Effect of poly(3-hydroxyalkanoates) as natural polymers on mesenchymal stem cells
2019-10-31VeraVoinovaGarinaBonartsevaAntonBonartsev
Vera Voinova, Garina Bonartseva, Anton Bonartsev
Vera Voinova, Anton Bonartsev, Faculty of Biology, M.V.Lomonosov Moscow State University, Moscow 119234, Russia
Garina Bonartseva, Anton Bonartsev, A.N.Bach Institute of Biochemistry, Research Center of Biotechnology of the Russian Academy of Sciences, Moscow 119071, Russia
Abstract
Key words: Mesenchymal stem cells; Polyhydroxyalkanoates; Poly(3-hydroxybutyrate);Tissue engineering; Differentiation; Natural functions; Microbiota; Bone; Intestine
INTRODUCTION
Mesenchymal stem cells (MSCs) are multipotent progenitor stem cells that can differentiate into multiple mesenchyme cell types, including fibroblasts, osteoblasts,chondrocytes, adipocytes, and myoblasts, and this capacity contributes to the regeneration of mesenchymal tissues such as bone, cartilage, muscle, ligament,tendon, adipose, and stroma.Due to these properties, MSCs are now considered one of the most promising cell types in the field of tissue engineering[1,2].
Tissue engineering is a multidisciplinary scientific, technological direction that combines the latest developments in engineering, materials science, cellular biology,biochemistry, and medicine, thus offering new approaches for the restoration of tissue functions.Tissue engineering is a fundamentally different paradigm in relation to surgery and transplantation.It is based not on approaches related to the substitution or functional compensation of tissue but on tissue regeneration: the organism itself can repair the damaged tissue if the appropriate conditions are provided.Tissue engineering involves an appropriate combination of cells (e.g., MSCs), scaffolds, and bioactive molecules.Together, these interactions allow intercellular communication and cell-biomaterial interactions, which work toward achieving the desired therapeutic response[3,4].
Different types of biomaterials can be used for the development of scaffolds for tissue engineering: metals, ceramics, and synthetic or natural polymers have been used.Nevertheless, biodegradable and biocompatible polymers of natural or synthetic origin, polyhydroxyalkanoates, are believed to be one of the most promising biomaterials for developing scaffolds for bone regeneration.Since the beginning of the 21stcentury, medical devices and dosage forms based on polyhydroxyalkanoates have been actively introduced into medical practices.The following types of chemically synthetic polyhydroxyalkanoates are actively used in clinical practice and scientific research: poly(2-hydroxypropanoic) (polylactic, PLA, polylactides), poly(2-hydroxyacetic) (polyglycolic, PGA, polyglycolides), poly(6-hydroxycaprolactone), and their copolymers, for example, polylactide-co-glycolides (PLGAs) and polymers similar in structure, such as poly-p-dioxanone[5].Natural polyhydroxyalkanoates -poly(3-hydroxyalkanoates) (PHAs) represent polyesters of 3-hydroxyalkanoic acids,e.g., poly(3-hydroxybutyrate) (PHB) is a linear polyester of 3-hydroxybutyric acid.The biopolymer includes only the R-form of 3-hydroxybutyric acid due to which it is a partially crystalline polyester in an isolated and purified form (the PHB crystallinity is 55%-80%).For industry and research purposes, natural PHAs can be produced biotechnologically by bacterial producer strains[6].The following natural PHAs can be distinguished depending on the length of the alkane side radical: PHB, poly(3-hydroxyvalerate), poly(3-hydroxyhexanoate), poly(3-hydroxyoctanoate),etc.During bacterial biosynthesis, as a rule, it is not homopolymers of other PHAs but their blockcopolymers with 3-hydroxybutyrate that are the most frequently obtained: poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV), poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) (PHBHHx), poly(3-hydroxybutyrate-co-3-hydroxyoctanoate),etc.All of them are quite different in their physicochemical properties, such as crystallinity, melting temperature, glass transition temperature, hydrophobicity,plasticity, elastic modulus, and others[7,8].
The chemical structure and the main properties of synthetic poly(2-hydroxyalkanoates) (PLA, PGA, and their copolymers PLGA) are similar to the chemical structure and the physicochemical and biological properties of natural PHAs: the ability to biodegrade in an organism without the formation of toxic products; biocompatibility with human organs and tissues; optimal mechanical properties (relatively high strength, plasticity); other physicochemical properties(thermoplasticity, specific diffusion properties); and the opportunity to use efficient technological processes for their production[5-9].Therefore, the study of natural PHAs(including PHB) as biomaterials for the regeneration of different tissues and organs is of paramount importance.Chemically synthetic PHAs can be considered biomimetic analogs of PHB[5,9].However, it should be taken into account that the properties of natural biopolymers are determined by their functions in nature[10].In particular, the properties of bacteria-origin biopolymers are closely associated with their role in the bacterial world.In most cases, this important information is not taken into account in research on the biomedical applications of various biomaterials.However, why should we not give it proper importance? In this review, we attempt to fill these gaps in the current scientific field by considering the natural properties of polymers in connection with their effect on cells.
A highin vitrobiocompatibility of PHB has also been shown in studies using cell cultures.Therefore, PHB is a promising material for tissue engineering.Cell cultures of various origins, including murine and human fibroblasts, rat, rabbit, and human MSCs, human osteogenic sarcoma cells, human epithelial cells, human endothelial cells, rabbit articular cartilage chondrocytes, rabbit smooth muscle cells, and human neurons in direct contact with PHB when cultured on polymer films and scaffolds exhibited satisfactory levels of cell adhesion, viability, and proliferation.However, the study of the effect of PHAs on the growth and differentiation of MSCs is of particular importance for the use of PHAs as biomaterials for tissue engineering[8,11].
OVERVIEW OF THE EFFECT OF PHAS ON MSC GROWTH AND DIFFERENTIATION
Osteogenic differentiation of MSCs in vitro
It was shown that human[12,13], rat[14-17], and rabbit[18]MSCs isolated from both adipose tissue[14,15]and bone marrow[14-17], murine calvarial preosteoblast cells MC3T3-E1 S14[19], and human induced pluripotent stem cells[20]grown on scaffolds from PHB and its copolymers PHBV and PHBHHx undergo spontaneous differentiation in the osteogenic direction in regular medium.Osteogenic differentiation in osteogenic medium is more pronounced (Table 1).The osteogenic differentiation of MSCs and osteoblastic cells grown on PHA scaffolds was confirmed by a change in cell morphology[13,15,17,18,21], inhibition of their proliferation[14,15,17], an increase in alkaline phosphatase (ALP) activity, deposition of calcium salts[12,15,22], and the expression of markers of osteogenic differentiation and bone formation (ALP, type 1 collagen,Runx2, osteocalcin, osteopontin) determined by enzyme immunoassay and PCR[12,15,16,20].
Chondrogenic differentiation of MSCs in vitro
However, other studies have not confirmed the induction of osteogenic differentiation of MSCs on PHBHHx scaffolds[23-25].It was shown that the spontaneous differentiation of MSCs grown on PHBHHx scaffolds occurs in the chondrogenic direction, which was revealed by a change in the expression of the chondrogenic marker genes of MSCs:Aggreggan,col2,sox9,col10andpthrp[26].PHB, PHBV, and PHBHHx scaffolds support chondrogenic differentiation in the chondrogenic medium[27,28].Chondrogenic differentiation was also demonstratedin vivoafter implantation of PHB scaffolds seeded with MSCs and was associated with the expression ofsox-9,collagen II,aggrecan,safranin-O, and glycosaminoglycans[29].
