基于不同光谱变换的剑湖茭草鲜生物量估测研究
2019-10-23余哲修罗恒春牛晓花
张 超 余哲修 黄 田 张 一 罗恒春 牛晓花
( 西南林业大学林学院,云南 昆明 650233)
湿地植被是湿地的重要组成部分之一,对湿地植被生物量的监测和评价,可以衡量湿地生态系统的健康状况,进而深入研究湿地生态系统碳循环、氮循环、净初级生产力等[1-2]。高原湿地是湿地科学领域研究的热点问题之一,位于滇西北的高原分布有许多独特的湿地,其生境复杂、物种多样性丰富,并且生态系统脆弱,人为活动自然因素的影响已造成生态系统破坏和湿地资源丧失,因此对高原湿地植被生物量的演变状况进行及时、准确的监测和评价显得至关重要[3-5]。
传统的湿地植被生物量估算主要靠人工实地布设样方,收割采集,既费时费力,又具有破坏性,并且湿地以水体或沼泽为主,环境较为复杂,部分区域难以实地设置样方采集植被[6-7],利用遥感技术对湿地植被的生物量进行调查估算,能够减少人力物力的耗费,相比传统调查方法,对湿地生态系统的研究能发挥更多的价值。高光谱遥感具有波段数量多、光谱分辨率高、光谱连续等特点,可以获取地物丰富的反射光谱信息[8],对高光谱数据的变换可以减少背景、光照等因素的影响,能够更好地提取地物光谱特征,一些植被的光谱反射率经过不同的数学变换后,可以增强光谱与其生理生化参量的相关性,从而利用高光谱数据定量反演植被的理化参数[9-11]。基于高光谱数据估测湿地植被生物量在国内外已被成熟应用,如Sun等[12]利用多种高光谱植被指数对洪河湿地优势植物地面鲜生物量估测,ATSAVI指数与鲜生物量的相关系数达0.85(P<0.01);Adam等[13]基于随机森林回归,利用高光谱构建的NDVI和EVI对iSimangaliso湿地公园的莎叶草(Cyperus papyrus)生物量进行估测,最高R2为0.90;Luo等[14]结合激光雷达和高光谱数据对张掖国家湿地公园的芦苇(Phragmites australis)生物量进行估测,研究表明,结合LiDAR和高光谱数据估测的精度比单独使用高光谱数据精度提高了16.5%;李伟娜[15]利用CHRIS数据估测了隆宝滩湿地植被生物量,得出基于+36°VInew为自变量建立的指数模型估测鲜生物量效果最佳;卢霞等[16]通过高光谱数据构建了窄波段植被指数,估测了三洋港湿地碱蓬(Suaeda glauca)的地上鲜生物量,得出基于构建的线性回归模型精度最高。
