荒漠草原不同土壤条件下猪毛蒿幼苗种群的点格局分析
2019-10-24李月飞李学斌张义凡
李月飞,陈 林,李学斌,张义凡,苏 莹
1 宁夏大学西北退化生态系统恢复与重建教育部重点实验室,银川 750021 2 宁夏大学西北土地退化与生态恢复省部共建国家重点实验室培育基地,银川 750021 3 宁夏大学西部生态与生物资源开发联合研究中心,银川 750021 4 宁夏大学科学技术处,银川 750021
当斑块的特征在某种程度上可以量化并能够被预测时,称之为空间格局。它是植物种群自身特性、种间关系及环境条件综合作用的结果,是种群的基本特征[1-2]。种群空间分局格局能在很大程度上反映种群内个体的空间配置特点[3],种群与生境的关系及其在群落中的地位和作用[4]。分析种群分布格局有助于认识种群的生物学特性(如生活史策略、喜光、耐阴)、生态学过程(如种子扩散、种内和种间竞争、干扰等)及其与环境因子之间的相互关系具有重要作用[5-6],因此一直是生态学家研究的热点。空间格局对种群密度和空间尺度均有很强的依赖性,不同尺度获得的格局信息存在很大差别[7]因此通常依靠特征空间尺度进行的传统格局分析方法,难以推广或者应用到其它尺度上去[8-12]。而点格局克服了传统方法只能分析单一尺度空间分布格局的缺点,目前已被广泛适用于种群格局的分析中[13-15]。
点格局分析方法是由Ripiey[16]首先提出后,国外研究者在Ripley K(t)函数分析法的基础上发展出了O-ring O(t) 函数分析方法,能够最大限度地利用空间信息来分析植物种群由小到大一系列连续尺度上的空间分布格局。它是将植物种群中的每一个个体都看做一个空间坐标中的点,以空间分布的点位图为基础进行点格局分析[17]。张金屯[1]首先将点格局分析引入国内,推动了植物种群空间分布的研究。马志波等[18]统计了1996—2015年期间以点格局为题的国内外中文核心期刊。结果表明,国内的生态学格局的应用研究占据主导地位,研究对象包括以树木为主的乔、灌、草等不同生活型的植物,甚至包括景观。但概括性统计量、零模型与点过程模型等方面研究薄弱。概括性统计量使用单一,且以Ripley的K函数及其变形为主。为了对种群分布格局进行更为深入和清晰的分析,合理的解释实测数据与零模型的偏离,Wiegand等[19]提出了基于不同零模型的分析方法。王鑫厅等[20]认为选择合适的零模型是解决生态问题的关键,仅通过完全随机模型的检验来分析种群的格局,或许很难论证复杂的生态过程[21-22]。