Proliferation of MSCs in vitro
In spite of stimulation of MSC osteogenic differentiation in regular medium, the proliferation of MSCs grown on scaffolds from PHB and PHBV can both have a higher[18,20,21,30]or lower rate[15,17,28,31]in comparison with controls (e.g., tissue culture plastic) in contrast to the suppression of MSC proliferation in osteogenic or chondrogenic media where MSC differentiation was pronounced[17-20].PHBHHx scaffolds contribute to increasing cell growth[23,24,30,32]but can also cause apoptosis[33](Table 1).
Other types of MSC differentiation in vitro
Moreover, in neurogenic medium PHB[35,37], PHBV[37], PHBHHx[34,36,37],PHBVHHx[34,36], poly(3-hydroxybutyrate-co-3-hydroxyoctanoate)[35], PHB4HB[37], and their composites support the differentiation of MSCs in the neurogenic directioninvitro, which results in a change in cell morphology and expression of genes of specific proteins: Nestin, glial fibrillary acidic protein, and βIII-tubulin[34-37].The differentiation of MSCs in the epidermal (to keratinocytes) and endothelial (to vascular endothelial cells) directions was also demonstrated in appropriate special stimulating media using nanofibrous scaffolds produced by electrospinning as a substrate for cell growth[38,39].

Table 1 The effect of natural poly(3-hydroxyalkanoates) on the growth and differentiation of stem cells

images/BZ_50_235_245_2256_3301.png

PHBV (with 12 mol % 3-HV; Mw= 530000)Scaffolds produced by electrospinning with randomly oriented fibers Human bone marrow MSCs 1, 7, 11 d(proliferation tests); 7 d(differentiation tests)No difference between cell proliferation on scaffolds from PHBV and poly-εcaprolactone(trypan blue assay)Good quality of cell adhesion and spread with developed filopodia.Cells have a nearly spherical shape(CM)Higher modulation of osteogenic differentiation in comparison with MSCs grown on poly-εcaprolactone scaffolds in osteogenic medium (calcein and Alizarin red staining assay)[13]PHBV (with 5 mol % 3-HV, Mw= 680000)Scaffolds produced by electrospinning with randomly oriented nanofibers Human induced pluripotent stem cells 1, 3, 5, 7, 10 d(proliferation tests); 7, 14 d(differentiation tests)The higher proliferation rate in comparison with TCPS (MTT assay)Original-like MSCs formed colonies at day 5(LM).Higher stimulation of osteogenic differentiation in comparison with cells grown on TCPS in osteogenic medium (ALP activity assay, RTPCR for measure of ranx-2, col-1,ALP, osteonectin,osteocalcin expression levels;Western blot assay for osteocalcin and osteopontin expression levels)[20]PHBHHx (with 12 mol % 3-HHx), and PHBVHHx Films casted from polymer solution Human bone marrow MSCs 4, 72 h(proliferation tests); 14, 21 d(differentiation tests)Higher cell proliferation rate(for PHBHHx and PHBVHHx) in comparison with PLA and with TCPS (for PHBVHHx)-No effect on cell differentiation in regular medium(FM: ALP and van Kossa staining)[23]PHBHHx (with 8.3 mol % 3-HHx, Mw =1210000)Smooth compressionmolded films,porous scaffolds casted from solution films,electrospun scaffolds Human bone marrow MSCs 5, 6, 7, 14 d(proliferation tests); 14 d, 5 wk(differentiation tests)The same (for compressionmolded films)and higher (up to 2-fold for solution-casted and electrospun films) cell proliferation in comparison with TCPS Spindle-like,similar to original MSCs, good cell adhesion and spreading in regular medium(FM, SEM).No differentiation in regular medium.Inhibition of differentiation in osteogenic medium(osteocalcin assay, Alizarin red S staining,RT-PCR for measure of collagen I and osteonectin expression level)[24]PHBHHx (Mw =300000)Scaffolds produced by electrospinning with randomly oriented fibers Rat bone marrow MSCs 3 d - The welldeveloped stress fibers spanned the entire cell body and supermature focal adhesions (CM,immunofluoresce nce)No osteogenic differentiation: no significant expression of osteocalcin,osteopontin,osteonectin,runx2, in regular and osteogenic media at day 3(RT-PCR)[25]Chondrogenic differentiation

PHB and PHBHHx in a ratio of 1:2 (by weight)Porous flat scaffolds with pores of 200-300 μm in diameter produced by salt leaching from polymer solution using salt crystals as porogens and lyophilization Human adiposederived stromal cells 7 d (proliferation tests); 14 d, 5 wk(differentiation tests)100% cells viability at day 7(FM)After 1 d, the differentiated cells attached to scaffolds.At 7 d and 14 d, the differentiated cells produced extracellular matrices to fill the voids of the scaffolds (SEM)Chondrogenic differentiation in chondrogenic medium:increased glycosaminoglyca n and collagen content.No chondrogenic differentiation in regular medium:a very slight increase in collagen content(biochemical glycosaminoglyca n and collagen content assays)[27]PHBHHx (Mw =600000)Films casted from polymer solution Murine bone marrow MSCs;chondrocytes isolated from cartilage from knees of mice 1 d (24 h) No change in cell proliferation in comparison with TCPS (RT-PCR analysis of proliferating cell nuclear antigen)-Spontaneous chondrogenic differentiation in regular medium at day 1 (as in chondrocytes with the exception of col1):upregulation of aggrecan, col2,sox9, col10, pthrp,and col1 genes,downregulation of osteocalcin,Cbfa1/Runx2,MMP13 genes,microRNAs miR-29a and miR-29b(alcian blue staining for glycosaminoglyca ns;immunostaining for type II collagen; RT-PCR analysis of chondrogenic markers)[26]PHBV Porous cylindrical(5 mm diameter, 2 mm thick)scaffolds with a pore size of 30-300 μm produced by salt leaching Swine bone marrow MSCs,cartilage progenitor cells,and chondrocytes After 1 wk of in vitro culture subcutaneous implantation in vivo into nude mice for 6 wk.Higher wet weight and volume of the cell-scaffold constructs seeded with cartilage progenitor cells and chondrocytes in comparison with MSCs 6 wk after implantation.Good compatibility between the cells and the scaffold and production of considerable amounts of extracellular matrix after 1 wk of in vitro culture(SEM)Higher chondrogenic differentiation in vivo of cartilage progenitor cells and chondrocytes:increased expression of sox-9, collagen II,aggrecan, safranin-O,glycosaminoglyca ns in comparison with MSCs (alcian blue staining,immunostaining,RT-PCR)[29]

PHBHHx Films casted from polymer solution Human umbilical cord MSCs 3, 7, 14 d(proliferation tests); 14, 28 d(differentiation tests)Higher MSCs proliferation in chondrogenic medium in comparison with TCPS (MTT assay)Good spreading of cells and their proliferation(SEM)Chondrogenic differentiation in chondrogenic medium:increased glycosaminoglyca n, proteoglycan,and collagen content,upregulation of collagen II and aggrecan, genes(amino-sugars and hydroxyproline assays, toluidine blue staining, RTPCR)[28]Epidermal differentiation PHBV, (Mw =450 kDa)Scaffolds produced by electrospinning with randomly oriented nanofibers Bone marrow MSCs 1, 3, 7, 14 d(proliferation tests); 3, 7, 14 d(differentiation tests)100% viability of cells.Higher cell proliferation in epidermalinduction medium in comparison with regular medium.Lower cell proliferation on PHBV scaffolds in regular and epidermalinduced medium in comparison with TCPS (CM,MTS assay)Good adhesion and spreading of cells tended to start forming a monolayer at day 7.The spindlelike, fully extended morphology of MSCs at day 3.The keratinocytic morphology of MSCs characterized by polygonal cells at days 7 and 14(SEM, CM)The epidermogenic