本研究以大理剑湖湿地为研究区,选取剑湖湿地分布最多的茭草(Zizania latifolia)为研究对象,结合实测茭草的生物量和茭草反射光谱数据,通过不同的光谱变换分析,建立光谱变量和鲜生物量的估算模型。旨在探讨利用高光谱反演湿地植被生理生化参数的可行性,从而为湿地生态系统的研究提供参考。
1 研究区概况
剑湖湿地位于云南省大理白族自治州剑川县境内,地处东经 99°55′,北纬 26°28′,总面积约7.5 km2,平均海拔2 186 m,湖面积6.23 km2,平均水深2.7 m,最大水深6.0 m。剑湖湿地植被类型包括:草丛沼泽型群系4个,漂浮植物型、浮叶植物型、沉水植物型群系10个。沉水植被是剑湖湿地植被中分布面积最广、类型最丰富、多样性程度最高的类型。剑湖湿地共有维管植物443种,其中蕨类植物10科14属20种,种子植物102科267属423种,在102科中,裸子植物3科3属4种,被子植物99科264属419种。剑湖湿地植被以茭草的分布面积为最大。
2 材料与方法
2.1 数据采集与处理
以生长健康的茭草为研究对象,选择植被的生长季采样,采样时间为2017年7月27日。现地确定采样点位置时,每个采样点间隔大于200 m,共设置33个样地(图1),528个样方,共采集茭草反射光谱曲线1 320条。
样地大小为1 m×1 m,将整个样地划分为16个0.25 m×0.25 m的小样方,每个样地选取3个样方实测光谱,每个样地选取1个样方,采用皆伐法采集样品,采集样本后现场放置于便携式电子天平(精度0.01 g)称量。
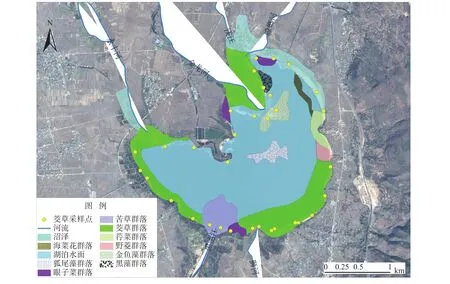
图1 研究区位置及采样点分布Fig. 1 Sketch map of location of study area and sample points distribution
利用ASD Field Spec 3野外光谱仪采集光谱数据(波长为350~2 500 nm)。测量在晴朗、无风、少云条件下进行,测量时间10:00—15:00。光谱采样时,将茭草叶片展平,仪器探头垂直于叶片,每个样本测量30次光谱,视实际环境随时对仪器定标。测量光谱时,受空气水汽、太阳光强度、仪器本身等影响,光谱曲线部分谱段存在噪声和毛刺,使用ASD光谱仪配套的ViewSpec Pro软件将测量误差较大及不理想的曲线剔除;在筛选数据之后对剩余数据进行平滑和去噪处理,并分别对每个样点测量的光谱数据求平均光谱;剔除噪声集中的1 350~1 400、1 750~1 950 nm和2 350~2 500 nm 3个波段范围;去噪后对光谱进行平滑处理,使用Savitzky-Golay法对光谱曲线平滑[17]。
