不同林草复合配置对长柄扁桃林地土壤养分和酶活性的影响
2019-10-11王进鑫冯树林吕国利刘潇阳王怀彪王玉宝
黄 优, 王进鑫, 冯树林, 吕国利, 刘潇阳, 狄 龙, 王怀彪, 王玉宝
(1.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100; 2.西北农林科技大学 资源环境学院,陕西 杨陵 712100; 3.榆林市林业产业开发办公室, 陕西 榆林719000; 4.榆林市榆阳区林业工作站, 陕西 榆林 719000)
榆林沙区位于毛乌素沙地南缘,是我国北方的自然生态屏障。榆林市榆阳区沙地矿产资源丰富,近年来大规模开发利用使当地经济迅速发展,但开发的同时使许多采矿区出现地面塌陷、土壤养分下降等问题[1],生态破坏日益严重,经济发展与环境资源之间的矛盾日趋突出,生态恢复迫在眉睫[2;3]。近年来,有关学者对矿区治理优良树种选择及其空间配置的研究表明榆林矿区乔木层由单一树种杨树变成以樟子松为优势种的群落,灌木层长柄扁桃、沙地柏的优势得到明显体现,改善了植被结构和土壤养分状况。
目前长柄扁桃的研究主要集中在其生物学特性[4]、引种栽培[5]、生态经济价值[6]等方面,关于长柄扁桃与其他植物复合配置的种间关系还未见相关报道。长柄扁桃(Amygdaluspedunculata)作为毛乌素沙区优良固沙植物且适宜沙地栽培的木本油料植物[7],其自身价值决定了其在发展生物质能源和保护榆林沙区脆弱生态环境方面上的应用前景[8]。目前榆林地区已发展长柄扁桃林约6.67×104hm2,然而由于沙地土壤养分贫瘠,造成人工林生长十分缓慢。已有研究表明不同植被种植模式对土壤的质量和土壤理化性质的影响不同[9-10],不同林草间作模式在一定程度上有利于提高土壤养分含量和营养水平[11-12],通过合理配置植被恢复模式对土壤养分积累和酶活性的促进都有明显改善作用[13-14]。为了充分利用物种间的互利共生作用,改善长柄扁桃林地养分状况,我们早期开展了长柄扁桃与当地主要适生草种种间关系的研究,通过室内研究发现,苜蓿、沙打旺等豆科牧草可促进长柄扁桃种子萌发和幼苗生长,适合与长柄扁桃进行搭配形成林草复合系统[15],但未进一步对其田间试验效果进行研究。因此,研究矿区不同林草复合配置对长柄扁桃林地土壤养分与酶活性的影响具有重要意义。本研究选择榆林市乡土树种长柄扁桃为目的树种,于2013年进行长柄扁桃不同林草复合配置模式的设计与种植,在2018年进行土壤养分与土壤酶活性的调查分析,对比不同复合配置模式对长柄扁桃土壤养分和酶活性的影响,筛选出对土壤改良效果较好的种植模式,为榆林煤矿塌陷区的植被恢复与生态建设提供理论依据。
1 研究区概况
