耐草甘膦杂草的研究现状
2019-10-21贾芳崔海兰李香菊于惠林
贾芳 崔海兰 李香菊 于惠林
摘要:草甘膦是目前世界上用量最大、应用范围最广的农药,因为在转基因抗草甘膦作物田中过度依赖其除草,耐草甘膦杂草将演替成优势种群。耐受性杂草不但增加了杂草防除难度和成本,而且还会导致在农田生态系统中因过量使用草甘膦而出现一系列生态风险问题。本文通过对草甘膦特性、耐草甘膦杂草现状和耐受机制等进行较系统的总结和分析,以期为我国未来抗除草剂作物商业化种植后制定杂草治理策略奠定基础,也为草甘膦在转基因作物田高效安全地使用提供理论依据。
关键词:草甘膦;耐受性;杂草;耐受机制;研究现状;展望
中图分类号:S451 文献标志码:A 文章编号:1003-935X(2019)01-0001-09
Abstract:Glyphosate is currently the most widely used herbicide in the world. People depend on glyphosate heavily for glyphosate-resistant crops, which makes glyphosate-resistant weeds the dominant population in the community. Weed population shifts not only increased the difficulty and costs of weed control but also caused a series of ecological risks in agroecosystems because of glyphosate overuse. Through summarizing and analyzing application and action mechanism of glyphosate, and current status and tolerance mechanism of glyphosate-tolerant weeds, this article provides a basis on weed management tactics and appropriate use of herbicides after commercial planting of glyphosate-resistant crops in China in the future.
Key words:glyphosate; tolerance; weeds; tolerance mechanism; research status; prospect
1 草甘膦的特性
草甘膦(glyphosate),学名为N-(膦酰基甲基)甘氨酸,分子式为C3H8NO5P,纯品为非挥发性白色固体;1971年由美国的贝尔德等发现,1974年由孟山都公司开发生产成为内吸、传导、灭生性的茎叶处理除草剂,其商品名为农达、镇草宁。因其在水中溶解性差,之后又推出草甘膦异丙胺盐、草甘膦胺盐、草甘膦钾盐与草甘膦钠盐等[1]。草甘膦能有效防除300多种1年生、2年生与多年生禾本科、莎草科、阔叶杂草、灌木与树木[2]。最初应用于果园和非耕地杂草防除,1996年孟山都公司推出转基因抗草甘膦(glyphosate-resistant,GR)作物后,使得草甘膦可以适用于农田[3]。经过40多年的发展,草甘膦已成为世界上使用量最大的除草剂产品,占全球农药总用量的15%左右,占全球除草剂30%的市场份额,2016年全球销售额为56.6亿美元。而中国是最大的草甘膦生产国,2016年产量为50.5万t[4]。目前我国原药和制剂(含混剂产品)有1 200多个产品已进行登记。