2.2 光谱变换方法
光谱变换后可以降低背景环境、光照和大气散射等对光谱的影响,以凸显植被生化参数对光谱吸收带来的差异。本研究中进行了24种光谱变换,见表1。
包络线去除能有效增强波段的光谱吸收和反射特征,有利于提取特征波段[18]。对数变换可以增强可见光波段的差异,还有利于降低光照变化引起的乘性因素影响[19-20]。倒数变换有利于增强光谱在可见光波段的差异[21]。导数变换可以去除部分背景、噪声等对光谱的影响,在一定程度上还能凸显不同物质光谱特征的细微差别[22-24]。
连续小波变换能够提取植被反射光谱局部吸收特征信息[25-26],不同尺度的变换有助于提取各植物的细节信息和全局信息[27]。选择合适的小波基函数和变换尺度可使其具有凸显光谱曲线局部吸收和反射特征的能力,有利于光谱和植被理化参数的分析[28]。本研究选取 MexicanHat 小波基函数,其在时域和频率都有很好的局部化[29],并在8 个尺度(21、22、23、24、25、26、27、28)下对光谱曲线进行分析。
2.3 模型构建方法
分析各种光谱反射率变换值与生物量的相关性,筛选出与生物量相关性高的变量建立其估算模型。在所有样本中,随机选择80%用于建模,其余20%作为检验数据。分别用单变量回归和多元线性逐步回归建立模型,常用的单变量拟合模型见表2。多元逐步回归分析将每个自变量逐个引入模型,最终保留对因变量解释显著的自变量。

表1 光谱变换方法及表达式Table 1 The methods and expressions of spectral transformation

表2 单变量拟合模型Table 2 Single variable fitting models
2.4 模型检验
采用决定系数(R2)、均方根误差(RMSE)、预估精度(P)和相对分析误差(RPD)评价模型精度。模型决定系数是衡量建模效果优劣的常用指标,是衡量模型所推算的估算值与真实值间的差异化程度,决定系数在0 ~ 1之间,值越大模型预测精度越高。均方根误差可以反映估算数值偏离实测值的程度,RMSE值越小模型的精度越高。预估精度能反映出模型的平均预估能力。相对分析误差可以用来检验估算模型的稳定性及预测能力。RPD>2时,表明模型估测效果较好;1.4<RPD<2时,表明模型估测效果一般;RPD<1.4时,表明模型不能用于估测[30-31]。计算方法分别见式(1)~(4)。

