亚精胺对镉胁迫下杞柳生长和生理特性的影响
2019-10-23尹泽润诸葛玉平娄燕宏崔秀敏冯文杰李亚平
尹泽润,诸葛玉平*,娄燕宏*,潘 红,崔秀敏,王 会,冯文杰,樊 平,李亚平
(1.土肥资源高效利用国家工程实验室,山东农业大学资源与环境学院,山东 泰安 271018;2.山东省现代创新发展促进中心,济南 250000;3.滨州市农业农村局,山东 滨州 256600)
我国镉污染农田面积已超过2×105hm2,全国超过三分之一的省区存在土壤镉污染现象[1-2]。据报道,已有多地生产的稻米不符合农产品质量安全标准,成为了“镉米”[3-4]。人体长期过量摄入镉,会严重影响肾和肝的生理功能,影响钙、镁的吸收代谢,导致一系列疾病[5]。近年来,镉污染事件频发,镉污染土壤修复已经迫在眉睫[6]。植物修复因其具有绿色、高效、低廉等优点被广泛应用,利用植物对重金属的吸附、转移、清除等作用实现污染土壤的生态修复。已有研究证明,柳属植物多对镉具有较强的耐受性,其中部分品种对镉的富集能力较强[7-8]。杞柳(Salix integra)是柳属(Salix)灌木,它的枝茎生长能力很强,枝条产量高,尤其是细长而坚韧的柳条,去除掉表皮之后,是一种优良的编织材料,也是我国栽培最广泛的灌木柳,具有显著的经济效益。因此,探讨杞柳修复镉污染土壤的潜力,对于丰富用于植物修复的植物种类具有重要的意义。但是,在镉胁迫下,杞柳生长势必受到镉的抑制作用而影响其修复效果,所以杞柳镉耐受性的强化研究仍有待进一步加强。
亚精胺是广泛存在于动植物体内的一种内源激素,可以提高RNA聚合酶活性,对蛋白质与RNA的合成有一定的促进作用[9],同时可延缓动植物衰老、提高抗氧化酶活性、增强蛋白质表达[10]。当前国内外对于施用外源亚精胺来缓解植物非生物胁迫已有一定的研究。李军等[11]利用外源亚精胺缓解了盐胁迫对黄瓜幼苗光合作用的抑制。王学等[12]通过施用外源亚精胺提高了荇菜对汞胁迫的耐受性。张润花等[13]研究了外源亚精胺对盐胁迫下黄瓜幼苗体内抗氧化酶活性的影响。但有关外源亚精胺能否缓解杞柳镉胁迫的相关研究尚鲜见报道。因此,本试验采用营养液培养的方法,使用扦插培育的柳苗进行试验,研究镉胁迫对杞柳生长与根系构型的影响,确定镉胁迫对杞柳的半抑制浓度,明确施用外源亚精胺对缓解杞柳镉胁迫的作用及其生理机制,为应用杞柳进行镉污染土壤的修复奠定理论基础。
1 材料与方法
1.1 试验材料
供试杞柳(Salix integra)取自山东省临沂市莒南县,其品种经鉴定为大青叶,将其枝条采用扦插的方式于山东农业大学试验站进行繁育保存,然后剪取枝条采用水培扦插的方式于温室进行试验。
1.2 试验设计
本试验于2018年8—9月于山东农业大学温室(31±5℃)进行,采用营养液培养试验,将去掉顶端嫩芽的杞柳枝条剪至15 cm长,放入盛有1/2 Hoagland营养液的水培箱中进行扦插培育。培养至生长出3~5条不定根与叶片嫩芽之后,选取长势一致的柳苗进行试验。将柳苗移至新的水培箱中,以1/2 Hoagland营养液处理作为对照(S0),其余处理使用1/2 Hoagland营养液并加入相应浓度的CdCl2(分析纯),Cd2+处理分别为 S10(10 mg·L-1Cd2+)、S20(20 mg·L-1Cd2+)、S30(30 mg·L-1Cd2+)、S40(40 mg·L-1Cd2+),每个处理3次重复。处理7 d后将植株样品取出,测量生物量,计算相对生长率。