目前,草甘膦商品化产品最常用的是Roundup,其由草甘膦异丙胺盐与表面活性剂和水组成,其中草甘膦异丙胺盐的含量占41%,相当于350 g/L草甘膦。草甘膦对人畜低毒,1993—1994年美国国家环境保护局(EPA)将草甘膦列为毒性级别“E类”,即对人类无致癌性[5],按除草剂抗性行动委员会(HRAC)公布的作用分类方法为G组(有机磷类除草剂)[6]。
草甘膦是唯一以植物叶绿体中5-烯醇式丙酮酰莽草酸-3-磷酸合成酶(EPSPS)为靶标的除草剂。EPSPS在真菌、细菌、藻类和植物体内广泛存在,是芳香族氨基酸(包括色氨酸、酪氨酸、苯丙氨酸)生物合成过程中的一个关键酶。草甘膦竞争磷酸烯醇丙酮酸(PEP)和非竞争磷酸莽草酸(S3P)的方式同植物体内EPSPS进行绑定,形成结构稳定的EPSPS-S3P-草甘膦复合物,终止其催化的PEP与S3P向5-烯醇式丙酮酰莽草酸-3-磷酸(EPSP)的转变过程,从而抑制芳香族氨基酸和一些芳香化合物的生物合成,扰乱植物体的氮代谢[7]。草甘膦接触土壤后与铁、铝等金属离子结合而失去活性,在土壤中的半衰期为44~60 d[8]。草甘膦进入土壤后迅速代谢,降解产物有2种,即C—N健氧化裂解产生氨甲基膦酸(AMPA)、C—P键裂解酶分解产生肌氨酸,大部分草甘膦被微生物分解为AMPA,小部分降解为肌氨酸,2种中间代谢物进一步被微生物降解为磷酸、甘氨酸和二氧化碳[8]。植物体中也发现了类似的草甘膦降解代谢机制,在一些植物中还检测到了AMPA[9]。另外有研究表明,草甘膦还影响植物生理生化过程,如影响光合作用及葉绿素的合成;紊乱植物的碳氮平衡;引起植物出现应激氧化反应和阻碍植物激素及木质素的合成[10]。
2 抗草甘膦作物全球种植面积
转基因抗草甘膦作物的问世不但使草甘膦能够应用在作物田中,还可以减少除草剂用量、降低除草成本、提高除草效果和促进耕作制度变革。自1996年美国孟山都公司推出转基因抗草甘膦大豆后,至2017年全球16个国家种植转基因抗除草剂(herbicide-tolerance,HT)(包括抗虫抗除草剂复合性状,insect resistance/herbicide tolerance,IR/HT)作物,种植面积已达1.66亿hm2,占转基因作物种植总面积(约1.90亿hm2)的87%,其中抗除草剂大豆(包括IR/HT复合性状)面积为 0.94亿hm2,转基因抗除草剂(包括IR/HT复合性状)玉米为0.54亿hm2[11]。20年间转基因抗除草剂作物种植面积持续快速增长。转基因抗除草剂作物可为社会带来巨大的经济收益,就2015年种植转基因抗除草剂大豆使全球农户获利32.9亿美元,1996—2015年的累计收益达381亿美元[12]。
目前,转基因抗除草剂作物种类有大豆、玉米、棉花、油菜、甜菜及苜蓿,种植面较大的3个国家為美国、巴西和阿根廷[11]。抗除草剂基因在转基因大豆、玉米中分别有11、12个,涉及9种除草剂分别为草甘膦、草铵膦、2,4-D、异唑草酮、麦草畏、硝磺草酮、磺酰脲类、咪唑啉酮类和溴苯腈,在大豆、玉米中转化事件为28、201个[13-15]。1996—2017年的22年间,抗除草剂性状始终占据优势地位,但随着复合性状的增加,其优势小幅下降。复合性状作物能节省成本,所以深受种植农户的青睐,就2017年IR/HT复合性状作物在大豆、玉米和棉花中的种植比2016年增长了3%,占全球转基因作物种植面积的41%[11]。随着复合性状转基因作物市场越来越成熟,开发的产品越来越多,在未来的种植面积将持续增长,复合性状、多抗性是转基因作物发展的必然趋势。而在我国现还未允许批准商业化种植转基因抗除草剂作物。