式中:yi为第i个样本的实测值,为第i个样本的估算值,为所有样本实测值的平均值,n为样本总数,SD为样本标准差。
3 结果与分析
3.1 光谱特征分析
3.1.1 简单数学变换后的高光谱特征
经过9种简单数学变换后的光谱曲线见图2。由图2可知,9种光谱变换对茭草的光谱特征影响不同,变换前后的光谱曲线主要特征均表现在可见光和近红外波段。包络线去除变换后,可见光波段中叶绿素对绿光反射和红光吸收的特征波段被明显放大。平方根、立方根变换后反射率相比变换前有所增加,对数变换后的曲线反射率均变为负值,但3类变换曲线形态趋势变化较小。对数倒数、倒数对数、倒数平方根和倒数立方根变换后,曲线形态与变换前的曲线对称,对数倒数变换后相比变换前更容易分析茭草反射光谱的特征波段。倒数变换后551 nm处的“绿峰”和671 nm处的“红谷”特征被凸显出来,蓝光波段367 nm处的反射峰、短波红外的1 449 nm和2 207 nm水汽吸收和反射相比变换前明显突出,说明经过变换后的光谱更容易分析茭草光谱特征。
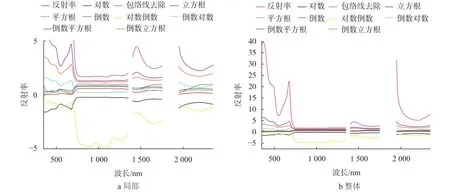
图1 不同形式的光谱反射率简单数学变换Fig. 2 The different simple mathematical transformation of spectral reflectance
3.1.2 导数变换后的高光谱特征
经过7种一阶导数和7种二阶导数变换后的光谱曲线见图3~4。由图3可知,茭草反射光谱经过导数变换后,相比简单数学变换可以更明显地表现出反射光谱特征。一阶微分的光谱可以表现出植被的“三边”参数特征,经过对数一阶微分、倒数一阶微分、对数倒数一阶微分和倒数对数一阶微分变换后曲线波动起伏变化较大,蓝边522 nm、黄边571 nm和红边718 nm相比一阶微分变换表现地更加明显,而平方根一阶微分和立方根一阶微分变换的曲线形态和趋势变化幅度较小,不能很好地突出可见光波段的光谱特征。在近红外波段范围,对数倒数一阶微分相比其他变换,增强了760、850、910、960 nm和1 120 nm附近水和氧气窄吸收带的特征,近红外波段的3个波峰和2个波谷能较好地表现出来。