依据镉胁迫下杞柳生物量的相关数据,通过SPSS中的IC50分析,得出镉胁迫对杞柳的半抑制浓度,再选取长势一致的柳苗进行试验。将柳苗移至另一个水培箱中,加入1/2 Hoagland营养液,同时加入CdCl2(分析纯)使Cd2+浓度达到半抑制浓度,以不喷施亚精胺的处理作为对照(C0),亚精胺购自Sigma(货号为S2626),亚精胺处理分别为C01(0.1 mmol·L-1)、C02(0.2 mmol·L-1)、C04(0.4 mmol·L-1)、C08(0.8 mmol·L-1)、C16(1.6 mmol·L-1)。采用叶面喷施的方式进行处理,每盆每日喷施5 mL亚精胺溶液,连续喷施7 d,C0处理喷施清水,每个处理3次重复。处理结束后将植株样品取出检测。
1.3 测定指标与方法
1.3.1 生物量与相对生长率
处理前后分别称质量,并计算相对生长率。
相对生长率=(镉胁迫或亚精胺处理始末生物量差值/镉胁迫或亚精胺处理初始生物量)/(对照处理始末生物量差值/对照处理初始生物量)
1.3.2 根系构型
将根系放入根系扫描仪Epson V700的玻璃槽中,注入1 cm的去离子水使根系充分散开,将扫描获取的图像导入Win RHIZO根系分析系统中进行分析获取数据。
1.3.3 生理指标
在处理完成之后,准确称取0.1 g植株叶片样品置于预冷处理的研钵中,用液氮研磨后,加入9 mL生理盐水,转移到离心管中,4℃离心10 min,收集上清液用于生理指标的测定。采用H2O2含量检测试剂盒(A064-1)、蛋白定量测试盒(A045-2)、超氧阴离子试剂盒(A052)、GR分析试剂盒(A062)进行检测,所有操作均严格按照试剂盒说明书执行。
1.3.4 数据分析
试验所用数据为3次重复的平均值,使用Excel 2010进行做图和数据处理,采用SPSS 17.0软件进行方差分析(多重比较方法为Duncan),比较不同处理间差异的显著性水平(P<0.05)。
2 结果与分析
2.1 镉胁迫对杞柳生长的抑制作用
镉胁迫显著抑制杞枊的生长,4个Cd2+处理(S10、S20、S30、S40)杞柳的相对生长率较 S0分别下降18.29%、38.02%、72.33%、75.38%(图1);S20显著小于S10,同时S30、S40显著低于S10和S20,但S30与S40之间差异不显著。根据结果计算半抑制浓度,得出IC50为24.50 mg·L-1。虽然杞柳生长受到镉胁迫的抑制,但当Cd2+达到较高浓度时(30、40 mg·L-1),杞柳仍具有一定的相对生长率(24.62%~27.67%),表明杞柳对镉胁迫具有较强的耐受性,且在应用于镉污染土壤修复方面潜力较大。
2.2 亚精胺对镉胁迫下杞柳的影响
2.2.1 亚精胺对镉胁迫下杞柳生长的影响