differentiation in epidermalinduction medium:increased expression of keratin, filaggrin,and involucrin.No epidermogenic differentiation in regular medium(CM, RT-PCR)[38]Adipogenic differentiation PHBHHx (Mw =300000)Scaffolds produced by electrospinning with randomly oriented fibers Rat bone marrow MSCs 3 d - Well-developed stress fibers spanned the entire cell body and supermature focal adhesions(CM,immunofluoresce nce)No adipogenic differentiation: no significant expression of PPARg, Lpl,ADFP, CD36 in regular and adipogenic media at day 3 (RT-PCR)[25]Endotheliogenic differentiation PHB/PHBV composite(30:70)Scaffolds produced by electrospinning with randomly oriented nanofibers Human adipose tissue-derived MSCs 7, 14, 21 d(proliferation tests); 7, 14, 21 d(differentiation tests)Lower cell proliferation on scaffolds in endothelialinduction medium in comparison with TCPS.Higher cell proliferation on scaffolds in regular medium in comparison with TCPS (CM,MTT assay)Good adhesion and spread,typical spindleshape morphology, and cell-to-cell interactions.Good distribution of cells in regular medium.MSCs in endothelialinduction medium formed circle-like structures characteristic of endothelial cell organization,mimicking the tubular organization of blood vessels at day 21 (SEM, CM,calcein-AM staining)Endotheliogenic differentiation in endothelialinduction medium:increased expression of VECadherin, vWF factor, and VEGFR2(immunostaining,flow cytometry,RT-PCR)[39]Neurogenic differentiation

SEM: Scanning electron microscopy; CM: Confocal microscopy; FM: Fluorescence microscopy; LM: Light microscopy; RT-PCR: Reverse transcription polymerase chain reaction; TCPS: Tissue culture plastic; ALP: Alkaline phosphatase; MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide;MTS: 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium; XTT: 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide; PHB: Poly(3-hydroxybutyrate); PHBV: Poly(3-hydroxybutyrate-co-3-hydroxyvalerate); PHBHHx: Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate); MSCs: Mesenchymal stem cells; PHAs: Poly(3-hydroxyalkanoates); PLA: polylactic acid.
The effect of PHAs on tissue regeneration
The osteoinductive properties of PHB and its copolymers were displayed not onlyin vitrobut alsoin vivowhen scaffolds derived from these polymers were used as bonesubstituting biomaterials and were implanted into bone defects.PHB, the main polymer of a homologous series of the family of PHAs, is the best-known microbiological polyester, which is a promising alternative to biodegradable synthetic thermoplastics and is actively used in regenerative medicine and tissue engineering.The biodegradation rate of PHB and its copolymers is much lower than the biodegradation rate of synthetic PLA, PGA, and their copolymers with the very intensive process of bone tissue reconstruction, which makes PHB use for the regeneration of bone tissue more relevant.Polymer biodegradation mainly occurs due to the phagocytic activity of specialized cells (macrophages, foreign body giant cells,and osteoclasts),i.e.specialized cellular biodegradation of these biopolymers takes place.The greater biocompatibility of PHB and its copolymers compared with synthetic PHAs due to the absence of the effect of acidification of surrounding tissues by the polymer biodegradation products is also of great importance.When the tissue reaction to PHB was compared with tissue reactions to synthetic PLA, PGA polyesters, and their copolymers, several studies found mild or moderate tissue reactions to PHB, while severe chronic inflammatory reactions were not infrequently observed for PLA, PGA, or PLGA[8,11].
Porous scaffolds and patches from PHB were shown to promote bone tissue regeneration, which was demonstrated in experimental models of critical (parietal bone of the rat skull) and noncritical (rat femur) bone defects.At all stages of the process of bone defect regeneration, a minimally pronounced tissue reaction to implantation was observed.This reaction was associated with the gradual bioresorption of the polymeric material by osteoclasts, active vascularization of the scaffolds, and sprouting of the newly formed bone tissue into the pores of the PHB scaffolds[8,11].Regeneration of bone tissue in the PHB scaffolds was also indicated by the elevated expression of osteogenic markers,e.g., collagen type I[15,40].or their expression was in opposite reduced[41].We observed the appearance of the newly formed bone tissue evenly throughout the volume of the porous biopolymer scaffolds in the form of islands, not from its edges, while a fibrous capsule was not formed around the material, indicating complete integration of the biopolymer material with the bone tissue[42].Moreover, it was shown that other bone tissue devices based on PHB (nonwoven patches, porous scaffolds) could contribute to the regeneration of other tissues of various organs, including nerves, intestine, heart, and vessels.A high level of vascularization (angiogenesis) and the increased expression of vascular endothelial growth factor were shown in the tissue defect area during regeneration using PHB devices[8,11,15,41].It should be noted, however, that PHB and its copolymers can activate immune cells during implantation causing the secretion of proinflammatory cytokines by these cells[43,44], which is a typical tissue reaction to implantation of virtually any material.
The problem of the interaction of PHAs and MSCs is closely related to the use of these polymers for the manufacture of medical devices for regenerative surgery and tissue engineering of the intestine using devices from synthetic polyhydroxyalkanoates.The successful transplantation of tubular prostheses on the basis of PGA nonwoven material was carried out to replace a dog esophagus.For this purpose, the prostheses were seeded with fibroblasts and keratinocytes and placed in the abdominal cavity to develop the tissue engineering constructionin situ.Development of the mucosa and muscular membrane was observed after the esophageal substitution, while the prosthesis that was not seeded with cells was rejected[45].
Successful use of the tissue engineering construction based on a surgical mesh fabricated of PGA with a PLA cover as a patch for the stomach wall was demonstrated in a model of stomach wall defects in rats.The surgical mesh was seeded with embryonic epithelial cells, and patch fusion with the stomach wall tissues and the development of mucous and smooth muscle membranes were demonstrated[46].Japanese researchers developed a tissue engineering construction based on a porous tubular prosthesis from PGA seeded with embryonic epithelial intestine cells.In rats, the anastomosis of a tubular prosthesis with rat jejunum was surgically generated to analyze the regenerative potential of the construction, and the development of mucosa with villous epithelium on a polymer scaffold and its integration with the intestine wall tissues were observed[47].With improvements in the endoscopic suturing technique, new opportunities for the application of tissue engineering techniques arise for the regeneration of gastrointestinal tract tissues; good results have already been demonstrated, proving the techniques to be safe and reliable.Takeshitaet al[48]successfully used a poly(6-hydroxycaprolactone) scaffold produced by 3D printing and seeded with fibroblasts to close intestinal wall defects by an endoscopic suturing technique.