在对羊草-大针茅草原退化群落优势种群空间点格局分析中,应用了完全随机模型、泊松聚块模型和嵌套双聚块模型分析了退化草原中优势种群的空间格局,发现了群落中4个优势种群均呈现在大聚快中分布着较高密度的小聚块,这是种群适应过渡牧压的一种表现形式[21]。高福元等[3]基于不同零模型分析了小尺度下三江平原沼泽湿地主要优势种发现,竞争是引起格局变化的重要因素,同时发现实测值偏离完全随机模型的程度越大,越有可能符合泊松聚聚块模型,偏离泊松聚块模型的程度越大,越有可能符合嵌套双聚块模型。在空间格局的测定方法中,邻接格子样方法自设计以来,因具有样方取样的有点,能克服单一取样尺度带来的影响而受到广大生态研究者的推崇。但这种方法不能清晰的再现空间格局,因此王鑫厅基于寻找一种既能直观观察和分析植物个体如何分布,又能测度种群空间分布格局的类型的方法的需求,提出了摄影定位法[23],同时验证了其可靠性与准确度。摄影定位法将繁重的室外工作搬入室内,利用地理信息系统获得数据信息,工作效率大幅提高,也节省了大量的时间和花费。更为重要的是可以把演替的群落、种群尺度监测细化到个体水平,可以分析不同尺度上的动态格局,获取更为丰富的生态学信息。不过该方法存在一定的局限性,但对于草原种群格局的研究具有更为广阔的前景。
在气候条件一致的情况下,土壤异质性决定了种群空间分布的差异[24-25]。杨洪晓等[8]对毛乌苏沙地油蒿种群点格局的分析中得出,油蒿的空间分布格局与生境有密切的联系。南笑宁等[26]黄土高原中部红砂与荒漠锦鸡儿种群空间分布格局的研究发现,红砂与荒漠锦鸡儿在不同坡向的分布具有显著的差异。荒漠草原土壤资源存在广泛的空间异质性,土壤类型也多样。植物种群的空间分布格局不仅是因种而异,也存在于同一种群不同生活史阶段[27]。有研究表明,同一物种,在其生长发育的不同阶段,其空间分布格局也不相同,幼苗具有更强的环境敏感性[27-28]。猪毛蒿为菊科蒿属植物,多年生草本或一二年生草本[29]。因具有较强适应恶劣环境能力而成为主要先锋物种和自然演替序列早起的优势建群物种[30], 目前学者对于猪毛蒿植物科学、化学和药理学/配药学研究较多[31]。但对于猪毛蒿幼苗种群在不同土壤条件下的的空间分布格局尚未见报道。鉴于此,本实验采用摄影定位法[23],基于不同零模型的点格局分析方法,对宁夏荒漠草原不同土壤条件下猪毛蒿种群生活史初期分布格局进行系统探讨,分析形成格局的原因,以期为进一步揭示猪毛蒿种群和群落的形成以及潜在的维持机制提供理论依据[32]。
1 材料与方法
1.1 研究区概况