3 耐草甘膦杂草发展现状及耐性机制
3.1 耐草甘膦杂草发展现状
3.1.1 杂草耐受性及耐受杂草种类 杂草耐受性(tolerance)与抗药性(resistance)不同,抗药性是指由于长期、大量使用除草剂或人为诱导、遗传操作,一种植物生物型在对野生型致死剂量处理下,能存活并繁殖的可遗传能力;耐受性是指一种植物天然耐受除草剂处理的可遗传能力,在没有选择或遗传操作条件下,用除草剂处理后能存活、繁殖[16]。杂草在进化过程中,由于多种原因会产生对除草剂不敏感的自然突变体,在除草剂的选择作用下,一些对除草剂敏感的杂草种群被杀死,而那些对除草剂不敏感的自然突变体被保留下来,发展成抗药性种群。耐受性是杂草种群在自然发生状态下便可以忍受除草剂伤害,不需要除草剂的筛选或者说抗性个体数量的积累[17]。同一植物不同种群对同一种除草剂耐受程度存在差异。
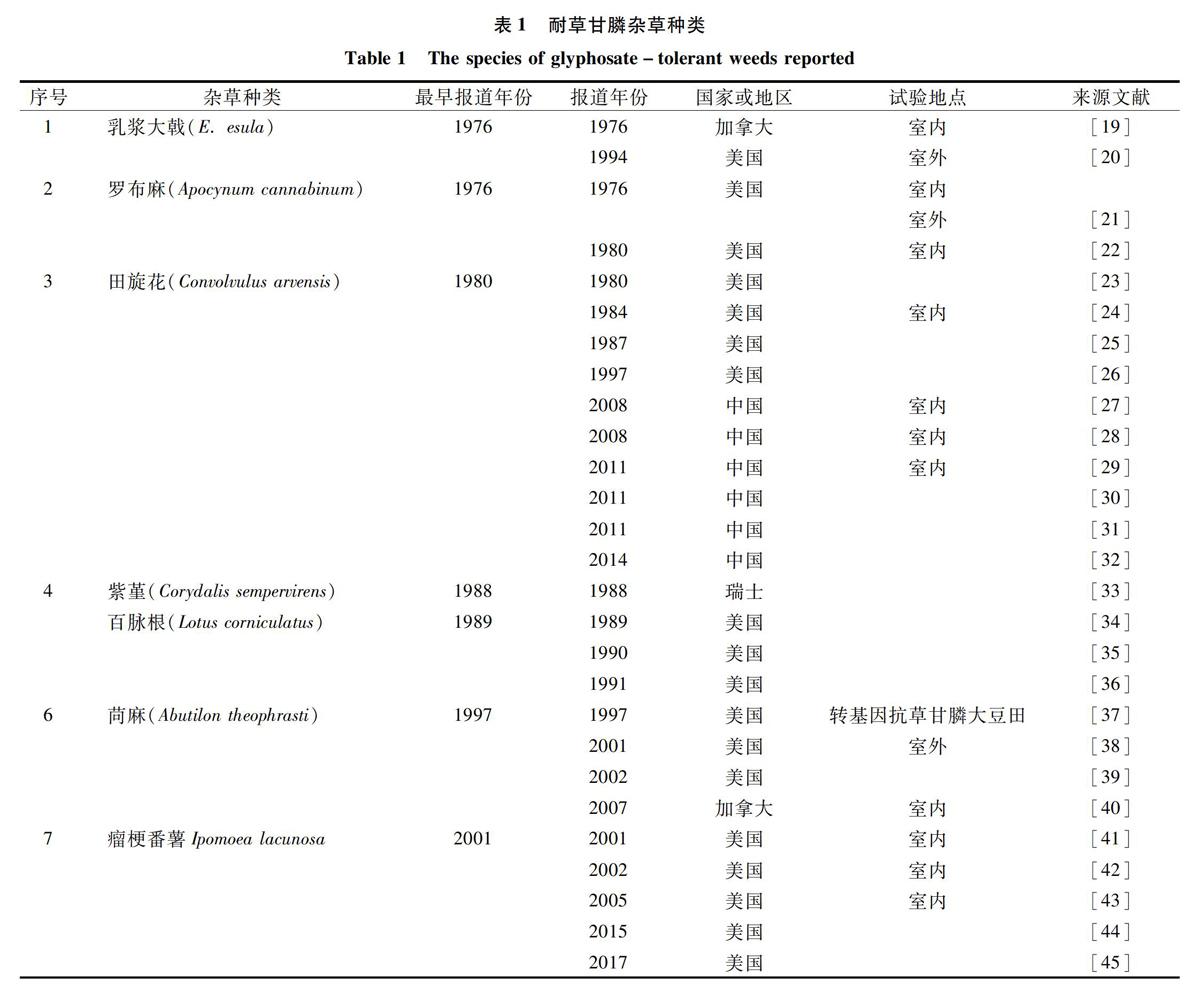
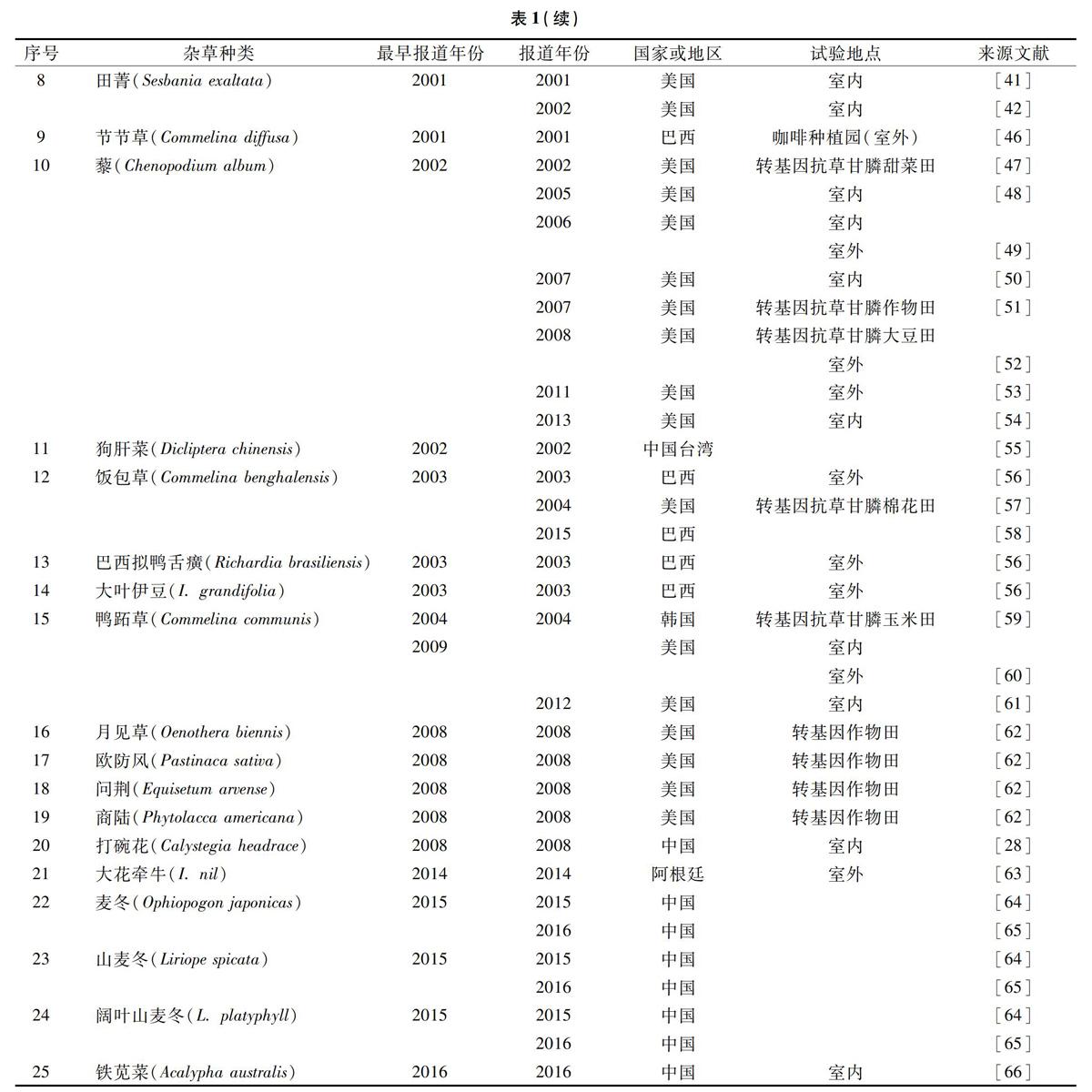
1996年澳大利亚首次发现硬直黑麦草(Lolium rigidum)对草甘膦产生抗性[18],到2019年1月,全球共29个国家发现43种抗草甘膦杂草。草甘膦造成的杂草抗药性问题引起全球各国的广泛关注。1976年Gottrup等报道了对草甘膦有耐受性杂草乳浆大戟(Euphorbia esula),通过与加拿大蓟(Cirsium arvense)对比发现,乳浆大戟对草甘膦有着天然的耐受性,而后的几十年间对草甘膦耐受性的杂草种类相继被发现(表1)。耐受性杂草出现频率相对较低,这是由于敏感性生物型所占比例大,而耐受性生物型所占比例极小。吴加军对江苏地区34科161种杂草进行草甘膦耐受程度测定,并按防治效果将杂草划分为敏感杂草、一般耐受杂草、中度耐受杂草和高度耐受杂草,在秋熟杂草中,耐受杂草只占测试种类的4.4%,高度耐受杂草占测试种类的3.3%[67]。费云燕等研究了南京大豆田中耐性杂草的种类和特性鉴定,发现14种杂草对草甘膦存在耐性,耐性杂草具有粗壮根茎、蜡质叶片等特性[68]。
3.1.2 耐受杂草对杂草群落演替的影响 虽然有报道称在GR作物田中植物和节肢动物种类显著降低,但并没有结论性证据表明GR作物对杂草生物多样性和物种丰富度有直接的影响,主要是由于耕作制度和杂草管理策略改变所带来的巨大选择压间接影响了杂草群落和种群多样性[69]。杂草群落受其特定的生长环境选择压的影响。在GR作物田中,少耕或免耕的耕作制度和单一依赖草甘膦控制杂草策略在很大程度上影响了杂草群落组成和密度,为特定适应的杂草种类增长提供了生态机会,并导致杂草种群的重大变化,以至于不可避免地发生种群的演替[62]。种群演替归咎于杂草对作物生产中使用策略变化的“生态适应”。2003年美国对11个州的GR作物田进行调查,发现在GR棉田和GR大豆田杂草群落从敏感种群向耐受性和抗性杂草种群演替[57]。至于杂草种群演替速度,有报道认为草甘膦耐受性杂草可在种植GR作物后5~8年时间演变成优势种类,也有报道认为杂草种群演替的速度是不确定的[62],但种群演替将带来的结果是增加杂草防除难度和成本,限制了GR作物的广泛应用。