Intestinal progenitor stem cells are intensively used for tissue engineering of the stomach, small intestine, and bowel in combination with polymer scaffolds[49,50].However, investigation of intestinal stem cells on scaffolds from PHAs or even synthetic polyhydroxyalkanoates or the development of tissue-engineered devices based on PHA scaffolds seeded with intestinal stem cells are very rare.A good example of such rare studies of particular interest is the work of Costello CM[51]and Shaffiey SA[52].Their porous scaffolds based on PLGA of a complex microarchitecture(modeling the small intestine mucosa villi) were developed for tissue engineering of the intestinal mucosa.The active growth of the intestinal epithelial cells was demonstrated on the obtained scaffolds using both the commercialCaco-2line and cells isolated from the human intestinal mucosa.The development of villus-like,polymer-cellular hybrid structures, the differentiation of cultivated cells in functional cells of the intestinal mucosa (enterocytes, goblet cells, and Paneth cells), and mucus production was demonstrated[51].Then, the researchers developed a tissue-engineered construction of such a complicated structure seeded with coculture of intestinal stem cells, myofibroblasts, macrophages, and probiotic bacteria and demonstrated enhanced mucosal regeneration in the dog rectum upon using this construction[52].
ROLE OF PROPERTIES OF PHAS AS NATURAL BIOPOLYMERS IN THEIR EFFECT ON MSC GROWTH AND DIFFERENTIATION
The role of the microstructure in the effect of PHAs on MSC growth and differentiation
The microstructure and topography of the surface on which cells grow can play particularly important roles in the growth and differentiation of MSCs.Moreover,these features can even reverse the effects of bioactive molecules on cell growth or differentiation[24,25,53].The effect of the biological activity of polymers on the growth and differentiation of MSCs bothin vitroandin vivodepends on the microstructure of 3D devices made from them: surface topography[24], 3D-microstructure[25], porosity,pore size, and shape[9,54].Different cells prefer different surfaces.For example, MSCs and osteoblasts prefer rougher surfaces with the appropriate size of pores[55,56], while fibroblasts prefer smoother surfaces, and epithelial cells attach only to the smoothest surface[57].This appropriate roughness affects cell attachment because it provides the right space for the growth of MSCs or supplies solid anchors for their filopodia.A scaffold with the appropriate size of pores provides better surface properties for anchoring type II collagen filaments and for their penetration into internal layers of the scaffolds seeded with chondrocytes, which is supported by the interaction of extracellular matrix proteins with the material surface[24,25].
However, the study of the proliferation and differentiation of MSCs grown on electrospun scaffolds with randomly oriented and aligned fibers yields contradictory data: the aligned fibers stimulate the osteogenic differentiation of MSCs[25], or there was no significant difference in cell differentiation compared with randomly oriented fibers[16].Moreover, scaffolds with porous surfaces can inhibit the osteogenic differentiation of MSCs cultivated in the osteogenic medium[24].The appropriate surface properties may also promote cell attachment and proliferation by providing more spaces for better gas/nutrient exchange or more serum protein adsorption[9,54].
The scaffolds from PHAs, which are manufactured by a series of techniques (salt leaching, electrospinning, phase separation, blending,etc) have a fibrous porous microstructure, which is a biomimetic structure similar to the extracellular matrix[58].Surprisingly, PHAs are easily processed by various methods to produce such biomimetic structures (Table 1).It may also be related to the natural properties of PHAs, such as the complex structure of bacterial carbonosomes, where the synthesis of PHB is carried out[59].
The role of physicochemical properties in the effect of PHAs on MSC growth and differentiation
It is generally accepted that the effect of PHAs on the growth and differentiation of MSCs is connected with the physicochemical properties of the polymeric material and the 3D and surface microstructure of scaffolds made from this polymer.Indeed, the chemical composition, hydrophilicity, and charge of the polymer surface have significant influences on cell attachment, viability, and proliferation[60].For example,physical or chemical modifications of the surface of PHA devices can improve the attachment and growth of cells on the surface.Lipase treatment increases the viable cell number on PHB films by 100-200 times compared with untreated PHB film.Treatment of PHB films with NaOH also led to a 25-fold increase in the viable cell number compared with untreated PHB film[61].It was shown that treatment of PHB film surfaces with low-pressure ammonia or oxygen plasma improved the growth of human fibroblasts, epithelial cells of respiratory mucosa, and rat bone marrow stromal osteoblastic cells (rat bone marrow MSCs) due to increased hydrophilicity(but with no change in microstructure) of the polymer surface[17,22,62].It was suggested that the improved hydrophilicity of the films after PHB treatment with lipases, alkali,and plasma could allow cells in suspension to more easily attach to the polymer films compared with untreated films.The influence of the hydrophilicity of the biomaterial surface on cell adhesion was demonstrated previously[63].Some researchers used PHAs only as a framework to cover biomaterials and investigated the effect on the growth and differentiation of MSCs.Thus, collagen type I, containing or not containing chondroitin sulfate, was covered on the textile scaffold from PHB threads,and the osteogenic differentiation of human MSCs isolated from bone marrow was studied[64].
Surprisingly, some PHAs (including PHB and PHBV) have piezoelectric properties similar to those of natural bone.Mechanical stress in bone tissue produces electrical signals that stimulate bone growth and remodeling[13].Electrical stimulation resulted in significant increases in the osteogenic differentiation of MSCsin vitroand improved the regeneration of critical bone defectsin vivo[65].Thus, the osteogenic activity of PHB and its copolymers could be due to the piezoelectric properties of these biopolymers.
The physicochemical properties of PHAs are related entirely to the function of natural PHAs as storage polymers in a bacterial cell.The relative hydrophobicity of PHB is due to the elimination of the effect of high osmotic pressure on the storage polymer[6,7,10].The semicrystalline structure and thermoplasticity of PHAs are connected with the necessity to maintain the particular state of the polymer in the cell,which should be simultaneously stable in water medium and available for controlled digestion by PHA depolymerases.Chemical inertness is the main reason for PHA biocompatibility and is associated with the nontoxicity of storage polymers for bacterial cell[6,10,66].
The role of biodegradation as a natural property in the effect of PHAs on MSC growth and differentiation
The ability of PHAs to biodegrade may be the stand-alone reason for the biological activity of these polymers, including the relation with MSCs.As biodegradable polymers, natural PHAs cause targeted activation of macrophages and osteoclasts -cells that directly cause biodegradation of the polymers.It was shown that macrophages and osteoclasts adhere strongly to and proliferate on polymer films.On the polymer surface, macrophages are activated by a polymeric material, which contributes to cell phagocytic activity[67,68].The adhesion of macrophages to the surface of a polymeric material plays a key role: it was shown that biodegradation of polymeric membranes occurs only when macrophages adhere to their surface, and if macrophages cannot adhere to the membrane, the polymer does not degrade[68].
Macrophages can uptake microparticles from low-molecular PHB with a size of 1-10 μm by phagocytosis[67].At high concentrations of PHB particles (> 10 µg/mL),phagocytosis is accompanied by toxic effects and changes in the functional state of macrophages[67], while nanoparticles (15-250 nm) of PHB and its copolymers do not show significant cytotoxic effects on macrophages even at a high concentration of 1 mg/mL unlike nanoparticles from PLA[69].Phagocytosis of microparticles from PHB is accompanied by an increase in the production of nitric oxide and tumor necrosis factor alpha in activated macrophages.Macrophages died after the uptake of a large number of microparticles.It was also demonstrated that phagocytosis of particles from PHB decreases with time due to the active process of PHB biodegradation[67].