研究区位于宁夏盐池县皖记沟村(37°47′—37°57′ N,107°22′—107°33′ E),地貌为鄂尔多斯缓坡起伏高原,地势南高北低,土壤主要类型为灰钙土、风沙土和基岩风化残积土等。土壤质地以沙壤、粉沙壤和沙土为主。该地区属于中温带半干旱区,典型中温带大陆性气候。年平均气温8.2 ℃,多年平均降水量289 mm,降水的年际和年内变化剧烈(农牧交错带村域土地利用变化及驱动机制研究——以盐池县皖记沟村为例)7—9月份降水量占全年的60%以上,潜在年蒸发量2014 mm。年无霜期为165 d.
1.2 样地选择及测定
在盐池皖记沟村围封3年的草地围栏中选取3块10 m×10 m地表平坦、群落外貌均匀且具有代表性的猪毛蒿群落为样地。2003年以来,宁夏全面实施围封禁牧,给草场提供了一个休养生息的机会,部分草地生态环境出现逆转。所选样地围封前为自由放牧的草地,由于食草牲畜的啃食和践踏,地上植被生物量较低,围封后具体情况如表1所示。
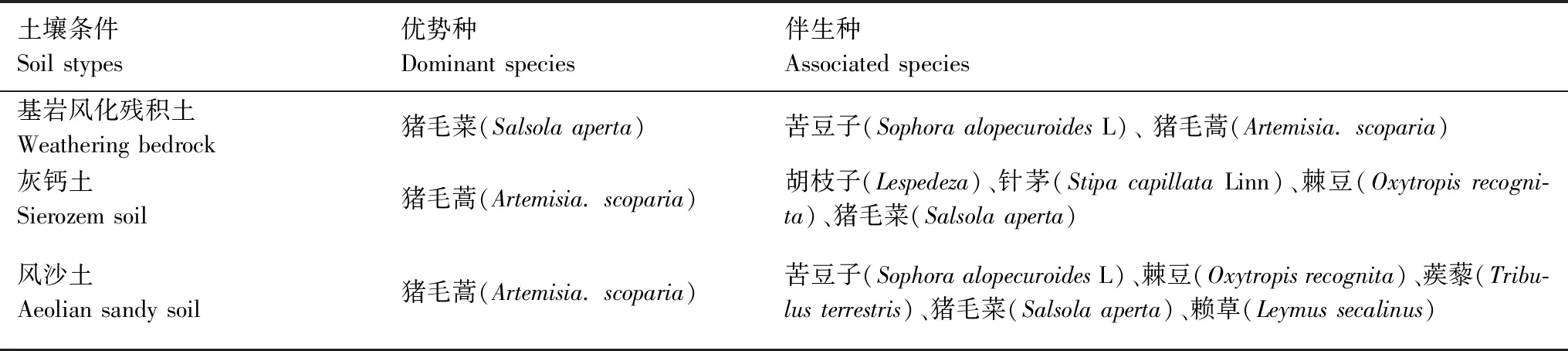
表1 实验样地基本情况
王鑫厅等发现采用摄影定位法测得的值和实测值之间无显著性差异,这种方法是可行、可靠的。因此,本实验参照王鑫厅等人的实验方法于2017年5月份采用摄影定位法测定了上述3个样地内猪毛蒿幼苗种群空间格局,其具体操作过程如下:在各个样地中,将选择的10 m×10 m的群落片段,用白色pvc细管标记,将其分割成400个50 cm×50 cm的亚样方,仔细的将亚样方内的枯落物和立枯物去除。把Nikon D5300数码相机镜头竖直向下尽量保持在离地面1m的位置,从左端第一个亚样方开始按照从左到右,从上到下的顺序依次对每个亚样方进行拍照,将照片按照1—400的顺序进行编号并输入到计算机中。 在Photoshop7.0图像处理软件中对照片进行裁切并拼接成完整的群落片段图。之后在ArcGIS 10.3中对每个样地的群落片段图进行矢量化处理,得到样地内猪毛蒿个体的坐标,以便后续进行数据分析。
1.3 分析方法
1.3.1点格局分析
本研究运用成对相关函数g(r)进行猪毛蒿种群幼苗的点格局分析,g(r)源于Ripley′sK函数,但克服了Ripley′sK函数的累积效应,二者存在g(r)=(2πr)-1dK(r)/dr的相互关系。在点格局分析中,通过Monte-Carlo方法绘制置信区间来判定点格局的分布状况,如果在包迹线以上,则符合聚集分布;在包迹线内,则符合随机分布;在包迹线以下,则符合均匀分布。本研究中选择g(r)函数,Monte-Carlo拟合99次,置信水平为99%,置信区间通过使用最大值和最小值获得,步长为0.1 m[33]。