3.2 杂草对草甘膦的耐性机制
相对于杂草对草甘膦的耐受作用机制而言,杂草对草甘膦的抗性机制研究得更为透彻。杂草对草甘膦抗性机制可分为靶标抗性和非靶标抗性。靶标抗性可分为EPSPS发生的突变和拷贝数增加,而非靶标抗性主要包括吸收障碍、转导受阻、屏蔽或隔离作用、氧化代谢和其他解毒代谢[7]。而杂草对草甘膦的耐受性作用机制与此极为类似,其耐受草甘膦的主要原因是EPSPS独特的蛋白结构、EPSPS拷贝数及基因表达量增加,杂草植株有利的形态生理特性和有效代谢及传导。杂草对草甘膦的耐受性也可能同时存在多种耐受性机制[66-67]。
3.2.1 EPSPS独特的蛋白结构 杂草对草甘膦耐受性因其具有独特的EPSPS蛋白结构,导致其对草甘膦天然的不敏感。刘延研究发现,田旋花在靠近EPSPS的保守活性位点101位的氨基酸是极性氨基酸丝氨酸,优先与植物体内的草甘膦结合,使得草甘膦无法竞争性地占据扩增前引物延伸(PEP)结合位点,从而保护活性区域,因此草甘膦不易与田旋花的EPSPS结合,从而使得田旋花对草甘膦具有耐药性[28]。毛婵娟等研究发现,麦冬、土麦冬和阔叶麦冬对草甘膦的耐受性机制使3种麦冬都具有十分独特的蛋白结构,其EPSPS的91位谷氨酸(Glu)缺失,降低了EPSPS与草甘膦的亲和力,因此3种麦冬草对草甘膦天生不敏感[65]。
3.2.2 EPSPS拷贝数和表达量 增加EPSPS拷贝数和表达量也是提高杂草耐受草甘膦的原因之一。毛婵娟等研究发现,3种麦冬杂草对草甘膦耐受性从大到小表现为阔叶麦冬>土麦冬>麦冬,其部分原因是麦冬和土麦冬的EPSPS基因各具有2个拷贝,阔叶麦冬EPSPS有3个拷贝,并且在喷施过草甘膦之后,3种麦冬杂草的EPSPS表达量都显著上调[65-66]。Yuan等通过使用诺瑟杂交(northen blot)对比狗肝菜和熊耳草(Ageratum houstonianum)在喷施草甘膦后测定体内EPSPS mRNA表达量,发现耐受型杂草狗肝菜比敏感型杂草熊耳草的EPSPS mRNA增长相对缓慢、持久和稳定[55]。Hollnder-Czytko等研究发现,在 5 mmol/L 草甘膦存在下,细胞培养耐草甘膦紫堇(C. sempervirens)植物的EPSP合成酶,与草甘膦敏感型植株的比较,EPSP合成酶活性增加了30~40倍,mRNA翻译活性以及EPSP合成酶的转录水平增加了8~12倍。EPSP合成酶特异性DNA序列的相对丰度上两者之间没有明显差异,耐草甘膦紫堇(C. sempervirens)植物的EPSP合成酶的过度产生不是基于相应基因的扩增[33]。
3.2.3 有利的形态生理特性 杂草形态生理特性会影响其对草甘膦的吸收和抵抗能力。Ziska等发现,偃麦草(Elytrigia repens)在高浓度CO2环境条件下,植物气孔关闭,蒸腾作用减少,减少对草甘膦的摄入[70],从而提高了对草甘膦的耐受性。朱金文研究发现,喜旱莲子草在高浓度锰离子胁迫下,其地下根茎会显著增长,适应环境能力增加,提高喜旱莲子草对草甘膦的耐受程度[71]。叶片表皮特殊结构与杂草对草甘膦的耐受性也有联系。毛婵娟等研究发现,3种麦冬叶片上的气孔主要位于下表皮,显著降低了植物对除草剂的吸收。