In addition to macrophages, fibroblasts and osteoblasts are also capable of uptaking PHB microparticlesin vitro.Even at high concentrations of PHB particles (> 10 μg/mL), phagocytosis is not accompanied by toxic effects on fibroblasts[67].The endocytosis of microparticles by osteoblasts is accompanied by the differentiation of cells in the osteogenic direction as indicated by the increase in ALP activity[70].The PHBHHx nanoparticles were used to study the mechanism of stimulation of cell proliferation.In concentrations from 0.02 to 0.1 g/L, they stimulated an increase in the calcium ion current in the cell cytoplasm, which is one of the main signaling pathways of activation of their division[71-73].
Oligomers of PHAs may also have their own biological activity.Therefore, it was shown that oligomers of PHB and its copolymers with 4-hydroxybutyrate and 3-hydroxyhexanoate (with a chain length of 20-25 monomers) are not toxic to cells (up to a concentration of 20 μg/mL) and have biological activity.They stimulate proliferation, inhibit apoptosis, and suppress the release of calcium into the cytoplasm and the formation of intercellular contacts between the B cells of the pancreas of mice[74].
The process of biodegradation of material by osteoclasts or macrophages can stimulate MSC differentiation in the osteogenic direction.Active osteoclasts grown on biodegradable ceramic scaffolds were shown to influence the development of MSCs into osteoblasts when coculturedin vitroby STAT3 activation[75,76].Therefore, the ability of PHA to biodegrade itself may be an indirect cause of the induction of MSC differentiation.The ability of hydrolytic and enzymatic degradation (with slow rate) is strongly connected with the need for controlled consumption of this polymer as an energy supply by enzymatic cleavage in special granules of a bacterial cell[6,10,59].
Moreover, the products of PHA biodegradation are common substances in human organisms because of the presence of PHB and its copolymers in bacteria of the human microbiota.A series of symbiotic and infectious bacteria of mammals(including human) of the genusesAgrobacterium, Clostridium, Ralstonia, Bacillus,Burkholderia, Vibrio, Legionella, Pseudomonas, Mycobacterium, Acinetobacter,Sphingomonas, Fusobacterium, Neisseria, Streptomyces, andBordetella, Rickettsiacan synthesize this biopolymer or have the enzymes (or their genes) of PHB biosynthesis,PHA-polymerase.
Some of these bacteria, such asPseudomonas sp.are capable of synthesizing not only PHB but also various PHB copolymers[66].The role of this biopolymer in the bacteria of animal microbiota remains a very poorly understood problem.However, a number of papers have suggested the participation of PHB in the relationship between bacteria and the host organism.Kimet al[77]suggested that the PHB synthesis facility allows symbiotic bacteria of theBurkholderiagenus to survive in the intestine of theRiptortus pedestrisbean bug in the stress conditions induced by the host organism immune system to regulate the number of these bacteria.Apparently, PHB synthesis by bacteria fromRhodobacteralesmodulated the gastrointestinal tract microbiota in sea cucumbersApostichopus japonicuscontributing to an increase in the animal size multiple times[78].
Moreover, the efficiency of PHB in the fight against infectious diseases in animals was demonstrated: the use of PHB powder as an additive to feed protectedArtemia naupliicrustaceans from infectious disease caused by pathogenicVibrio campbelliibacteria.The efficiency of PHB was 100 times larger than that of the 3HB monomeric precursor[79].In addition, PHB has the ability to suppress pathogenic bacteria, not onlyVibrio sp.but alsoE.coliandSalmonella sp.[80].PHB oligomers (with a length of approximately 80 3HB monomers) possess effective concentration-dependent antibacterial and antifungal properties againstStaphylococcus aureus,Klebsiella pneumoniae, andCandida albicansthrough disruption of the bacterial wall/membrane and changes in the transmembrane potential[81].
At the genus level, dietary PHB increased the abundance of beneficial bacteria in the intestine of Pacific white shrimpLitopenaeus vannamei, such asBacillus,Lactobacillus,Lactococcus,Clostridium, andBdellovibrio.This beneficial effect was associated with activation of the rapamycin (mTOR) signaling pathway, which plays a crucial role in intestinal inflammation and epithelial morphogenesis[82].However, the mTOR signaling pathway is involved in MSC proliferation and differentiation.Suppression of the mTOR signaling pathway enhances the osteogenic capacity of stem cells, while mTOR activation causes MSC hyperproliferation[83,84].
Moreover, the modified products of PHB biodegradation methyl-esterified dimers and trimers have pronounced antioxidant activity: 3-fold greater hydroxyl radical-scavenging activity than glutathione and 11-fold higher activity than vitamin C or 3HB monomer[85].Unfortunately, the role of possible antioxidant properties in biocompatibility or biological activity of PHAs is unexplored, while it can be also be connected with their effect on MSCs.Oxidative stress is one of the regulatory mechanisms for MSC osteogenic and adipogenic differentiation.For example,antioxidant vitamin D3 promotes the osteogenic differentiation of MSCs under normal conditions and partly protects cells from oxidative stress damage by activating the endogenous antioxidant system[86,87].Maybe this is one of the main reasons for PHA biocompatibility, especially with stem cells.But if PHAs have their antioxidant properties as their natural functions,e.g., in bacterial cells[85], it can explain the mechanism of interaction of PHAs with stem cells.
However, the possible effect of intestinal microbiota bacteria on PHA polymeric devices (and the effect of the polymeric material of these devices on symbiotic bacteria) is not always taken into account when polyhydroxyalkanoates are used to regenerate intestinal tissues.In particular, the roles of natural PHAs and the degradation products in the interaction with intestinal progenitor stem cells and microbiota bacteria remain entirely unexplored.However, the fundamental roles and functions of PHB in relation to human intestinal microbiota bacteria should be taken into account in the development of medical devices based not only on PHB and other natural PHAs but also on their biomimetic analogs, such as synthetic polyhydroxyalkanoates, especially devices based on these polymers for intestine surgery.
The role of 3-hydroxybutyrate in the effect of PHAs on MSC growth and differentiation
The effect of PHAs on the differentiation of MSCs can also be associated with the bioactivity of 3HB that is produced during PHB and its copolymer biodegradation as the primary monomeric precursor of these polymers.Thus, 3HB at a concentration of 0.01 to 0.1 g/L (0.1-1 mmol/L) can cause activation of the proliferation of human keratinocytes of theHaCaTline and mouseL929fibroblasts by increasing the concentration of calcium ions in the cell cytoplasm and by inhibiting the apoptosis and necrosis of fibroblasts[71-73].Such bioactivity of 3HB is not surprising because 3HB is a natural metabolite in the mammal body.It is the so-called ketone body with a pronounced versatile biological activity.For example, 3HB performs regulatory functions under various special conditions such as diabetes, starvation, and a ketogenic diet[88,89].3HB at a concentration of 0.005-0.1 g/L (0.05-1 mmol/L) induced osteogenic differentiation of mouse MC3T3-E1 preosteoblast cells (unfortunately, not MSCs), which was determined by an increase in ALP activity, calcium deposition(Alizarin red S dye assay), and osteocalcin expression.It should be noted that PHAs enhanced osteogenic differentiation of both MSCs and exactly this cell line of preosteoblast cells.