选择合适的零模型是解决生态学问题的关键所在,因此本研究选择完全空间随机模型、泊松聚块模型和和嵌套双聚块模型进行对比,可以更好的揭示不同土壤类型下猪毛蒿种群空间分布格局的分布特征。
1)完全空间随机模型
完全空间随机模型其实质是均质泊松过程,常作为零假设检验物种的分布格局,也是零模型中最简单的一种模型。在这个模型中任何一点(或个体)在研究区域内任何一个位置出现(或发生)的机会是相同的;同时,点间(或个体间)是相互独立的,任何两点间(个体)均不发生相互作用[20]。
2)泊松聚块模型
泊松聚块模型其实质是均质托马斯过程,它描述的是一种聚块机制,泊松聚块模型的K函数与g函数的表达式如下:
其中,r为尺度,ρ为该过程中母体的密度,σ2为高斯分布的方差。
3)嵌套双聚块模型
嵌套双聚块模型[3]是泊松聚块模型的多代扩展,泊松聚块过程的子代产生自己的后代个体,嵌套双聚块模型的K函数与g函数表达式如下:
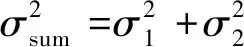
而参数ρ进而σ2意义同泊松聚块模型,其中,下角标1代表第一代,下角标2代表第二代[33]。
1.3.2数据处理
采用ArcGIS 10.3进行数据坐标点采集,数据计算在Programita软件[19]下完成。荒漠草原不同土壤条件下猪毛蒿个体分布的位点见图1,其详细的零模型参数见表1。
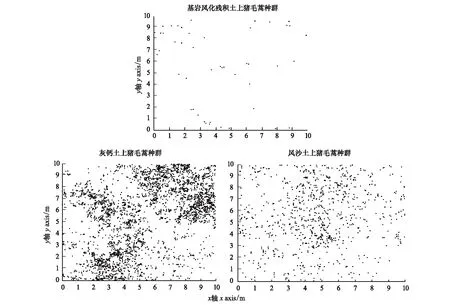
图1 不同土壤条件猪毛蒿种群个体分布位点图Fig.1 Individual distribution locus map of Artemisia capillaris population under different soil conditions a:基岩风化残积土上猪毛蒿种群 Sample a: Population of Artemisia capillaris on weathered residual soil of bedrock;b:灰钙土上猪毛蒿种群 Sample b: Population of Artemisia capillaris on calcareous soil;c:风沙土上猪毛蒿种群 Sample c: Population of Artemisia capillaris on aeolian sandy soil