施用亚精胺对杞柳镉胁迫具有一定的缓解作用,5个处理(C01、C02、C04、C08、C16)杞柳的相对生长率均显著大于C0(图2),其增长幅度分别为44.97%、85.48%、102.76%、89.83%、76.78%。C04处理显著大于C01、C02、C08、C16处理,且C08和C16处理显著大于C01处理,而与C02处理间差异未达显著水平;同时,C08处理显著大于C16处理,C02处理显著大于C01处理。由此可知,C04处理(0.4 mmol·L-1)提高镉胁迫下杞柳相对生长率的效果最好,且亚精胺对镉胁迫的缓解效应并不随其浓度增大而增强。
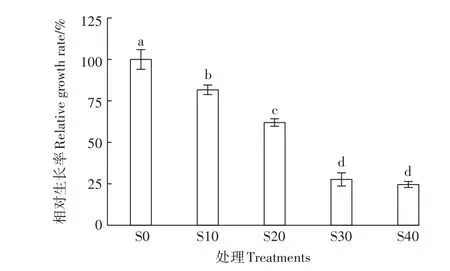
图1 不同镉浓度处理下的杞柳相对生长率Figure 1 Relative growth rate of Salix integra under different cadmium treatments
2.2.2 亚精胺对镉胁迫下杞柳根系构型的影响
施用外源亚精胺可显著缓解镉胁迫对杞柳根系生长的抑制作用,但各根系指标对亚精胺缓解作用的响应浓度存在差异(图3)。杞柳的根长、根系面积、表面积在C01浓度下就对亚精胺产生显著响应(图4),且总根长与面积在C02处理时达到最高;C02、C04、C08处理根长、根系面积显著大于C01和C16,且C16处理根系面积显著大于C01(11.93%)处理;C02、C08、C16处理的根系表面积显著大于C01处理,C02显著大于C16;投影面积、平均直径与根系体积在C02处理浓度即对亚精胺产生显著响应,在C04处理时达到最高(表面积也为最高),且C0与C01间差异不显著;C04、C16分别与C02、C08之间根系投影面积差异不显著,C04显著大于C16,C02、C08间差异不显著。综上可知,亚精胺虽然显著缓解了镉胁迫对杞柳根系生长的抑制,并在C04处理时达到最佳效果,但其缓解作用同样不随施用浓度的进一步提高而增强。
根系构型是表征植物根系生长发育的重要指标,C02、C04、C08、C16处理根系连接数、节点数、分叉数、根尖数显著大于C0、C01处理(表1),且C01处理下节点数、连接数、根尖数显著大于C0处理,分别提高了26.80%、4.88%、5.06%。C02、C04、C08、C16处理的分叉数间均无显著差异;而C04处理的连接数显著大于C02、C08、C16处理;C04、C08、C16处理节点数显著大于C02处理;C02、C04处理根尖数显著大于C16处理。连接数、节点数、根尖数在C01浓度就对亚精胺的缓解产生响应,而分叉数则是在亚精胺浓度达到0.2 mmol·L-1(C02)后显著响应,连接数与根尖数对于高浓度的亚精胺更为敏感,在亚精胺浓度大于0.4 mmol·L-1(C04)后,其缓解效应降低。
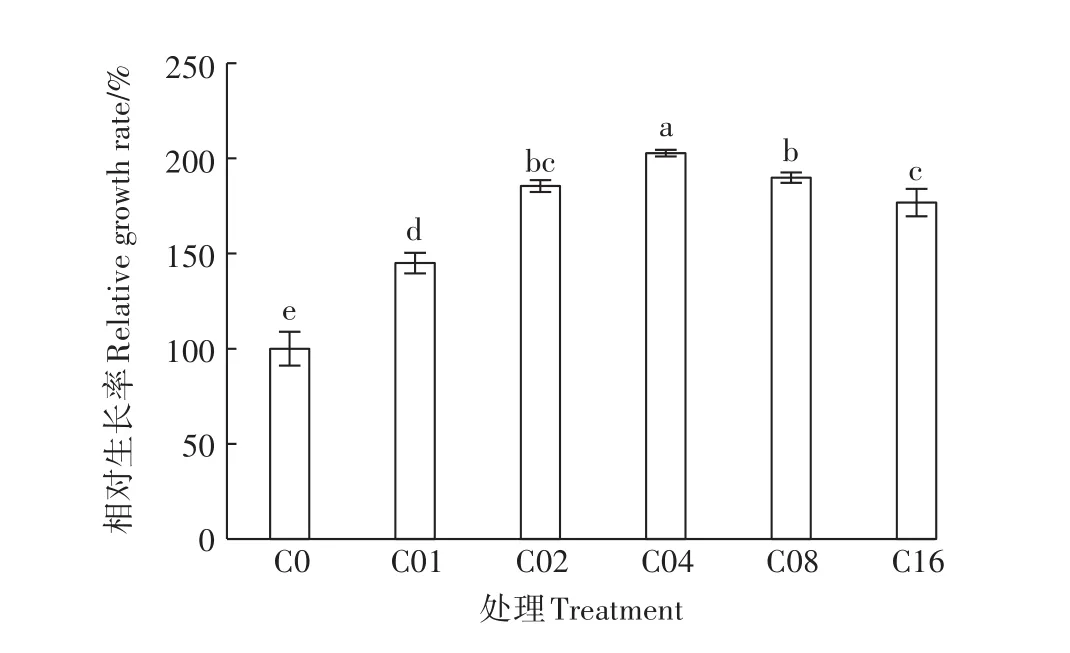
图2 镉胁迫下不同亚精胺处理杞柳相对生长率Figure 2 Relative growth rate of Salix integra treated with different spermidine levels under cadmium stress