在1 400~1 750 nm的短波红外波段,1 450 nm附近为含水分和二氧化碳的强吸收区域,在变换前的光谱曲线形态上表现为波谷,1 650 nm附近与植物所含水分的波谱特性有关,因此在1 650 nm附近表现为波峰,倒数一阶微分、对数倒数一阶微分和倒数对数一阶微分3种变换突出了从波谷到波峰变化速率最快的1 500 nm附近的波段。
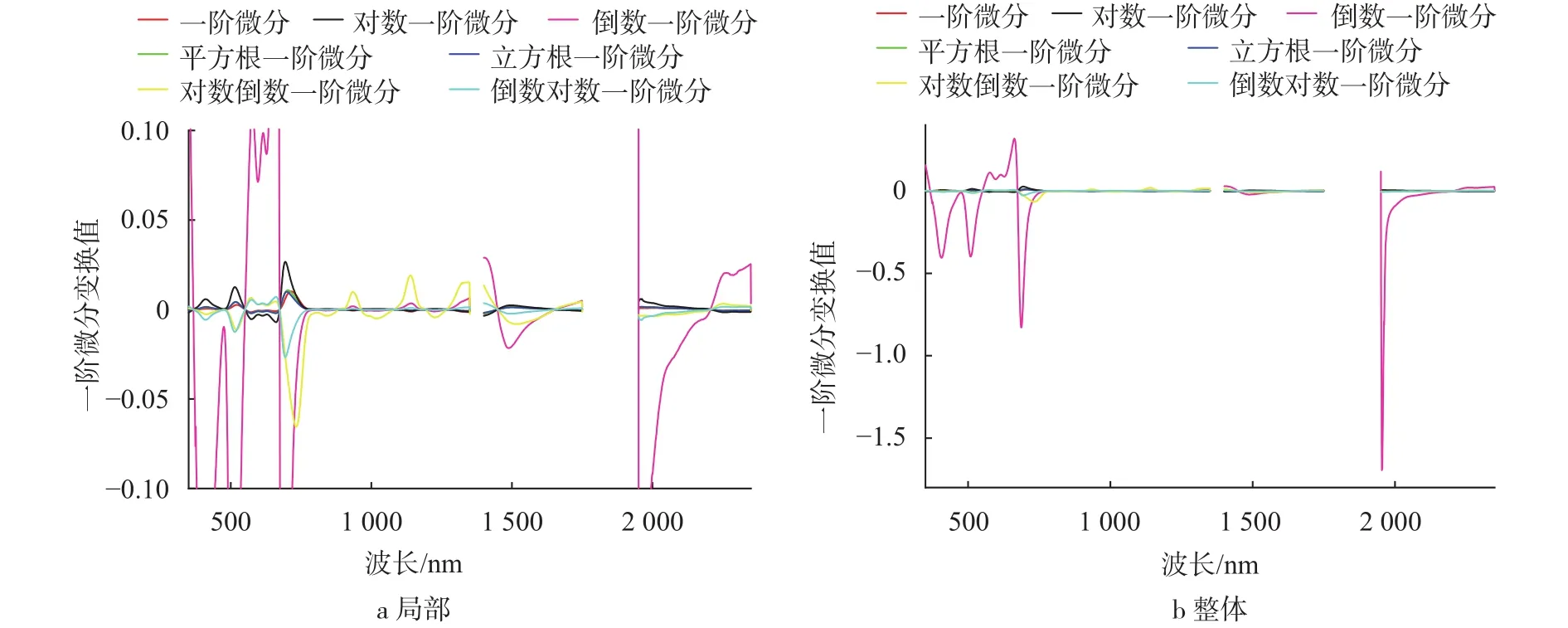
图3 不同形式的光谱反射率一阶导数变换Fig. 3 The first derivative transformation of spectral reflectance in different forms
图4 为7种二阶微分变换的曲线,光谱变换前1 950~2 350 nm波段主要的光谱特征由叶片细胞液、细胞膜和水分的影响形成,相比一阶微分的波峰和波谷更多,在简单数学变换和一阶微分变换后,1 950~2 350 nm的短波红外波段特征没有很好的表现出来,经过倒数二阶微分和对数倒数二阶微分变换后凸显出来,主要表现在2 060~2 170 nm和2 180~2 270 nm附近的范围。
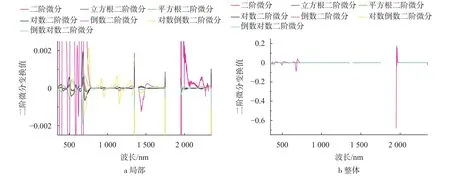
图4 不同形式的光谱反射率二阶导数变换Fig. 4 The second derivative transformation of spectral reflectance in different forms
3.1.3 小波变换后的高光谱特征
经过连续小波变换后的光谱数据见图5。由图5可知,原始光谱在8个尺度下分解,随着分解尺度地增加,茭草的光谱曲线特征逐渐减弱。第1尺度下几乎未表现出明显的特征;第2~3尺度下表现出了“红谷”的吸收特征;第4~6尺度下表现出了“绿峰”的反射特征、“红谷”的吸收特征、近红外波段的吸收谷和多重反射特征,以及短波红外的水汽吸收特征;第7~8尺度下光谱数据逐渐失真,只能表现出茭草光谱的整体特征,而细节特征已经丢失。从光谱分析方面来看,综合比较8个尺度下的光谱特征,中等尺度的连续小波变换既能表现茭草光谱整体特征,同时又不丢失细节特征,第4~5尺度下的连续小波变换较适合分析原始光谱信号。从小波变换后的光谱值来看,分解尺度越高光谱值的绝对值越大,从波长来看,光谱值较大的区域集中于近红外波段,负值集中于可见光和短波红外波段的水汽吸收部分。整体来看,较大和较小的小波系数代表了光谱低频特征,说明可见光和近红外波段对低频特征的响应明显。