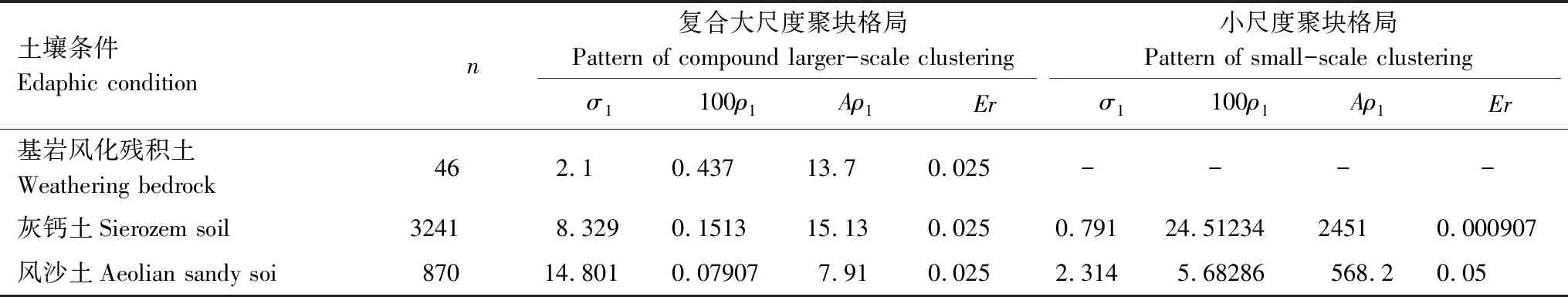
土壤条件Edaphic conditionn复合大尺度聚块格局 Pattern of compound larger-scale clustering小尺度聚块格局 Pattern of small-scale clusteringσ1100ρ1Aρ1Erσ1100ρ1Aρ1Er基岩风化残积土Weathering bedrock462.10.43713.70.025----灰钙土Sierozem soil32418.3290.151315.130.0250.79124.5123424510.000907风沙土Aeolian sandy soi87014.8010.079077.910.0252.3145.68286568.20.05
A:研究区域的面积(10 m×10 m)size of study(10 m×10 m);n:格局中点的数目 number of points of the pattern;σ1和σ2:聚块尺度参数 parameter describing the cluster size;ρ1和ρ2:母体格局的密度 the intensity of the parents pattern;Aρ1:研究区域母体的数量 the number of parents in the plot of sizeA;Er:模型估计的误差(cm)fraction of the total sum of squares of the empirical g and L function not explained by the fit (in centimeter)
2 结果与分析
2.1 不同土壤条件下猪毛蒿幼苗种群密度的比较
在10 m×10 m的取样范围内,灰钙土上猪毛蒿幼苗种群的密度(32.41株/m2)>风沙土上猪毛蒿幼苗种群的密度(8.7株/m2)>基岩风化残积土上猪毛蒿幼苗种群密度(0.46株/m2)。
2.2 基于完全空间随机模型的点格局分析
利用完全空间随机模型分别对基岩风化残积土、灰钙土和风沙土上的猪毛蒿幼苗群落进行点格局分析:由图2看出基岩风化残积土上的猪毛蒿群落在0.15—0.25 m之间表现为聚集分布,在其余的尺度上表现为随机分布;灰钙土上的猪毛蒿群落在0—2.85 m之间表现为聚集分布,在2.85—3.75 m之间表现为随机分布,当尺度>3.75 m表现为均匀分布;风沙土上的猪毛蒿群落在0—1.85 m之间表示为随机分布,在1.85—2.35 m之间表现为随机分布,当尺度>2.35 m表现为均匀分布。