研究区位于榆林市煤炭矿区,该地区矿藏资源丰富,煤炭储量2.71×1011t,占全国的1/3[16],采空区达2 925.64 km2,塌陷面积361.93 km2;经度109°6′E,纬度38°20′N,平均海拔1 154 m,属毛乌素沙地南缘风沙草滩区,地势平缓,大多为波状起伏的固定、半固定沙地,土壤为风沙土,其表层疏松,沙地降雨可快速渗入地下形成地下水,水质好可作为优质灌溉水,有利于发展沙区经济林产业。该地属于温带半干旱大陆性边缘季风气候,气候干燥,日照充足。年平均气温8.3 ℃,年均降水量365.7 mm,主要集中在7—9月,年均蒸发量为1 508~2 502 mm,无霜期156 d,干湿交替明显。该区主要优势植被有樟子松(Pinussylvestris)、沙柳(Salixcheilophila)、沙蒿(Artemisiadesertorum)、柠条(Caraganakorshinskii)、沙竹(Psammochloavillosa)、紫穗槐(Amorphafruticosa)等。
2 材料与方法
2.1 样地设置
在榆林补浪河乡魏家峁村长柄扁桃基地布设6种长柄扁桃与草本的配置模式:沙打旺(Astragalusadsurgens)+长柄扁桃(SC)、草木樨(Melilotussuaveolens)+长柄扁桃(CC)、苜蓿(Medicagosativa)+长柄扁桃(MC)、红豆草(Onobrychisviciifolia)+长柄扁桃(HC)、小冠花(Coronillavaria)+长柄扁桃(XC)、长柄扁桃纯林(CK)作空白对照。其中长柄扁桃林龄为5 a,平均株高149.96 cm,平均冠幅154.77 cm×153.97 cm,株行距2.0 m×3.0 m,草种沙打旺、草木樨、苜蓿、红豆草、小冠花采用条播法,在长柄扁桃(2.0 m×3.0 m)行间进行播种,即果园行间生草,生草带边缘远离树干约1.0 m。春季条播,行距20~25 cm,播深2~3 cm,播后镇压,草带宽1.0 m,采样时草种覆盖度达65%~97%,高度为30~123 cm。
2.2 样品采集和处理
于2018年9月份进行土壤取样,土样采集方法按照多点混合采样法,每个样品由S形6点样混合而成,在长柄扁桃行间草带处取样,深度为0—20 cm,20—40 cm和40—60 cm,同时设置纯林为空白对照组,每种配置模式的土壤样品按四分法留取500 g后,去除植物根系、碎石、动植物残体等杂物装入塑封袋带回实验室,过筛后储存备用。
2.3 指标测定方法
2.3.1 土壤养分指标测定 有机质(OM)采用重铬酸钾容量法;全氮(TN)采用凯氏定氮法;碱解氮(AN)采用碱解扩散法;速效钾(AK)采用醋酸铵浸提火焰光度法;速效磷(AP)采用碳酸氢钠浸提钼锑抗比色法。每个处理测定3个重复,每个重复测定3次,结果取其平均值[17]。
2.3.2 土壤酶活性测定 蔗糖酶活性采用3,5-二硝基水杨酸比色法测定,过氧化氢酶活性采用高锰酸钾滴定法测定,脲酶活性采用苯酚—次氯酸钠比色法测定,碱性磷酸酶活性采用磷酸苯二纳比色法测定。每个处理测定3个平行样本,每个样本测定3次,结果取其平均值[18]。
2.4 统计分析
采用Excel和SPSS 20.0软件对试验数据进行处理和统计分析,Origin 9.0进行作图。用one-way ANOVE和LSD法进行方差分析和多重比较,不同字母表示显著性差异,用Pearson进行不同指标相关性分析。
3 结果与分析
3.1 不同复合配置模式的土壤养分特征