另外通过扫描电镜发现,与麦冬气孔下陷明显、叶片表皮细胞平周壁瘤状细胞外突相比较,阔叶土麦冬气孔下陷不明显,叶片表皮细胞平周壁突起不明显,叶片表面光滑使得药液在叶片上的存留减少,更能减少对草甘膦的吸收,这样就很好地解释了阔叶土麦冬对草甘膦的耐受性最高的原因[65]。叶片角质层也是影响杂草对草甘膦耐受性的一个重要因素。Norsworthy等研究杂草瘤梗番薯因其角质层上有特殊的化学组成显著减少了药剂的摄入,与角质层的含蜡量关系不大[72]。刘小龙报道的与此结论相同,通过对比藜、反枝苋和铁苋菜的蜡含量,发现蜡含量的高低与草甘膦耐受性大小之间并没有直接的联系[66]。Cruz-Hipolito等报道的与此研究结果相反,其认为角质层中蜡含量越高越可以有效地减少药剂渗透进入植物组织中[73]。
3.2.4 有效的代谢机制 杂草植株对药剂的运输传导直接影响了对草甘膦耐受性。Sammons等研究发现,草甘膦在苘麻中是从源到库运输的,源的“推力”是运输所必需的,而库的“拉力”则决定了草甘膦在苘麻不同组织中的分配,从而决定了苘麻不同组织器官对草甘膦敏感性不同,报道认为草甘膦在植物不同组织中的分配与剂量和表面活性剂的种类无关[74]。Maroli等通过比较杂草瘤梗番薯对草甘膦的高耐受型和低耐受型的种群,发现低耐受生物型的种群植物体内所含碳类代谢物质较多,表明其具有更大潜能将草甘膦从“源”运输到“库”中,因草甘膦运输是伴随着糖类运输完成的。而高耐受型种群中则含有种类更加丰富的有机酸、氨基酸和糖类[45]。
4 展望
由于在GR作物田中过度依赖草甘膦除草,使得抗草甘膦雜草种类发展到43种,而耐性杂草更应不容忽视。耐性杂草因其独特的生理结构使得它们在推荐剂量下无法死亡,这不但会降低杂草防除效果、增加杂草防除成本,而且在长期喷施草甘膦农田中会发生杂草群落的演替,耐草甘膦杂草会逐渐成为优势杂草,从而导致草甘膦的过量使用并危害农田生态环境,耐受性杂草会成为草甘膦应用最大的阻碍[75]。杂草对草甘膦的耐性可能多种耐性机制(包括表型、生理、分子层面)并存,因此在耐草甘膦杂草的控制上,一方面要确保杂草管理的多样性,这可以通过寻找新的除草剂类型提供更好的综合防控技术,多种除草方式相结合的管理模式进行轮作等;另一方面,提高施药技术,加强管理,注意施药时期和时间等[76-79]。在施药时可以与其他药剂混用,增加除草杀草谱,提高防效。如在GR大豆田中,草甘膦可以与咪唑乙烟酸、唑酮草酯、氟磺草醚、精喹禾灵、烯草酮等混用[80-81],在GR玉米田中可以与莠去津、硝磺草酮和2,4-D等混用[76,82]。我国在2014年启动了草甘膦和草铵膦的“双草认证”项目[4]。
2017年我国转基因农产品种植面积达到了2.8万hm2,是全球第八大转基因作物种植国家[11],但是还未有转基因抗除草剂作物的种植。目前84.5%的EPSPS基因专利被美国孟山都公司、拜耳集团、美国先锋公司、先正达集团等四家巨头公司掌握。相比之下,我国所获得的EPSPS基因专利数量少、保护范围窄[83]。抗草甘膦基因可以从微生物或植物中获得,而我国目前抗草甘膦基因主要来源于微生物。与微生物相比,植物中具有更加丰富的除草剂抗性基因和除草剂代谢基因[84]。因此开展对草甘膦耐受性杂草的研究可以发掘更多对草甘膦不敏感的基因,为开发抗草甘膦转基因作物提供新的思路。此外,研究杂草对草甘膦的耐受性机制有助于对基础植物生理生化过程和植物对有毒化学物质防卫机制的理解,更有助于对耐受性杂草制定有效防除措施[15]。
参考文献:
[1]苏少泉. 草甘膦述评[J]. 农药,2005(4):145-149.