图3 镉胁迫下不同亚精胺处理的杞柳根系扫描图Figure 3 Root scanning of Salix integra treated with different spermidine levels under cadmium stress
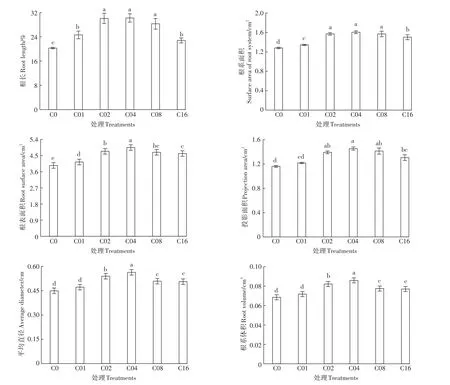
图4 镉胁迫下不同亚精胺处理杞柳的根系生长情况Figure 4 Root growth of Salix integra treated with different spermidine levels under cadmium stress
2.2.3 亚精胺对镉胁迫下杞柳总蛋白、谷胱甘肽还原酶、产生超氧阴离子活力单位和过氧化氢含量的影响
总蛋白是表征植物总代谢和生理活性的重要指标。亚精胺显著提高杞柳总蛋白含量(34.92%~117.46%),且C08、C16处理显著大于C01、C02、C04处理,且C16处理显著大于C08处理(图5)。亚精胺处理显著降低了谷胱甘肽还原酶(GR)、产生超氧阴离子活力单位和过氧化氢(H2O2)含量,C02、C04、C08和C16处理的GR活性显著小于C01处理(降低了47.72%~73.90%),且C02、C04、C08和C16间差异不显著。C08处理产生超氧阴离子活力单位显著小于C01处理(降低了30.90%),而与其他亚精胺处理(C02、C04、C16)差异不显著。C04处理H2O2含量显著小于 C01、C02和 C16处理(降低了 23.38%~28.99%),而与C08处理间差异不显著,且C01、C02、C08和C16间差异不显著。因此,施加亚精胺通过降低过氧化物含量来缓解杞柳镉胁迫,同时促进了蛋白质的合成,显著提高了镉胁迫下杞柳体内的总蛋白含量。
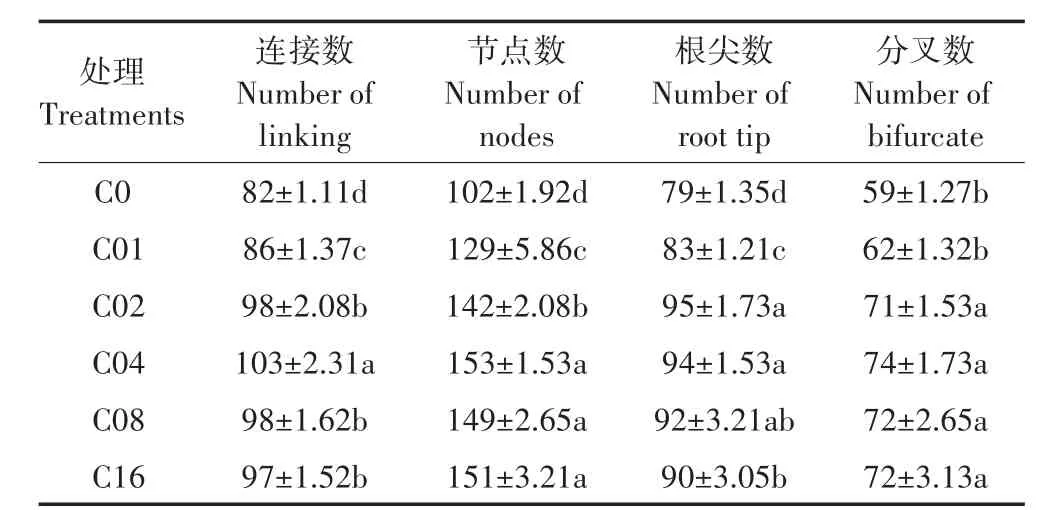
表1 镉胁迫下不同亚精胺处理杞柳根系构型Table 1 Root configuration of Salix integra treated with different spermidine levels under cadmium stress
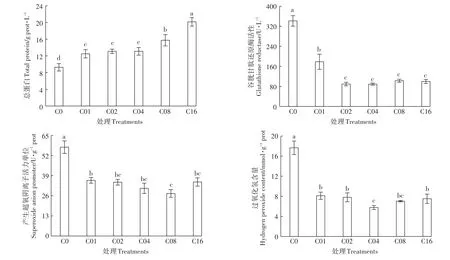
图5 镉胁迫下不同亚精胺处理的杞柳叶片酶活性Figure 5 Enzyme activity in leaves of Salix integra treated with different spermidine levels under cadmium stress
3 讨论
杞柳因生长速率快、生物量高、易繁殖,同时其柳条去除表皮后可作为重要的编织材料,而被广泛种植并应用于工艺品编织。本研究表明,杞柳在40 mg·L-1的镉溶液中仍能存活,其相对生长率为24.62%,表明杞柳应用于镉污染土壤修复具有巨大潜力。但在镉胁迫下,杞柳生长也受到明显的抑制(其相对生长率在24.50 mg·L-1的镉胁迫营养液中下降50%),这又不利于其在镉污染土壤修复中的广泛应用。因此,亟需寻找提高杞柳对镉胁迫耐受性的强化措施。