土壤养分可以反映土壤性能的优劣,它既影响植物生长,又被植物生长反作用从而影响土壤质量状况[19]。林草复合种植后,土壤有机质、全氮、碱解氮、速效磷、速效钾均有明显增加。土壤有机质主要来源于动植物残体、根系作用和生物作用等。从表1可以看出,不同土壤层有机质随着土层深度的增加,有机质含量明显增加,均呈现一定的表聚性特征。与对照相比,SC,CC,MC,HC,XC处理的0—20 cm土层土壤有机质含量分别增加了230.23%,208.82%,358.20%,104.82%和166.67%,均与对照达到显著水平(p<0.05);在20—40 cm土层,各配置模式与对照相比,有机质含量分别增加了157.06%,210.22%,310.22%,129.44%和85.40%;40—60 cm土层,增幅分别为136.41%,349.49%,406.67%,102.31%和123.33%;与表层相似,20—40 cm和40—60 cm土层土壤有机质含量各配置模式均较对照显著增加(p<0.05)。
表1 不同林草复合配置长柄扁桃土壤有机质含量g/kg
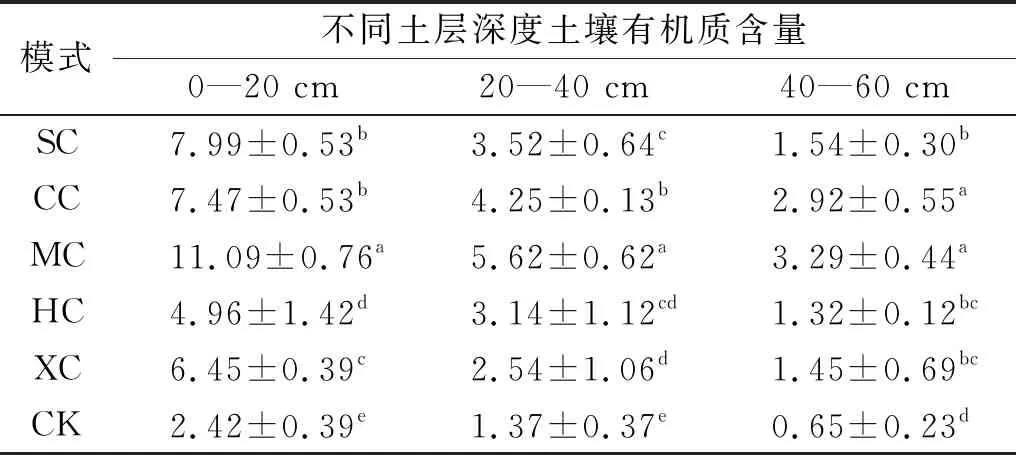
模式不同土层深度土壤有机质含量0—20 cm20—40 cm40—60 cmSC7.99±0.53b3.52±0.64c1.54±0.30bCC7.47±0.53b4.25±0.13b2.92±0.55aMC11.09±0.76a5.62±0.62a3.29±0.44aHC4.96±1.42d3.14±1.12cd1.32±0.12bcXC6.45±0.39c2.54±1.06d1.45±0.69bcCK2.42±0.39e1.37±0.37e0.65±0.23d
注:SC:沙打旺+长柄扁桃; CC:草木樨+长柄扁桃; MC:苜蓿+长柄扁桃; HC:红豆草+长柄扁桃; XC:小冠花+长柄扁桃; CK:长柄扁桃对照。不同字母表示同一土层中不同植被恢复模式间差异显著(p<0.05)。下同。
全氮含量是衡量土壤氮素储备供应能力的指标,氮含量越高表明土壤供氮能力越强[20]。从图1可以看出,复合配置模式土壤全氮含量平均波动范围在0.13~0.90 g/kg,属于中等水平。除表层是CC全氮含量最高外,20—40 cm和40—60 cm土层土壤全氮含量均表现为:SC>CC>MC>HC>XC>CK,且SC,CC,MC均较对照显著增加(p<0.05),20—40 cm土层分别增加了78.89%,68.89%和60.00%,40—60 cm土层增加幅度分别为75.76%,57.58%和48.48%。碱解氮含量高低反映了土壤中近期氮素供给水平,以及植物生长可利用氮素的高低。5种复合配置模式和长柄扁桃单独种植土壤碱解氮含量都表现出明显的层次性,不同土层碱解氮含量变化趋势也一致,均表现为:MC>SC>CC>HC>XC>CK。增幅分别为MC 40.43%~127.54%,CC 26.91%~77.45%,SC 38.83%~114.84%,HC 12.08%~85.16%,XC 4.73%~64.91%,除XC外,其他草种与长柄扁桃复合配置均达到显著水平(p<0.05)。土壤速效磷反映土壤供磷能力。