[2]苏少泉,滕春红. 草甘膦应用现状与未来发展[J]. 世界农药,2014,36(3):8-11.
[3]何秀玲. 草甘膦和抗草甘膦作物的崛起与展望[J]. 世界农药,2018(3):51-54.
[4]杨益军. 2018年中国(全球)草甘膦市场分析[J]. 农药市场信息,2018(5):27-31,36.
[5]United States Environmental Protection Agency. Reregistration eligibility decision(RED):glyphosate[EB/OL]. (2018-12-20)[2012-04-07]. http://www.epa. gov/oppsrrd1//REDs/old _ reds glyphosate. pdf.
[6]李香菊,梁帝允,袁会珠. 除草剂科学使用指南[M]. 北京:中国农业科学技术出版社,2015:5-8.
[7]陈世国,强 胜,毛婵娟. 草甘膦作用机制和抗性研究进展[J]. 植物保护,2017,43(2):17-24.
[8]Schuette J. Environmental fate of glyphosate[J]. Environmental Monitoring & Pest Management,1998,1(1):1-13.
[9]李 波,郭德华,朱 坚,等. 植物产品中草甘膦及其代谢物残留量的GC-MS分析[C]//全国有机质谱学术交流会. 丽江:中国物理学会,中国分析测试学会,2005:345-348.
[10]张 冬,张 宇,王 萌,等. 草甘膦对植物生理影响的研究进展[J]. 热带农业科学,2016,36(9):55-61.
[11]Global status of commercialized biotech/GM crops in 2017:biotech crop adoption surges as economic benefits accumulate in 22 years[M]. ISAAA,2017:1-88.
[12]Brookes G,Barfoot P. GM crops:global socio-economic and environmental impacts 1996—2015[R]. Dorchester,UK:PG Economics Ltd,2017.
[13]黎 裕,王天宇. 玉米转基因技术研发与应用现状及展望[J]. 玉米科学,2018,26(2):1-15,22.
[14]Global status of commercialized biotech/GM crops:2016[M]. ISAAA,2016.
[15]王园园,王 敏,相世刚,等. 全球抗除草剂转基因作物转化事件分析[J]. 农业生物技术学报,2018,26(1):167-175.
[16]强 胜. 杂草学[M]. 2版. 北京:中国农业出版社,2009:216-219.
[17]李香菊,崔海兰. 转基因耐草甘膦作物的环境安全性[J]. 植物保护,2011,37(6):38-43.
[18]Powles S B,Preston C. Evolved resistance to glyphosate in rigid ryegrass (Lolium rigidum) in Australia[J]. Weed Science,1998,46(5):604-607.
[19]Gottrup O,OSullivan P A,Schraa R J,et al. Uptake,translocation,metabolism and selectivity of glyphosate in Canada thistle and leafy spurge[J]. Weed Research,1976,16(3):197-201.
[20]Biesboer D D,Darveaux B,Koukkari W L. Controlling leafy spurge and Canada thistle by competitive species[J]. International Economic Review,1994,50(4):1169-1182.
[21]Wyrill J B,Burnside I C. Absorption,translocation,and metabolism of 2,4-D and glyphosate in common milkweed and hemp dogbane[J]. Weed Science,1976,24(6):557-566.
[22]Schultz M E,Burnside O C. Absorption,translocation,and metabolism of 2,4-D and glyphosate in hemp dogbane (Apocynum cannabinum)[J]. Weed Science,1980,28(1):13-20.
[23]Sherrick S L,Holt H A,Hess F D. Effects of adjuvants and environment during plant development on glyphosate absorption and translocation in field bindweed (Convolvulus arvensis)[J]. Weed Science,1986:811-816.
[24]de Gennaro F P,Weller S C. Differential susceptibility of field bindweed (Convolvulus arvensis) biotypes to glyphosate[J]. Weed Science,1984,32(4):472-476.
[25]Duncan C N,Weller S C. Heritability of glyphosate susceptibility among biotypes of field bindweed[J]. Journal of Heredity,1987,78(4):257-260.
[26]Westwood J H,Weller S C. Cellular mechanisms influence differential glyphosate sensitivity in field bindweed (Convolvulus arvensis) biotypes[J]. Weed Science,1997,45(1):2-11.
[27]劉 延,张朝贤,黄红娟,等. 草甘膦对田旋花和打碗花体内莽草酸含量的影响[J]. 杂草科学,2008(2):10-12.
[28]刘 延. 田旋花和打碗花对草甘膦的耐药性研究[D]. 北京:中国农业科学院,2008:55-84.
[29]张 猛,刘 延,张朝贤,等. 田旋花对草甘膦的耐药性机制[J]. 植物保护学报,2011,38(6):551-556.
[31]張 猛. 田旋花(Convolvulus arvensis L.)对草甘膦耐药性机理研究[D]. 中国农业科学院,2011:21-29.
[32]黄兆峰. 田旋花对草甘膦耐药性分子机制[D]. 北京:中国农业科学院,2014:18-35.