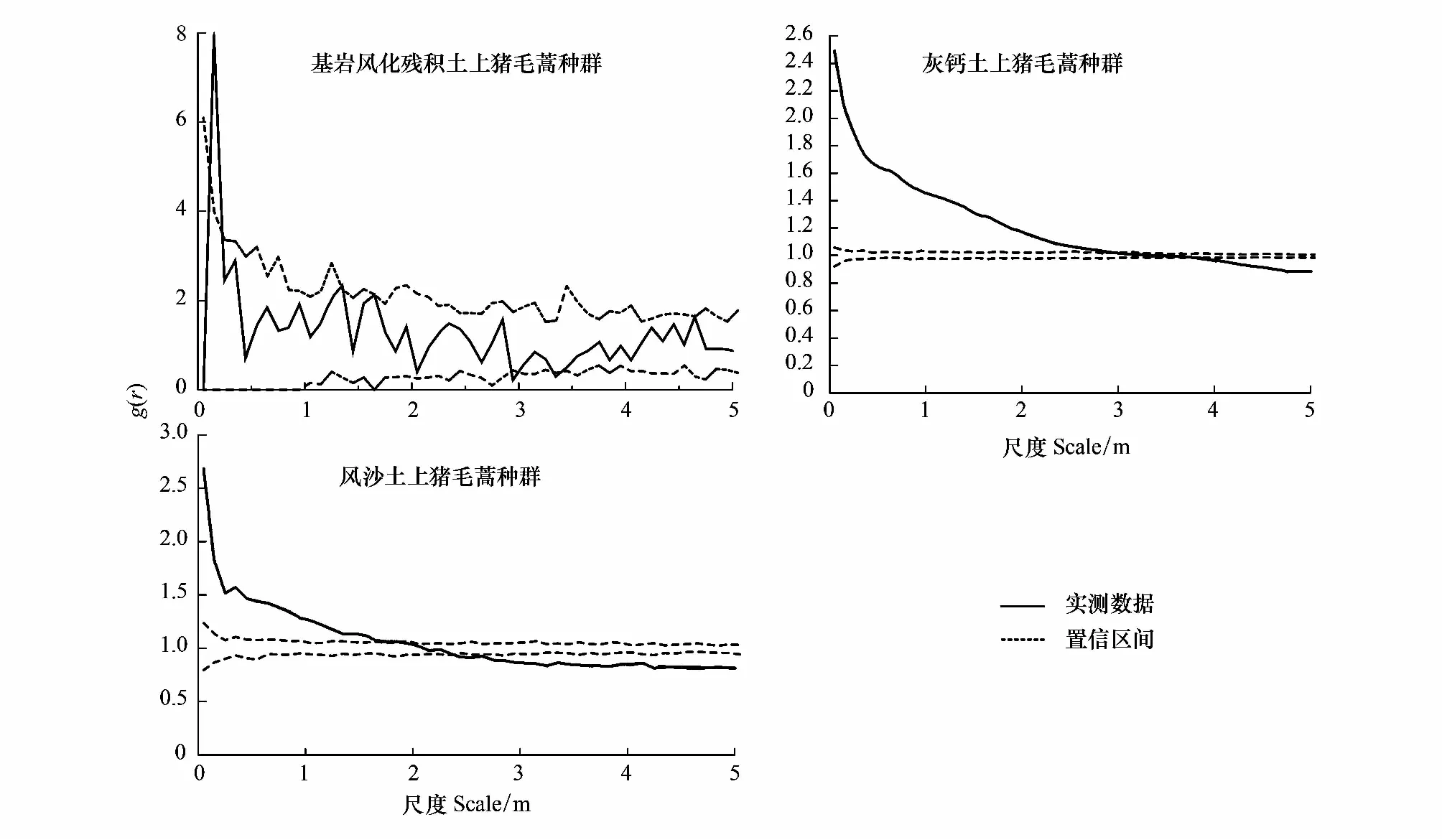
图2 基于完全空间随机模型的点格局分析Fig.2 Point pattern analysis based on complete spatial randomness
2.3 基于泊松聚块模型与嵌套双聚块模型的点格局分析
由图3可以看出基岩风化残积土上的猪毛蒿种群在整个测定尺度上完全符合泊松聚块模型;灰钙土上的猪毛蒿在0—0.05 m之间偏离泊松聚块模型,而在整个尺度上与嵌套双聚块模型相吻合(图4)。风沙土上的猪毛蒿在0—0.1 m和3.9—4.5 m之间偏离泊松分布聚块模型,而在整个尺度上与嵌套双聚块模型相吻合(图4)。
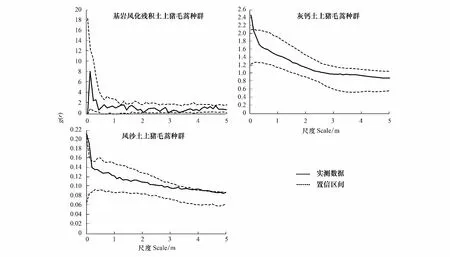
图3 基于泊松聚块模型的点格局分析Fig.3 Point pattern analysis based on Poisson cluster process
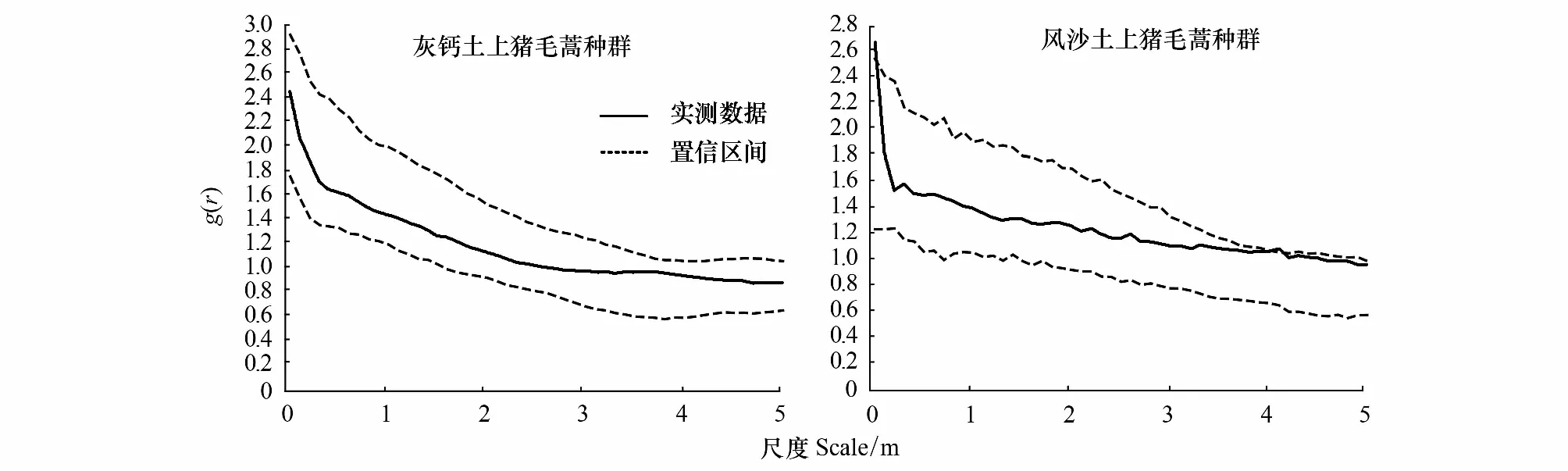
图4 基于嵌套双聚块模型的点格局的分析Fig.4 Point pattern analysis based on nested double-cluster process
2.4 不同不土壤条件下猪毛蒿幼苗种群格局的比较
采用完全空间随机模型对不同土壤条件下猪毛蒿幼苗种群空间格局分析时发现,灰钙土、基岩风化残积土和风沙土上的猪毛蒿幼苗种群在一定尺度上均表现为聚集分布。为了进一步揭示聚集分布的特征,采用泊松聚块模型检验,结果发现基岩风化残积土上的猪毛蒿幼苗种群在整个尺度上位于包迹线内,符合泊松聚块模型(图3)。而灰钙土和风沙土上的猪毛蒿幼苗种群在小尺度范围内在包迹线以上,不符合泊松聚块模型(图3)。进一步采用嵌套双聚块模型对风沙土和灰钙土上的猪毛蒿幼苗种群进行检验,结果显示灰钙土上的猪毛蒿幼苗种群完全符合嵌套双聚块模型(图4)。风沙土上的猪毛蒿幼苗种群除了在0—0.05m上偏离嵌套双聚块模型,在其余尺度完全符合嵌套双聚块模型(图4)。这也就意味着基岩风化残积土上猪毛蒿幼苗种群聚集分布的聚块内不存在较高密度的小聚块,而风沙土和灰钙土上的猪毛蒿幼苗种群存在较高密度的小聚块。可以看出,若实测值偏离完全空间随机模型程度越强则越符合泊松聚块模型,偏离泊松聚块模型程度越强则越辐射嵌套双聚块模型[3]。
3 结论与讨论