3.2 生物量与变换光谱的相关性分析
通过光谱变换数据和生物量之间的相关性分析(图6~7),可以筛选出与生物量相关性强的特征波段,构建茭草生物量估算模型。由图6可知,反射率、对数、倒数、平方根、立方根、对数倒数、倒数对数、倒数平方根和倒数立方根没有任何波段的反射率值与生物量达到显著相关水平。从曲线起伏形态上分析,导数变换相比简单数学变换波峰波谷明显增加,并且正负值交替变动较大,而二阶微分相比一阶微分起伏变化更强烈。从相关系数水平上看,在9种简单数学变换中,仅包络线去除变换有43个波段的反射率与生物量达到显著相关水平(P<0.05)。在14种导数变换中,平方根二阶变换后达到显著水平的波段最多,共有191个波段的反射率值与生物量达到显著相关水平(P<0.05),二阶微分变换达到极显著相关水平(P<0.01)的波段最多,共52个波段;相关性最高的波段为二阶微分变换的1 615 nm,相关系数为-0.730。
由图7可知,在所有变换中,小波变换后达到显著相关水平的波段最多,1~7分解尺度下共有1 097个波段,其中179个波段达到极显著相关水平(P<0.01)。第1尺度变换下共172个波段达到显著相关水平(P<0.05),55个波段达到极显著相关水平(P<0.01),相关性最高的波段为第2尺度(1 617 nm),相关系数为0.734。说明小波变换有利于筛选对生物量敏感的波段,并且能够提高反射率与生物量之间的相关性。
3.3 生物量估算模型分析
通过不同形式变换的光谱反射率与生物量的相关性分析,保留相关性达到显著水平的光谱变量用于建立估算模型,分别用包络线去除、一阶微分、二阶微分、对数一阶微分、对数二阶微分、倒数一阶微分、平方根一阶微分、平方根二阶微分、立方根一阶微分、立方根二阶微分、对数倒数一阶微分、对数倒数二阶微分、倒数对数一阶微分、倒数对数二阶微分和小波变换后的光谱特征波段为自变量。以生物量为因变量,建立茭草的生物量估算模型,拟合结果见表3。由表3可知,基于包络线去除、对数一阶微分、倒数一阶微分、倒数二阶微分、立方根一阶微分和倒数对数一阶微分的估算模型不能通过多元逐步线性回归方法建立,仅能通过单变量回归方法建立估算模型,说明这6种形式的光谱变换对生物量敏感的波段数较少。