亚精胺是植物体内最常见的内源激素之一,是植物代谢过程中产生的次类代谢多胺物质。已有研究表明,施用外源亚精胺可提高植物对盐胁迫、Hg2+胁迫的耐受性[13-15]。本试验结果表明,施用外源亚精胺对于缓解杞柳所受镉胁迫也具有十分明显的作用,其相对生长率在0.4 mmol·L-1的亚精胺处理下较对照提高102.76%,显著促进杞柳根系的生长发育,增加其总根长、表面积、体积来提高根系吸收能力,促进杞柳的生长。这与魏国芹等[14]研究报道的植物所受胁迫缓解后的根系变化相一致。
镉胁迫会导致植物抗氧化能力的隐形丧失,影响植物抗氧化酶活性,增加H2O2的内在积累进而导致DNA和蛋白质被破坏[16-17]。本研究发现,在1.6 mmol·L-1的亚精胺处理下,总蛋白含量达到20.15 g prot·L-1(较C0提高了117.46%),这主要是由于亚精胺具有促进蛋白质的合成功能,所以在缓解杞柳所受镉胁迫的同时,随着亚精胺施加浓度的提高而不断提高蛋白质合成速率,导致总蛋白含量持续显著上升[17]。这与杜长霞等[18]研究发现的亚精胺显著提升盐胁迫下黄瓜幼苗总蛋白含量的结果一致。此外,GR作为植物重要的抗氧化酶之一,是通过还原氧化型谷胱甘肽(GS-SG)为谷胱甘肽(GSH)而发挥抗氧化和解毒作用[19-21]。GSH可通过与重金属螯合来缓解植物所受胁迫,当镉胁迫被缓解后,植物体内的GSH与GS-SG重新达到平衡,表明施用亚精胺缓解了杞柳所受镉胁迫,从而使得GR的活性下降。这与张腾国等[21]在逆境胁迫下的GR活性上升结果相印证。同时,在胁迫条件下,植物叶片会产生积累大量的超氧阴离子自由基,H2O2与超氧阴离子过量都会对植株造成组织损伤[22]。本试验发现,0.4 mmol·L-1的亚精胺显著缓解了杞柳镉胁迫,其产生超氧阴离子活力单位与H2O2含量分别下降46.35%和67.39%。这与一氧化氮缓解水稻镧胁迫的研究结果一致,在镧胁迫下水稻叶片中的H2O2含量显著增加,在施用一氧化氮缓解胁迫后,其H2O2含量显著下降[23];田保华等[24]的研究发现镉胁迫下H2O2含量上升。究其原因,H2O2作为一种较为稳定的活性氧,并不会直接造成膜脂的氧化,而超氧阴离子可通过(H2O2+O-2→OH+O2)反应生成OH-对细胞膜脂产生过氧化作用,还会通过特异性点位攻击导致DNA断裂[25],从而对植物产生毒害,而外源亚精胺的喷施通过降低产生超氧阴离子活力单位与H2O2含量,减轻了杞柳所受超氧阴离子与OH的过氧化危害,从而缓解了杞柳所受的镉胁迫。
4 结论
(1)杞柳在镉污染的修复中具有较强的潜力,在40 mg·L-1镉浓度胁迫下仍保持较高的生长速率,其半抑制浓度为24.50 mg·L-1。
(2)施加亚精胺可有效缓解杞柳所受镉胁迫,促进其根系生长发育、提高总蛋白质含量、降低超氧阴离子与过氧化氢含量。其中,施加0.4 mmol·L-1的外源亚精胺对缓解杞柳镉胁迫效果最佳。