[33]Hollnder-Czytko H,Johnning D,Meyer H E,et al. Molecular basis for the overproduction of 5-enolpyruvylshikimate 3-phosphate synthase in a glyphosate-tolerant cell suspension culture of Corydalis sempervirens[J]. Plant Molecular Biology,1988,11(2):215-220.
[34]Boerboom C M. Selection and characterization of glyphosate tolerance in birdsfoot trefoil (Lotus corniculatus)[R]. Minnesota Univ,1989.
[35]Boerboom C M,Wyse D L,Somers D A. Mechanism of glyphosate tolerance in birdsfoot trefoil (Lotus corniculatus)[J]. Weed Science,1990,38(6):463-467.
[36]Boerboom C M,Ehlke N J,Wyse D L,et al. Recurrent selection for glyphosate tolerance in birdsfoot trefoil[J]. Crop Science,1991,31(5):1124-1129.
[37]Hartzler R G. Velvetleaf (Abutilon theophrasti) interference in soybean (Glycine max):a survey of yield loss estimates and management recommendations[J]. Crop Protection,1997,16(5):480-485.
[38]Hartzler R G,Battles B A. Reduced fitness of velvetleaf (Abutilon theophrasti) surviving glyphosate[J]. Weed Technology,2001,15(3):492-496.
[39]Fuchs M A,Geiger D R,Reynolds T L,et al. Mechanisms of glyphosate toxicity in velvetleaf (Abutilon theophrastimedikus)[J]. Pesticide Biochemistry & Physiology,2002,74(1):27-39.
[40]Zhou J,Bo T,Messersmith C G,et al. Glyphosate efficacy on velvetleaf (Abutilon theophrasti) is affected by stress[J]. Weed Science,2007,55(3):240-244.
[41]Norsworthy J K,Burgos N R,Oliver L R. Differences in weed tolerance to glyphosate involve different mechanisms[J]. Weed Technology,2001,15(4):725-731.
[42]Shaw D R,Arnold J C. Weed control from herbicide combinations with glyphosate[J]. Weed Technology,2002,16(1):1-6.
[43]Koger C H,Reddy K N. Glyphosate efficacy,absorption,and translocation in pitted morningglory (Ipomoea lacunosa)[J]. Weed Science,2005,53(3):277-283.
[44]Ribeiro D,NandulaV,Dayan F E,et al. Possible glyphosate tolerance mechanism in pitted morningglory (Ipomoea lacunosa L.)[J]. J Agric Food Chem,2015,63(6):1689-97.
[45]Maroli A S,Nandula V K,Duke S O,et al. Comparative metabolomic analyses of Ipomoea lacunosa biotypes with contrasting glyphosate tolerance captures herbicide-induced differential perturbations in cellular physiology[J]. J Agric Food Chem,2018,66(8):2027-2039.
[46]Santos I C,Silva A A,Ferreira F A,et al. Efficiency of glyphosate in the control of Commelina benghalensis and Commelina diffusa[J]. Planta Daninha,2001,19(1):135-143.
[47]Wilson R G,Yonts C D,Smith J A. Influence of glyphosate and glufosinate on weed control and sugarbeet (Beta vulgaris) yield in herbicide-tolerant sugar beet[J]. Weed Technology,2002,16(1):66-73.
[48]Loux M M,Stachler J M,Miller B A,et al. Response of common lambsquarters to glyphosate in the greenhouse and growth chamber[C]//Proc N Cent Weed Sci Soc,2005:202.
[49]Boerboom C M,Stoltenberg D E,Jeschke M R,et al. Factors affecting glyphosate control of common lambsquarters[C]//Proc N Cent Weed Sci Soc,2006:54.
[50]Schuster C L,Al-Khatib S K. Response of common lambsquarters (Chenopodium album) to glyphosate as affected by growth stage[J]. Weed Science,2007,55(2):147-151.
[51]Kniss A R,Miller S D,Westra P H,et al. Glyphosate susceptibility in common lambsquarters (Chenopodium album) is influenced by parental exposure[J]. Weed Science,2007,55(6):572-577.
[52]Westhoven A M,Stachler J M,Loux M M,et al. Management of glyphosate-tolerant common lambsquarters (Chenopodium album) in glyphosate-resistant soybean[J]. Weed Technology,2008,22(4):628-634.
[53]Sivesind E C,Gaska J M,Jeschke M R,et al. Common lambsquarters response to glyphosate across environments[J]. Weed Technology,2011,25(1):44-50.
[54]Yerka M K,Wiersma A T,Lindenmayer R B,et al. Reduced translocation is associated with tolerance of common lambsquarters (Chenopodium album) to glyphosate[J]. Weed Science,2013,61(3):353-360.
[55]Yuan C I,Chaing M Y,Chen Y M. Triple mechanisms of glyphosate-resistance in a naturally occurring glyphosate-resistant plant Diclipterachinensis[J]. Plant Science (Shannon),2002,163(3):0-554.
[56]Monquero P A,Christoffoleti P J. Seed bank dynamics in areas with frequent glyphosate application[J]. Planta Daninha,2003,21(1):63-69.
[57]Culpepper A S,Flanders J T,York A C,et al. Tropical spiderwort (Commelina benghalensis) control in glyphosate-resistant cotton[J]. Weed Technology,2004,18(2):432-436.