Moreover, it was demonstrated that 3HB hasin vivoosteoinductive activity that was shown in an osteoporosis model in female rats with removed ovaries.However,at lower concentrations, 3HB did not have a similar effect.For slower PHA biodegradation, 3HB formed at concentrations much lower than 0.05 mmol/L[90].It was also shown that 3HB stimulates the formation of gap junctions between neurons for the transmission of electrical signals, which is associated with improved memory and learning[91].The ability of PHAs to cause neurogenic differentiation of MSCs could also be related to the previously shown neuroprotective effect of 3HB.3HB manifests itself at very high doses because the effect of 3HB on the nervous system is due to the nutrient (energy) function of fatty acids for neurons[92].
Furthermore, if PHAs cause differentiation of MSCs only by 3HB biological activity, it is difficult to explain the data of a series of investigations in which the growth and differentiation of MSCs were compared among scaffolds fabricated of different PHAs (PHB and its copolymers usually).The effect of different PHAs on the growth and differentiation of MSCs differs significantly and even dramatically, while the content of 3HB monomers in PHB copolymers is rarely lower than 80 mol%[18,23,33,93].It was shown that 3HB functions through multiple mechanisms: inhibition of class I histone deacetylases; binding and activation of cell surface G-proteincoupled receptors, hydroxycarboxylic acid receptors, and free fatty acid receptor 3;histone modification by covalent binding with proteins (β-hydroxybutyrylation); and membrane channel and transporter regulation[89].The product of biodegradation of synthetic PLA and PLGA - L-lactate (2-hydroxypropanoate) was shown to be a natural ligand for Gi-coupled G-protein receptor 81 inhibiting cAMP-mediated intracellular signaling events such as lipolysis[94].
It was also revealed that the biodegradation products of some natural PHAs (e.g., 3-hydroxyoctanoate) have antimicrobial activity against a number of infectious gramnegative and gram-positive bacteria.They inhibit the production of metabolites associated with their pathogenic activity.However, their cytotoxic activity against human fibroblasts is manifested at much higher concentrations[86].
Short-chain fatty acids (including 3HB and L-lactate) are the critical metabolites of mammalian microbiota by which symbiotic bacteria execute the diet-based microbial influence on the host.They affect various physiological processes in the gastrointestinal, nervous, and immune systems and may contribute to health and disease (colitis, diabetes, cancer, asthma)[95].Thus, the observed effect of 3HB on MSC growth and differentiation could be associated with one of the mechanisms of microbiota influence on mammalian host organisms.
The possible role of natural signaling functions of PHB in the effect of PHAs on MSC growth and differentiation
Notwithstanding the above, data from a series of studies provide evidence of the biological activity of PHAs toward cells (including MSCs) through receptor or cellular signaling molecule-associated mechanisms[25,26,33,96].As we can see in Table 1 PHAs induce osteogenic and chondrogenic differentiation and support MSC differentiation in these and other directions: epitheliogenic, adipogenic, endotheliogenic, and neurogenic.It also can change the MSC proliferation rate, which is associated with change in cell morphology, physiological state, extracellular matrix synthesis, and expression of a series of differentiation markers: ALP, collagen I and II types,osteopontin, osteocalcin, glycosaminoglycan, aggrecan, sox-9,etc.The possible mechanism of the implementation of the effects of PHAs on cell proliferation,differentiation, and apoptosis can be accomplished through integrins - intercellular macromolecules for contact and recognition.It was shown that MSC differentiation and apoptosis of osteoblasts is carried out by a cascade mechanism that includes regulator proteins such as peroxisome proliferator-activated receptor gamma, which is triggered by the interaction of PHAs with integrins on the cell surface[25,33].It was also shown that the osteogenic and chondrogenic differentiation of MSCs induced by the polymer surface of PHAs could be regulated by microRNAs[26,96].The microRNAs miR-29a and miR-29b were confirmed to be significantly downregulated in MSCs grown on PHBHHx films through cell-material interactions.They can directly target the 3’ UTR ofcol2a1and are suppressed by the chondrogenic proteinSox9to induce chondrogenesis in MSCs[26].
Some researchers attribute the biological activity of PHB and its copolymers obtained by bacterial biosynthesis to the fact that the polymer biomaterials may be poorly purified from bacterial lipopolysaccharide or DNA.However, a previous study made clear that even a very well-purified polymer is able to activate the cellular response[68].
The problem of the biological activity of PHAs,e.g., their ability to effect stem cell growth and differentiation, seems to be closely related to the problem of endogenous PHB in mammalian tissues.The finding of bacterial PHB in various tissues of mammals was one of the most exciting stages in the history of PHA research.It should be immediately noted that it is not a high-molecular reserve polymer.The synthesis is typical for a number of bacteria, but the so-called short-chain complexforming PHB (cPHB) and low-molecular oligo-PHB (oPHB) were found[59].While specific PHB biosynthesis enzymes are available only in prokaryotes, this biopolymer was found in organisms of almost all types.Short- and medium-chain PHBs were found in many different mammalian organs and tissues (including in humans),i.e.in the blood plasma, heart, kidneys, liver, vessels (aorta), nerves, lipoprotein particles,platelets,etc.It was demonstrated that oPHB is located in eukaryotic cell membranes in the form of a PHB-polyphosphate-calcium complex.The oPHB has a molecular weight of 12200, which corresponds to a length of approximately 140 elements of 3HB.The oPHB concentration in human blood plasma can change in a rather wide range, from 0.6 to 18.2 mg/L at an average value of 3.5 mg/L[97].
Thus, what function does PHB have in the mammalian organism? In addition to the role of PHB as a reserve substance and energy depot in bacteria, the researchers who discovered endogenous PHB in mammalian tissues suggested some of its functions.The functions of endogenous PHB for both prokaryotic and eukaryotic organisms are apparently coupled with the regulation of different proteins due to the production by short-chain cPHB and oPHB of both noncovalent and covalent bonds with other biopolymers (proteins, nonorganic polyphosphates, and DNA).In the form of PHBCa-polyphosphate complexes, cPHB can function as nonprotein channels capable of conducting inorganic ions through the plasma membrane; these structures also form noncovalent complexes with channels proteins and become part of them)[97].It was demonstrated that cPHB binds to one of the proteins from the group of melastatin receptors (TRPM8) of mammals (and humans), which leads to a change in its functioning[97,98].TRPM8is a membrane calcium channel that functions as a temperature sensor of neurons of the mammalian peripheral nervous system.The binding of the proteinTRPM8receptor channel with cPHB is required for normal functionality of this receptor, which is associated with a change in the conformation state of PHB when passing through this polymer’s glass transition temperature, which is approximately 10 °C[59,97,98].
Another case of the modification of proteins is theOmpAprotein ofE.coli.The covalent attachment of cPHB to the specific serine residues ofOmpAmaintains the correct integration and orientation of this protein in the outer membrane of the bacterium.It was proposed that such modification of proteins by cPHB involves the folding and sorting of certain proteins[99].Similar processes have been described for mammals.Thus, it was shown that PHB oligomers are covalently bound to the calcium ATPase of the plasma membrane of human erythrocytes, simultaneously forming a complex with inorganic phosphates[100].