(1)基岩风化残积土并不适宜猪毛蒿幼苗个体的定居。在10 m×10 m的取样尺度内,猪毛蒿个体仅仅只有46株,密度为0.46/m2。自然条件下,植物通过繁殖在母体周围产生新个体而呈现聚集分布[21]。我们利用完全空间随机模型的检验结果得出,在基岩风化残积土上猪毛蒿幼苗种群几乎在整个尺度上呈现随机分布,也就是猪毛蒿幼苗个体之间是相互独立的,没有任何作用。但在自然条件下,植物通过繁殖在母体周围产生新个体而呈现聚集分布[21]。可见,存在于基岩风化残积土上的猪毛蒿幼苗并不是由原生境猪毛蒿母体繁殖而来。由于猪毛蒿种子为瘦果,体积小,重量轻[34],容易随风飘散[35],其他生境上的猪毛蒿母株进行种子传播时受到风力的作用将种子散落在基岩风化残积土上,在某些微生境上得以萌发以致出现随机分布。
(2)植物种群的空间分布格局是植物自身的生物特性(扩散机制、生物相互作用、克隆繁殖生长)与生境异质性共同作用的结果[36]。在小尺度上,种群内部关系是决定植物种群空间分布的最为重要的因素。若种群内部之间具有正向的生态关系,那么就容易呈现聚集分布特征;若种群内部之间具有负向的生态关系,那么就容易呈现聚集分布特征;若种间内部之间没有明显关系,那么就容易呈现随机分布特征[37]。种群的分布格局也随着空间的尺度不同而变化。我们用完全随机模型发现在一定的尺度范围内,灰钙土和风沙土上猪毛蒿幼苗种群在小尺度上均呈现聚集分布,随着尺度的增大先呈现出随机分布最后呈现均匀分布。不同学者对不同植物种群的点格局的分析也得出了类似的结果。例如,樊登星等[38]对北京山区栓皮栎林优势种群点格局分析结果中发现,栓皮栎种群在研究尺度0—39 m的范围内呈现聚集分布,当尺度大于39m时呈现随机分布,当尺度大于50m时呈现均匀分布。李尝君等[39]对艾比湖湿地自然保护区克隆植物群落空间格局分析的结果显示,罗布麻在0—1 m尺度上近似随机分布,在1—4.7 m尺度上聚集分布,在4.7—5 m尺度上均匀分布。赵成章等[40]对高寒退化草地甘肃臭草种群分布格局的分析结果中发现,甘肃臭草在斑块形成、扩散、稳定和衰退阶段亦有上述现象。大多数学者认为格局转变的驱动因素是自疏或他疏过程中引起的种群密度的降低[41]。小尺度的聚集分布有利于物种之间弱化种间竞争,增加对恶劣环境的适应性和抵抗力。随着尺度的增大,由于种间竞争过程中种群自调节机理不同引起聚集分布格局向随机分布和均匀分布转变[38-41]。然而在本研究中选取的为猪毛蒿的幼苗,种内竞争和种间竞争相对弱化,生物学特性和生态学过程都不是这阶段种群空间分布格局的主导因素,那么土壤的异质性就成为了猪毛蒿幼苗种群在完全空间随机模型下所呈现不同分布特征的主要驱动因素。猪毛蒿幼苗种群在不同土壤类型下以聚集、随机和均匀分布的形式来响应土壤资源斑块分布的异质性。土壤的异质性能够制约着植物种子萌发,幼苗生长机会的可利用性,促使植物在生活史早期具有不同的分布格局[27]。