[58]Santos S A D,Tuffisantos L D,SantAnnasantos B F,et al. Influence of shading on the leaf morphoanatomy and tolerance to glyphosate in Commelina benghalensis L. and Cyperusro tundus L.[J]. Australian Journal of Crop Science,2015,9(2):135-142.
[59]Park N I,Lee I Y,Kwon O S,et al. Tolerant mode of dayflower (Commelin acommunis L.) to glyphosate[J]. Korean Journal of Weed Science,2004,24(3):230-236.
[60]Ulloa S M,Owen M D K. Response of asiatic dayflower (Commelina communis) to glyphosate and alternatives in soybean[J]. Weed Science,2009,57(1):74-80.
[61]Vargas J M G. Glyphosate-tolerant asiatic dayflower (Commelina communis L.):ecological,biological and physiological factors contributing to its adaptation to Iowa agronomic systems[D]. Ames:Iowa State University,2012:66-99.
[62]Owen M D. Weed species shifts in glyphosate‐resistant crops[J]. Pest Management Science,2008,64(4):377-387(11).
[63]Vidal R A,Rainero H P,Kalsing A,et al. Exploration of herbicide associations to prevent glyphosate tolerant and resistant weeds[J]. Planta Daninha,28(1),159-165.
[64]Mao C,Xie H,Chen S,et al. Multiple mechanism confers natural tolerance of three lilyturf species to glyphosate[J]. Planta,2015,243(2):321-335.
[65]毛嬋娟,解洪杰,宋小玲,等. 麦冬草对草甘膦耐药性的形态学机制[J]. 杂草学报,2016,34(1):1-7.
[66]刘小龙. 铁苋菜(Acalypha australis L.)对草甘膦的耐受性机理研究[D]. 北京:中国农业科学院,2016:29-35.
[67]吴加军. 不同剂型草甘膦药效评价与抗(耐)草甘膦杂草监测[D]. 南京:南京农业大学,2006:45-56.
[68]费云燕,盖钧镒,赵团结. 南京大豆田间耐草甘膦杂草的种类与特性鉴定[J]. 江苏农业科学,2016,44(11):154-156.
[69]Owen M D,Zelaya I A. Herbicide-resistant crops and weed resistance to herbicides[J]. Pest Management Science,2005,61(3):301-11.
[70]Ziska L H,Teasdale J R. Sustained growth and increased tolerance to glyphosate observed in a C3 perennial weed,quackgrass (Elytrigia repens),grown at elevated carbon dioxide[J]. Functional Plant Biology,2000,27(2):159-166.
[71]朱金文. 高锰胁迫下空心莲子草的生理生化特性和草甘膦耐性研究[D]. 杭州:浙江大学,2008:37-57
[72]Norsworthy J K,Oliver L R. Effect of irrigation,soybean (Glycine max) density,and glyphosate on hemp sesbania (Sesbania exaltata) and pitted morningglory (Ipomoea lacunosa) interference in soybean[J]. Weed Technology,2002,16(1):7-17.
[73]Cruz-Hipolito H,Rojano-Delgado A,José A Domínguez-Valenzuela,et al. Glyphosate tolerance by Clitoria ternatea and Neonotonia wightii plants involves differential absorption and translocation of the herbicide[J]. Plant and Soil,2011,347(1-2):221-230.
[74]Sammons R D,Feng P C C,Chiu T. Glyphosate efficacy is contributed by its tissue concentration and sensitivity in velvetleaf (Abutilon theophrasti)[J]. Pesticide Biochemistry & Physiology,2003,77(3):83-91.
[75]筱 禾. 草甘膦的历史与现状[J]. 世界农药,2018,40(3):46-50.
[76]Eure P M,Stanley C A,Merchant R M,et al. Weed control,cropresponse,and profitability when intercropping cantaloupe and cotton[J]. Weed Technology,2016,29(2):217-225.
[77]Eure P M,Jordan D L,Fisher L R,et al. Efficacy of herbicides when spray solution application is delayed[J]. International Journal of Agronomy,2013.
[78]Morris S A,Pratt D. Analysis of the Lotka-Volterra competition equations as a technological substitution model[J]. Technological Forecasting & Social Change,2003,70(2):103-133.
[79]David P,Kells J J,Donald P. Substitutes for ammonium sulfate as additives with glyphosate and glufosinate[J]. Weed Technology,2003,17(3):576-581.
[80]Kimberly D W,Nader S,Lynette R B,et al. Weed control with postemergence glyphosate tank mixes in glyphosate-resistant soybean[J]. Canadian Journal of Plant Science,2014,94(94):1239-1244.
[81]Vidal N R,Vidal R A. Augmented reality systems for weed economic thresholds applications[J]. Planta Daninha,2010,28(2):449-454.
[82]Scroggs D M,Miller D K,Griffin J L,et al. Glyphosate efficacy on selected weed species is unaffected by chemical coapplication.[J]. Weed Technology,2005,19(4):1012-1016.
[83]Sikkema P H,Christy S,Hamill A S,et al. Response of common lambsquarters (Chenopodium album) to glyphosate application timing and rate in glyphosate-resistant corn[J]. Weed Technology,2004,18(4):908-916.
[84]宋 敏,劉丽军,苏颖异. 抗草甘膦EPSPS基因的专利保护分析[J]. 中国生物工程杂志,2010,30(2):147-152.