模型精度见表4。由表4可知,基于立方根二阶微分变换的估算模型精度最高,R2>0.8、P>80%、RPD在2.0以上,说明立方根二阶微分变换后的特征波段构建的回归模型适用于茭草鲜生物量估测;一阶微分、平方根一阶微分、对数倒数一阶微分虽然估测精度接近于70%,但R2<0.5、RPD<1.4,表明其不适用于鲜生物量的估算,其他单变量回归模型相比一阶微分、平方根一阶微分、对数倒数一阶微分估测效果更差,说明单变量回归模型不能对茭草鲜生物量进行估测。综合分析R2>0.8的模型,基于立方根二阶微分变换的多元回归模型对茭草鲜生物量估测效果最佳,其次为倒数对数二阶微分、对数二阶微分和小波变换,同时可说明小波变换虽然能提高波段对鲜生物量的相关性,但变换后仍然有部分光谱细节特征受到影响,因此基于连续小波变换的回归模型估算效果略弱于基于二阶导数变换的回归模型。
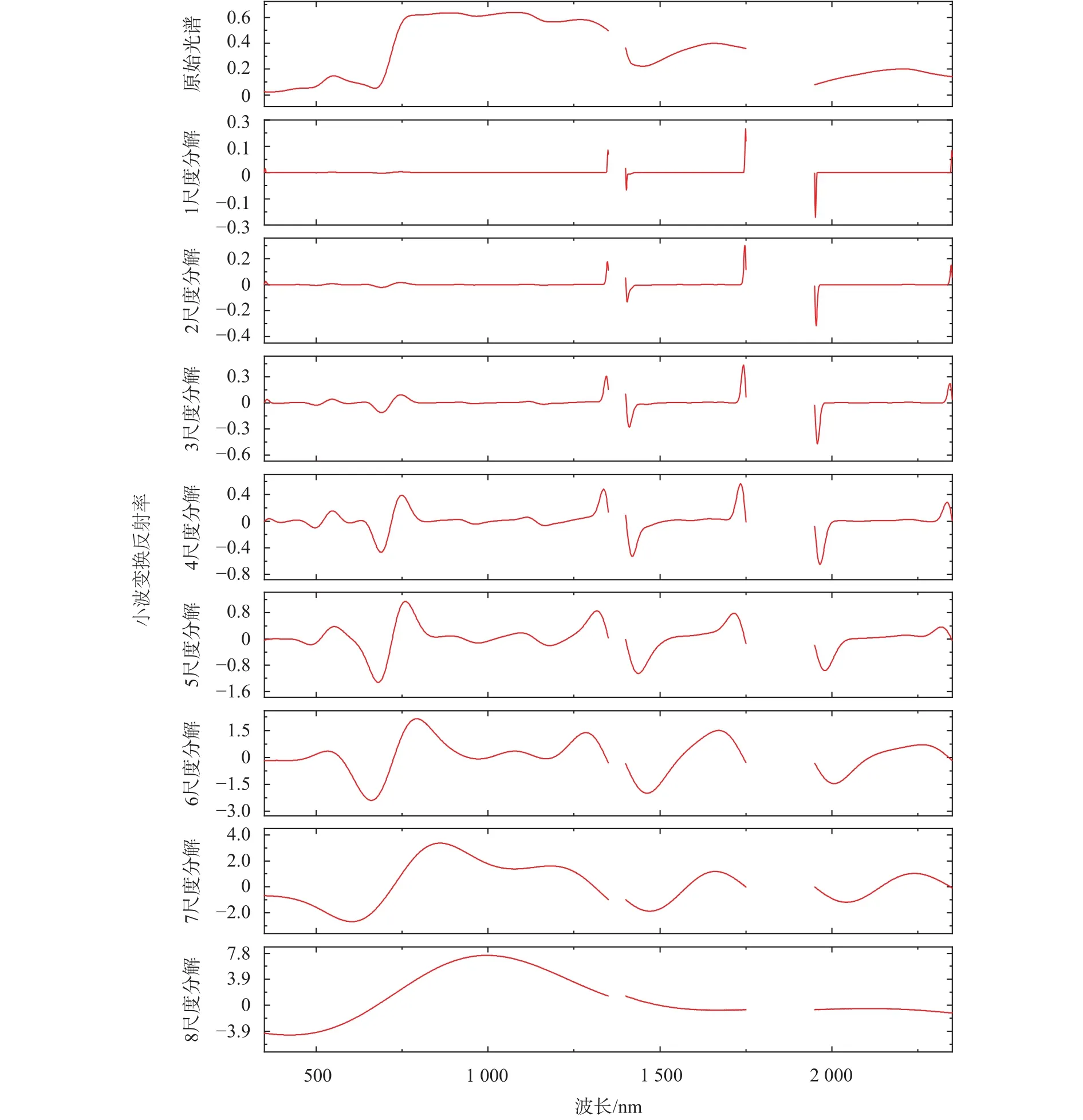
图5 不同尺度的光谱反射率连续小波变换Fig. 5 Continuous wavelet transformation of spectral reflectance under the different scales

图6 简单数学变换和导数光谱值与茭草生物量的相关性分析Fig. 6 Correlation analysis of simple mathematical transformation and derivative spectral value with biomass of Z. latifolia
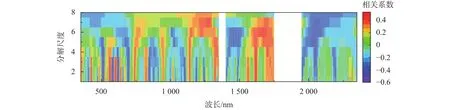
图7 小波变换光谱值与与茭草生物量的相关性分析Fig. 7 Correlation analysis between wavelet transform spectral value and biomass of Z. latifolia

表3 不同光谱变换下的茭草生物量回归模型Table 3 Regression models of Z. latifolia biomass under different spectral transformation
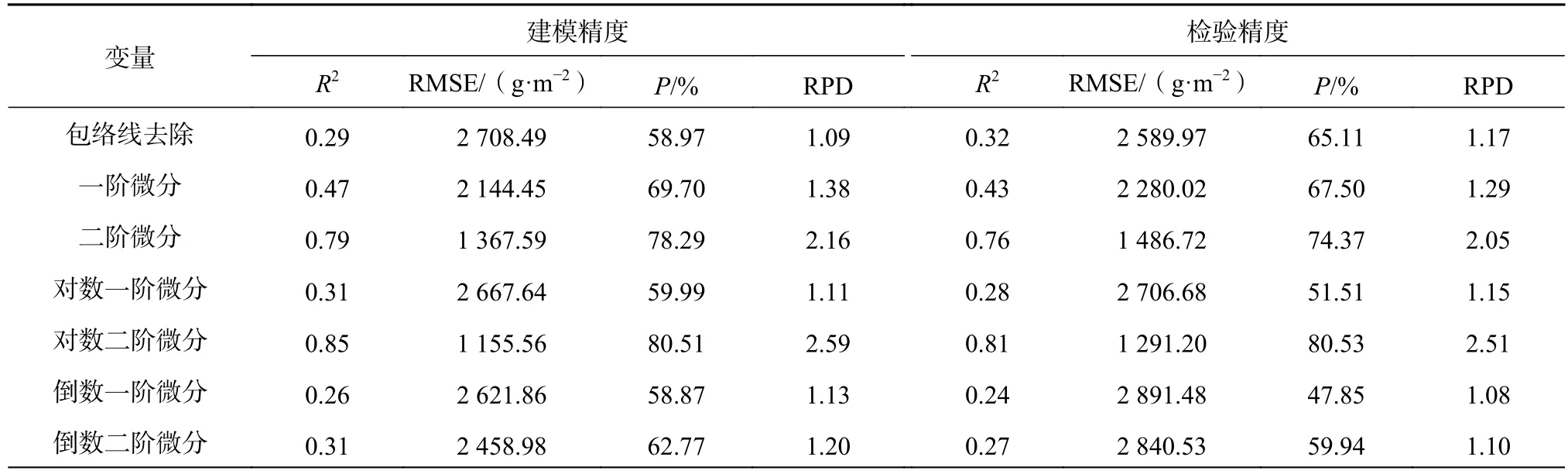
表4 模型精度评价Table 4 Evaluation of model accuracy
