论
本研究中黑尾近红鲌3 个群体的单倍型多样性(0.412)和核苷酸多样性(0.000 27)均低于Liu H.Z.等[11]和刘春池等[5]在龙溪河、习水河和木洞河的研究结果,同时也表现出两个支流群体(LOR 和LAR)的单倍型数量和核苷酸多样性均显著低于干流群体(HJ,表3)。栖息地破坏、过度捕捞和修建人工水坝等人为活动是造成鱼类遗传多样性、遗传结构不可逆转性恶化的主要因素[14-15],这可能是黑尾近红鲌遗传多样性水平下降的主要原因。3 个黑尾近红鲌种群的栖息环境相差较大,长江干流水流量大,河流形态、生态环境、水生生物多样性和河流连续性均优于支流[16-17],支流生态环境的脆弱,过度捕捞、环境污染等人为干扰对其种群数量所产生的影响较大。近年来黑尾近红鲌野生资源量呈急剧下降的趋势,种群数量的锐减可能是导致遗传多样性快速丢失的主要原因。

表4 黑尾近红 鲌群体内遗传距离(对角线,粗体)、群体间遗传距离(对角线下)和遗传分化系数F(st对角线上)Table 4 Genetic fixation index (Fst) (above diagonal) and genetic distance within (diagonal)and between (below diagonal) A.nigrocauda populations
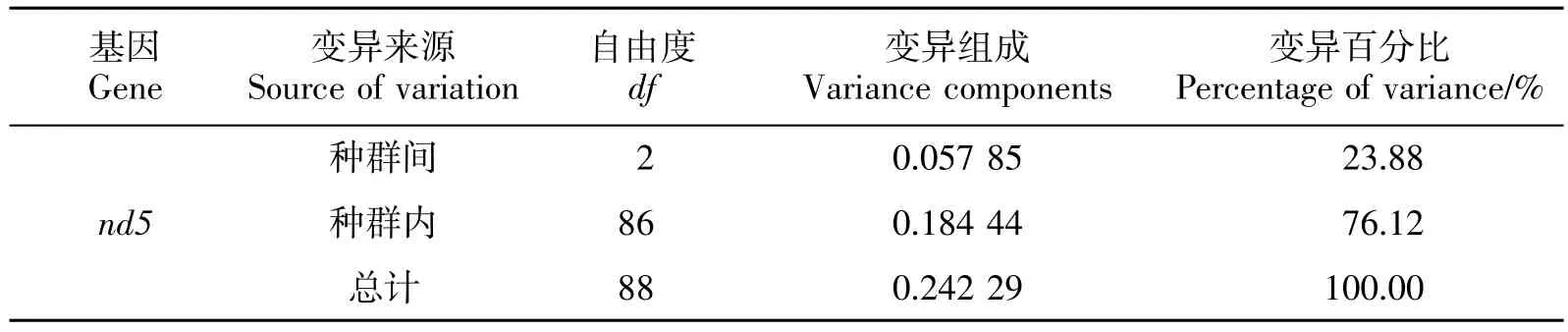
表5 黑尾近红 鲌群体分子变异分析结果Table 5 Analysis of molecular variance (AMOVA) for the A.nigrocauda populations

图3 黑尾近红鲌nd5 基因Bayes 系统进化树Figure 3 Phylogenetic trees of the nd5 sequence of A.nigrocauda reconstructed with Bayesian inference.
淡水鱼类种群很容易受栖息地环境改变、水体污染、水利水电工程的修建和外来物种入侵等因素的影响,从而导致生物多样性下降[18-20]。近年来,在辽河修建了很多引水工程,虽然短期内对鱼类群体没有影响,但长期以来会导致濒危物种生物多样性降低甚至灭绝[20]。本研究结果结合先前的研究结果表明[5,11],在最近 10 多年,研究区域内黑尾近红鲌的遗传多样性下降明显。龙溪河和濑溪河是黑尾近红鲌的主要栖息地,他们分别是长江和沱江的一级支流,水利水电设施的修建使得支流中的黑尾近红鲌与长江干流群体仅能进行单向的基因交流。同时,龙溪河与濑溪河流经的城镇较多,鱼类栖息环境受生活污染、工农业生产等人类活动的影响较大[4,11,21],遗传多样性丢失的速度更快。
遗传分化分析结果表明,3 个种群间存在显著的遗传分化(0.055 54~0.448 28,表4),群体内的变异程度高于群体间(表5)。黑尾近红鲌产粘性卵,受精卵就地孵化发育,幼鱼和成鱼没有长距离迁移特性,黑尾近红鲌的这些特性可能有助于地理种群间的遗传分化。但是,HJ 群体与 LOR、LAR 群体的取样地分别相距大约82 km 和126 km,如此短的地理距离,黑尾近红鲌的产卵习性等是否是其种群间出现遗传分化的主要原因,还待进一步确认。此外,在龙溪河和濑溪河上分别建有7 个和5 个人工大坝,并且这些大坝最早建于20 世纪20年代,严重阻碍了黑尾近红鲌群体间的基因交流,特别是向上基因交流已不可能,因此人工大坝可能也是导致这些种群分化的一个重要原因。从目前的大坝分布情况来看,LOR 群体和LAR 群体可以向下与HJ 群体进行基因交流,但LOR 群体和LAR 群体间几乎完全阻隔,并且LOR 群体和LAR 群体之间的Fst值最大,这也反映了大坝目前的阻隔形势严峻。同时,支流小种群间的遗传漂变和快速的基因丢失也可能加速了种群间的遗传分化[22]。但是,这些大坝的阻隔并不完全,3 个种群间存在一定程度的基因交流(表4),比如支流向下单向基因交流必然存在,同时大坝阻隔时间还相对较短,因此系统进化分析中,少数个体并没有与其自身地理种群聚类(图3)。
综上所述,长江上游黑尾近红鲌群体的遗传多样性下降明显,急需对其开展保护工作。虽然干流群体表现出较高的单倍型多样性水平,在黑尾近红鲌群体的遗传保护中扮演了重要角色,但是支流种群具有种群特有的单倍型,且与干流种群的遗传多样性水平差异显著,同时,支流群体能对干流种群形成较好的资源补充。因此,本研究认为干流和支流都应是黑尾近红鲌重要的保护单元。