(3)在一定的尺度范围内,基岩风化残积土上的猪毛蒿幼苗种群完全符合泊松聚块模型,说明在大聚快中不存在较高密度的小聚块。虽然灰钙土和风沙土上的猪毛蒿幼苗种群在一定的尺度范围内符合嵌套双聚块模型,即大聚快中分布着较高密度的小聚块,但灰钙土上猪毛蒿幼苗种群小聚块多余风沙土。有研究称这种格局是种群易化作用的的结果。在环境胁迫的条件下,植物通过表型可塑性实现互助,共同抵御外界不利条件。胁迫梯度假说认为易化作用与环境胁迫的强度成正相互作用,也就意味着胁迫程度越强,大聚块内就存在较多的高密度的小聚块[21-33]。然而灰钙土作为该研究区原生土壤比退化后的风沙土水分和营养资源优越,环境胁迫强度低,但却存在较高密度的小聚块,因此猪毛蒿幼苗风沙土和灰钙土上分布的格局并不是种群易化的结果。我们猜测应该是灰钙土和风沙土上存在更小的生境异质性,且灰钙土上的小的生境异质性大于风沙土。这就说明植物种群的分布类型和聚集强度在不同的生境尺度上对微环境异质性的响应不同。一些学者对不同植物种群分布格局与异质生境的响应研究中也得出了类似的的结论。杨洪晓等[8]对毛乌苏沙地油蒿种群点格局分析结果中表明,在小于24 m的一系列空间尺度上,固定沙地和半固定沙地的油蒿种群主要呈聚集分布,半固定沙地油蒿种群的聚集强度大于固定沙地。高杨等[42]对土壤营养异质性对鹅绒委陵菜分布格局和空间扩展的影响中发现,土壤营养异质性对鹅绒委陵菜种群的格局有一定的影响,鹅绒委陵菜种群空间拓展能力也由于土壤营养的异质性而发生改变。
本研究基于3种不同的零模型对不同土壤类型猪毛蒿幼苗种群进行点格局分析,发现在猪毛蒿种群种内和种间关系相对弱化的生活史初期阶段,不同土壤类型的猪毛蒿幼苗种群具有不同的分布格局,决定不同分布格局的主要因素为土壤的异质性[43]。然而对于分布格局与土壤异质性内在的联系还不清楚,未能揭示格局背后异质生境土壤驱动因子的贡献度。再者在资源异质性的生境中植物个体的生物学特性和生态学过程在整个生活史阶段是动态的,植物对环境的忍受程度也有差异,植物分布格局也是一个动态的过程[44]。因此研究不同生活史阶段的猪毛蒿种群分布格局,将会更深入的认识猪毛蒿种群在荒漠草原的分布特征。