Other researchers have indirectly confirmed that such conjugation of cPHB with proteins may have a specific physiological function.It was shown that conjugation of the antitumor peptides DP18L with 3-hydroxydecanoate enhances their antitumor activity[101,102].The formation of covalent bonds of PHB with proteins during biochemical processes can be indirectly confirmed by the ability of many bacterial strains, producers of high-molecular-weight PHB, to utilize other polymers added to the culture medium, which often differ significantly in their physicochemical properties (for example, hydrophilic PEG) by covalent crosslinking with the end of the synthesizing PHB chain and the formation of diblock copolymers (for example,PHB-PEG)[103].
There is a striking example of other biopolymers with such functionality that, in addition, are quite similar to PHB in chemical structure: polyprenols (in bacteria,plants, and fungi) and dolichols (in bacteria and animals).Polyprenylation (covalent binding to polyprenols) plays an essential role in the posttranslational modification of a number of proteins to anchor them in the membrane.All organisms (both eukaryotes and prokaryotes) have biochemical mechanisms for the synthesis of these biopolymers and their conjugation with proteins[88].Appropriately, there is evidence suggesting that the functionality of cPHB and oPHB in eukaryotes could be similar to the functionality of polyprenols and dolichols.Thus, according to these data, PHB can theoretically be used to regulate the effect of some cytokines or growth factors on stem cells (including nonspecifically) by covalent modification of appropriate receptors by this biopolymer.
Could PHB perform some type of signaling function for microbiota bacteria when communicating with eukaryotic cells (including stem cells) of the host organism? In general, can bacteria of animal microbiota affect stem cells? Very little is known about the influence of microbiota bacteria of human and laboratory animals on the growth and differentiation of stem cells (including MSCs), but some mechanisms of these interactions have been revealed.It was shown that peptidoglycans and polysaccharides of the bacterial cell wall could alter the functional activity of MSCs.They can stimulate the immunomodulatory activity of MSCs (production of cytokines), accelerate or inhibit the growth of MSCs, and regulate the differentiation of MSCs in adipogenic, chondrogenic or osteogenic directions[104-107].It was revealed that this biochemical response is realized through the activation of specific intracellular MSC receptors,NOD1andNOD2and receptors of theTLRfamily[104,106,108,109].It should be noted that in cells of the immune system (for example,macrophages), activation of these receptors led to stimulation of the production of proinflammatory cytokines and an antibacterial response.It was shown that the microbiota of the oral mucosa stimulated the differentiation and development of osteoclasts by activating the regulatory genesTLR2,TLR4,NOD1, andNOD2by peptidoglycans and polysaccharides of the bacterial cell wall.This led, to bone resorption, which plays an essential role in the pathogenesis and treatment of periodontal disease[110-113].In contrast, in MSCs, activation of these receptors leads to more complex and diverse physiological responses, such as the suppression of chronic inflammation, which can contribute to the treatment of colitis and dermatitis[108].
Some signaling substances of microbiota bacteria can simultaneously act as intermediaries in the regulation of PHB biosynthesis with bacterial communication with the host organism that deserves attention.Thus, the ability of histamine to regulate the synthesis of low-molecular-weight cPHB inE.colihas been demonstrated.Histamine plays an important role as a means of bacterial communication with the host organism and a regulator of the gastrointestinal tract immune system allowing bacteria to be considered “native” to the host organism.Therefore, the effect of histamine on cPHB synthesis can indicate the involvement of this biopolymer in the processes of adaptation and coexistence with the host organism[114,115].However, it is well known that histamine is one of the most important mediators of an immune response to foreign pathogens that is produced by basophils and mast cells.Histamine is involved in the inflammatory response and has a central role as a mediator of itching ( it acts as a neurotransmitter).Surprisingly, it was shown that the decrease in expression of histamine H2R receptors in human MSCs led to the suppression of cell osteogenic differentiation, which was detected by decreased expression of the osteogenic markers, osteocalcin, bone morphogenetic protein 2, and runt-related transcription factor 2[116].
Moreover, storage PHB and short-chain PHB as bacterial signal molecules can directly affect the differentiation of intestinal stem cells into mucosa enterocytes(absorptive cells).It was shown that intestinal stem cells grown in intestinal organoidsin vitroreadily uptake nanoparticles from synthetic PHAs (PLGA)[117], whereas PLGA scaffolds support their differentiation in enterocytes, goblet cells, and Paneth cells[51,52].It should be noted that alkaline phosphatase and bone morphogenic protein are the markers of differentiation both of intestinal stem cells into enterocytes and MSCs into osteoblasts[2,4,118].Maybe the effect of PHA scaffolds on MSC differentiation is caused by mimic action of PHAs on ALP- and BMP-pathways.
In Figure 1 we summarized the above speculations about the association of PHA natural functions and their effect on stem cells.The possible interaction of PHAs as bacteria-origin signal molecules with microbiota bacteria, intestinal stem cells, and immune cells was demonstrated in association with the effect of implanted devices from PHAs on MSCs.
The piezoelectric properties of PHB and its copolymers could also be associated with the natural functions of these biopolymers for bacteria of mammalian microbiota.There are a number of electroactive bacteria in animal microbiota, and they use electrical stimuli not only for energetic purposes but also for communication[119,120].Moreover, some of the electroactive bacteria of mammalian microbiota, such asPseudomonas aeruginosa, can synthesize and accumulate PHB and its various copolymers[121].Considering that PHB can be covalently linked to receptors of both bacterial and eukaryotic cells[98-100], it cannot be excluded that the piezoelectric properties of the polymer can be used by microbiota both for communication among bacteria of the microbiota and with other cells of the host organism, including intestinal stem cells.