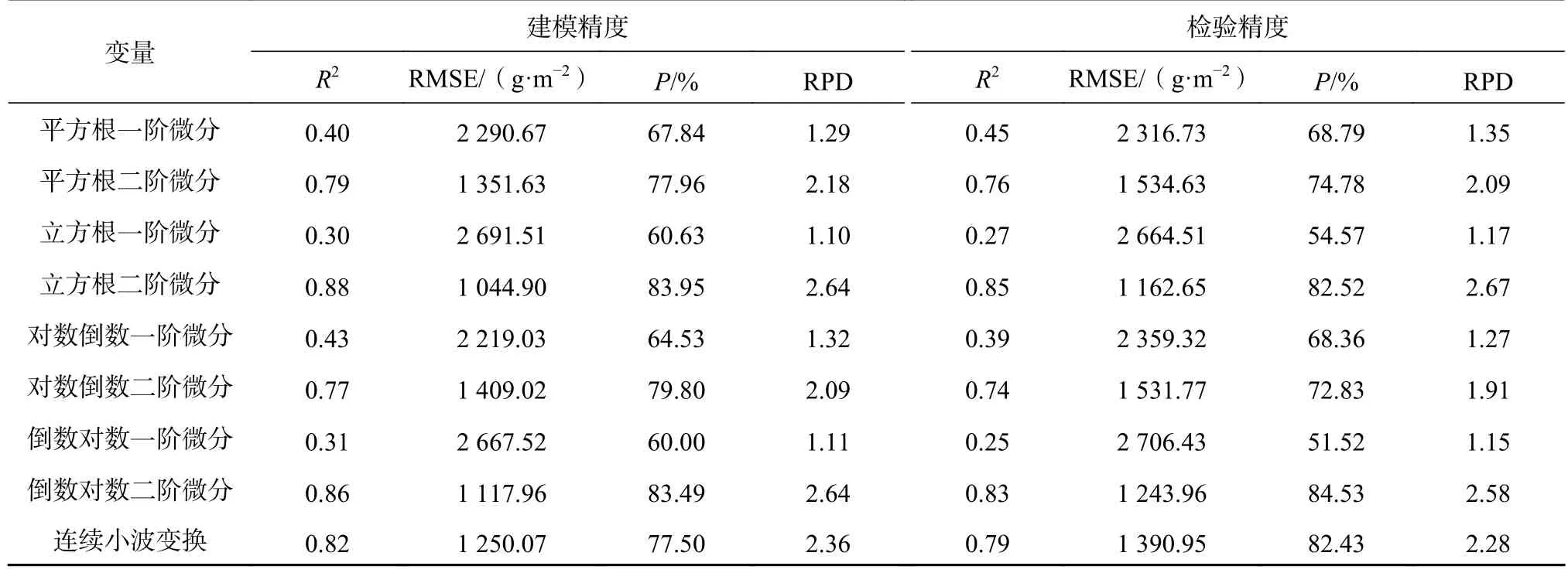
续表4
4 结论与讨论
通过在剑湖湿地实测茭草反射光谱,采集茭草样本,并进行现场称量,对平滑处理后的茭草光谱曲线进行了24种反射率变换。分析了各种变换与茭草鲜生物量的相关性,建立茭草鲜生物量估算模型,得出结论如下:
1)经过不同形式的光谱变换后茭草的反射光谱特征更容易分析,特征波段能够更明显反映出来。对数倒数变换和倒数变换增强了可见光波段的特征;对数倒数一阶微分变换增强了近红外波段的特征;倒数二阶微分和对数倒数二阶微分增强了短波红外的特征;4~5尺度的连续小波变换较适合分析原始光谱特征,既分析了整体特征,又保留了细节特征。
2)与生物量的相关性分析中,连续小波变换达到显著相关水平的波段最多,8个分解尺度下共有1 097个波段,相关性最高的为(R1617,S2),相关系数为0.734;简单数学变换和导数变换中,二阶微分变换达到极显著相关水平的波段最多,共52个波段,相关系数最高为,相关系数为-0.730。
3)从构建的16个鲜生物量估算模型分析,基于立方根二阶微分变换的多元回归模型对茭草鲜生物量估测效果最佳,R2、RMSE、P和RPD分别为0.88、1 044.90 g/m2、83.95%、2.64。
在未来研究中可以通过波段组合构建植被指数、更高阶的微分变换,或在一般数学变换的基础上进一步利用连续小波变换进行光谱分析和筛选对鲜生物量敏感的波段,在建模方法上采用非参数方法进一步研究探讨茭草生物量的估算。从实地采样方面来看,受现场环境限制和影响,采集的光谱虽然经过预处理后能达到应用要求,但在有条件的情况下应使用室内光源测定反射光谱,不仅环境时间可控,并且在水汽影响的波段有数据,使得光谱数据更完整。本研究通过不同的光谱变换提取与鲜生物量相关性高的光谱参量,参考徐新刚等[32]、付元元等[33]、邓兵等[34]学者的研究,构造植被指数或利用光谱曲线本身的特征参数与植被理化参数之间也有较高的相关性,因此今后可尝试利用植被指数和光谱曲线特征参数进行研究,以期得到可提高茭草鲜生物量估测精度的光谱参量。对于高光谱在湿地植被的理化参数的研究和应用上,多数学者对平原湿地研究较多,而剑湖湿地作为高原湿地,植被生长和湿地环境与平原湿地有所不同,未来还应该对不同生长期的茭草生物量进行研究,从而提高筛选光谱变量、建立估算模型的效果。