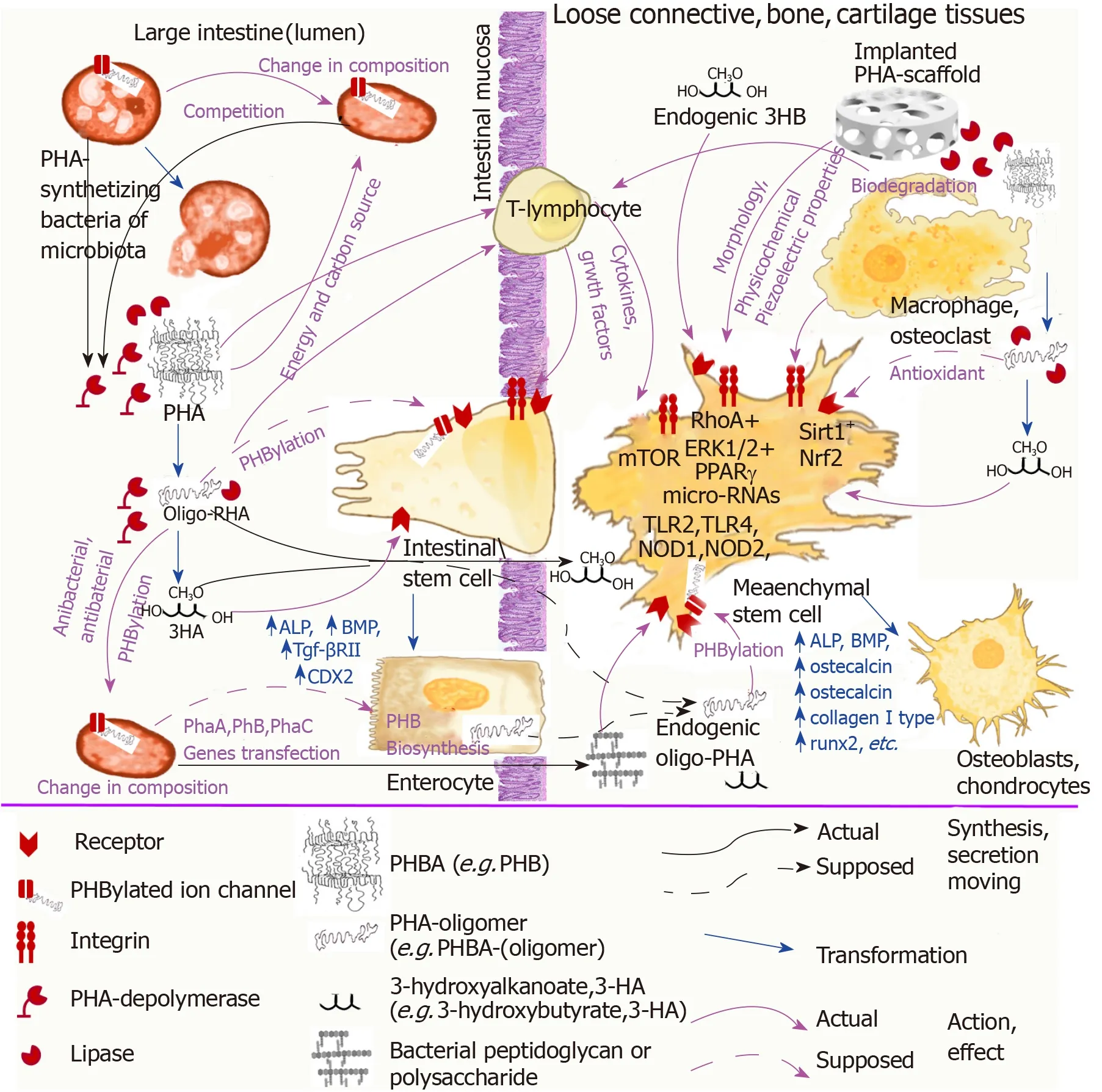
Figure 1 The scheme illustrating the association of PHA natural functions with the effect of implanted PHA scaffolds on mesenchymal stem cells differentiation.
The possibility that PHB can perform some type of signaling function in mammalian tissues can be perceptualized by the fact that the dimers and trimers of 3-hydroxybutyrate are sex pheromones in spiders[122].Moreover, it is possible that these pheromones could also be products of the bacterial biosynthesis of the arthropod microbiota.For example, in the beetleCostelytra zealandica, the sex pheromone is phenol, which is produced from tyrosine in special glands by symbiotic bacteria Morganella morganii[123].Although the dimers and trimers of 3-hydroxybutyrate were found in the fungusHypoxylon truncatum, the mechanism of their synthesis was not disclosed[124].
A better understanding of the mechanisms of interaction of PHAs with MSCs is the key to developing new therapeutic agents based on them.For example, if PHB causes the osteogenic differentiation through activation of some receptor of MSCs and PHBHHx causes the chondrogenic differentiation of MSCs through activation of another receptor of MSCs, then the medical devices based on PHB should be developed specifically for bone regeneration and medical devices based on PHBHHx-specifically for cartilage regeneration.But if the osteogenic differentiation of MSCs is caused by the main product of PHAs biodegradation - 3-hydroxybutyrate,and chondrogenic differentiation of MSCs is caused by some kind of microstructure of PHBHHx medical devices, then it is better to use any type of PHA depending on their physicochemical properties (the strength and elasticity of PHB and PHBHHx differ significantly) for bone regeneration and medical devices based on any type of PHAs with desired microstructure for cartilage regeneration.
At the first case of receptor-mediated induction of MSC differentiation, it can be connected with the similar receptor-mediated action of bacteria-origin PHAs when bacteria interact with each other or with the cells of the host organism that can help to discover mechanisms of PHAs action regarding signal functions and binding with receptors of PHB oligomers.At the second case, the bone regeneration process is mediated by internal product of lipid metabolism (3-hydroxybutyrate and connected with mechanisms of its metabolism).Maybe it is better to regulate 3-hydroxybutyrate metabolism to improve bone regeneration.The same is with the microstructure of PHAs devices because in this case, it is necessary to find the specific structural element regulating chondrogenic MSCs differentiation independent of the type of biomaterial at all.We should keep in mind that elasticity of biomaterials can also affect MSC differentiation[125].
Moreover, a more in-depth understanding of the interaction of PHAs with MSCs in relation to natural functions of these biopolymers can help to develop novel types of medical devices and pharmaceutical formulations based on them: medical devices with selective osteoinductive or chondroinductive activity and desired regenerative activity; prebiotics that promote regenerative activity of the intestinal wall through modulation of microbiota and stimulation of intestine progenitor stem cells;substrates for MSC cultivation in bioreactors; formulations for sustained delivery of drugs with biopolymer-compatible natural bioactivity; experimental model systems for MSC cultivation for drug testing; or therapeutic tissue-engineered systems simultaneously containing probiotic bacteria and MSCs as active ingredients for treatment of gastrointestinal system diseases.
CONCLUSION
PHAs have integral multifactor effects on MSCs through different mechanisms related to the main characteristics of PHA scaffolds that are difficult to separate from each other: the physicochemical properties of the polymer surface; the morphology of polymer devices; and the possible biological activity of these biopolymers.However,the intrinsic biological activity of natural PHAs plays an important role in the effect of implanted PHA devices on the proliferation and differentiation of MSCs as well as on regeneration of bone, cartilage, loose connective tissue, and other tissues.
In summary, studies that consider the problem of the influence of natural PHAs on the growth and differentiation of MSCs should be carried out in a manner that considers and integrates the functions of these biopolymers in nature.We suggest that these biopolymers of bacteria origin engage in some unexplored mechanism while signaling functions in our body through which microbiota bacteria can“communicate” with cells of the immune system, intestinal mucosa, and other tissues causing them to induce a physiological response.PHAs as bacterial signaling molecules can be involved in regenerative processes of intestinal mucosa regulated by microbiota bacteria through stimulation of intestinal stem cells differentiation into enterocytes and other mucosa epithelial cells and directly (as absorbed or endogenous oPHB) or indirectly effect the proliferation and differentiation of MSCs (Figure 1).
The above data on the participation of intercellular contact molecules such as integrins, microRNAs, the mTOR signaling pathway, antioxidant activity, and piezoelectric properties in the implementation of the biological effects of PHB and data on receptor regulation through the covalent binding of PHB to proteins can indicate the natural signaling function of this biopolymer.Unfortunately, this fascinating problem is studied very poorly.There is only very fragmentary and incomplete information.A broader understanding of the role of PHAs in nature could help to further clarify the unusual or contradictory data in the field of the interactions of PHAs with MSCs and to develop novel medical devices and pharmaceutical formulations based on them.
杂志排行

World Journal of Stem Cells的其它文章
- Microfluidic three-dimensional cell culture of stem cells for highthroughput analysis
- Neural stem cell transplantation therapy for brain ischemic stroke:Review and perspectives
- Unmodified autologous stem cells at point of care for chronic myocardial infarction
- Characterization of inflammatory factor-induced changes in mesenchymal stem cell exosomes and sequencing analysis of exosomal microRNAs
- Stem cell treatment and cerebral palsy: Systemic review and metaanalysis