从不同土层土壤速效磷含量对比来看,0—20 cm土层土壤速效磷含量明显高于20—40 cm和40—60 cm,层次差异明显。不同复合配置模式对土壤速效磷含量的增加十分有限,与赵燕娜等[21]对榆林沙区灌木固沙林土壤速效磷性质测定结果一致。与对照相比,只有MC增加1.29%(0—20 cm),22.32%(40—60 cm),但增加效果不显著(p>0.05),20—40 cm SC,CC,MC均显著增加,增幅分别为25.33%,23.33%,45.86%。其他草本与长柄扁桃复合配置土壤速效磷含量较对照均有所降低,说明草本植物的生长对于土壤速效磷含量的增加十分有限。速效钾能够被植物直接吸收利用的钾素,可以判断土壤中钾素的实际供应状态。6种配置模式不同土层速效钾含量对比表明,随着土壤深度增加,速效钾含量随之降低,表层土壤速效钾含量最高,但20—40 cm和40—60 cm土层速效钾含量下降不明显。在0—20 cm土层土壤速效钾含量较对照分别增加了47.24%,25.84%,34.10%,11.55%,1.11%;在20—40 cm土层,各配置模式与对照相比,速效钾含量分别增加了74.92%,29.40%,54.41%,24.05%,22.25%;40—60 cm土层,增幅分别为75.74%,44.18%,78.99%,45.24%,31.55%;各种配置模式土壤速效钾均较对照显著增加,说明植物对土壤速效钾的含量具有一定的改善作用。
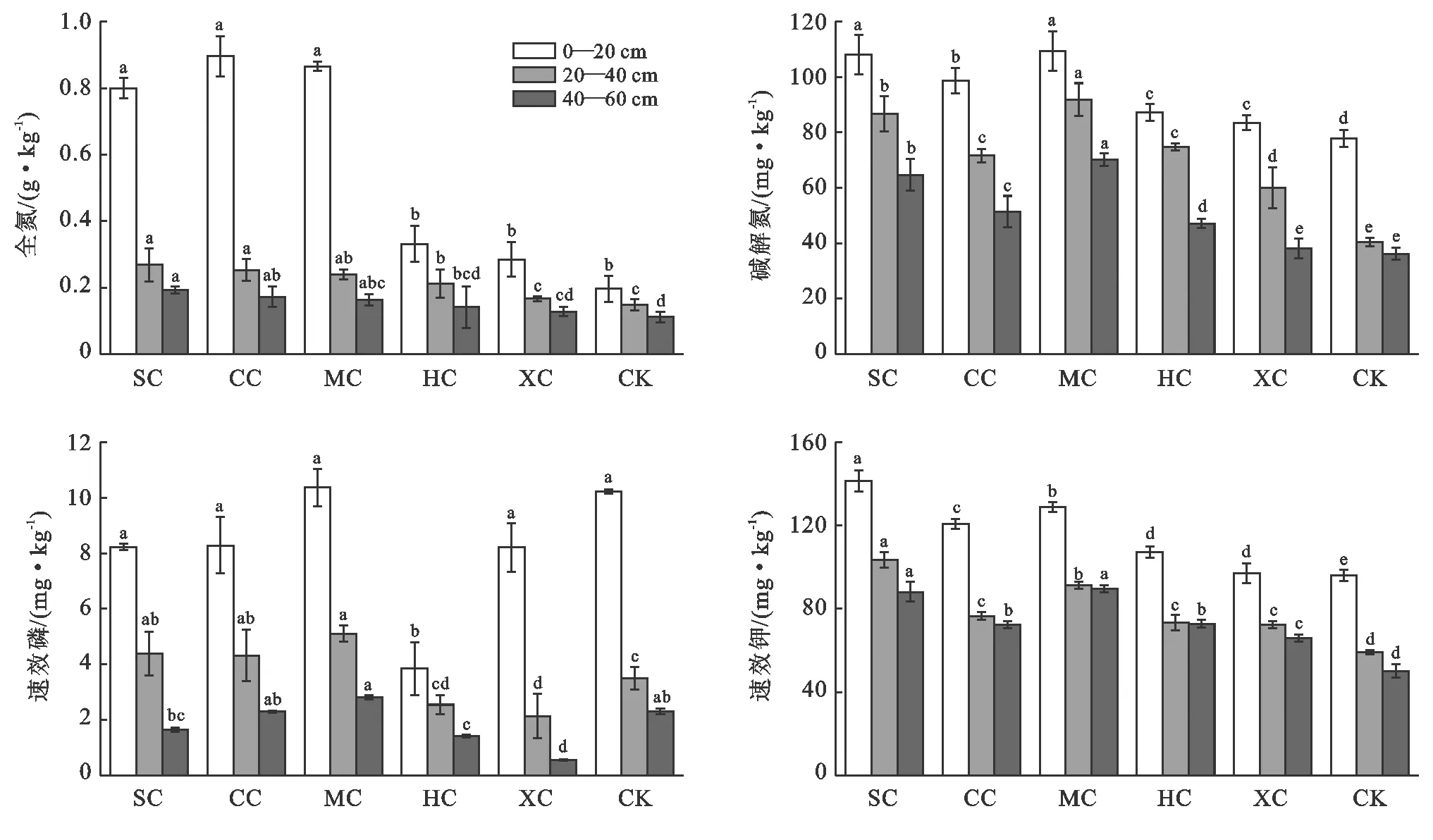
图1 不同林草复合配置模式长柄扁桃土壤氮磷钾含量变化
注:SC为沙打旺+长柄扁桃,CC为草木樨+长柄扁桃,MC为苜蓿+长柄扁桃,HC为红豆草+长柄扁桃,XC为小冠花+长柄扁桃,CK为长柄扁桃对照;同组不同小写字母为p<0.05水平下差异显著。下同。
3.2 不同复合配置模式土壤酶活性特征
不同复合配置模式长柄扁桃土壤酶活性方差分析结果如图2所示。除XC外,其余复合配置模式土壤蔗糖酶活性均有所增加,大小顺序依次为:MC>SC>CC>HC>对照>XC,分别较对照增加了111.21%,75.76%,34.82%,5.30%和-6.11%,MC和SC达到显著水平(p<0.05);在0—20 cm土层,SC、MC模式土壤蔗糖酶活性最高,XC模式活性最低,其他模式间土壤蔗糖酶活性无显著差异(p>0.05);与对照相比,20—40 cm土层中不同复合配置模式长柄扁桃土壤蔗糖酶活性增幅为83.33%~485.09%,均达到显著水平(p<0.05),40—60 cm土层增幅较小,为10.49%~240.78%,MC,SC和CC达到显著水平(p<0.05)。在不同复合配置模式中,0—20 cm和40—60 cm土层的土壤过氧化氢酶与对照相比没有显著差异(p>0.05),20—40 cm土层虽然差异显著但均低于对照,这说明本研究中林草配置模式对过氧化氢酶活性影响不大。与过氧化氢酶类似,不同复合配置模式长柄扁桃土壤脲酶活性均无显著差异(p>0.05),且普遍低于对照,林草复合未增加长柄扁桃土壤过氧化氢酶和脲酶活性。
除HC模式外,其余复合配置模式土壤碱性磷酸酶活性均有所增加,大小顺序为MC>SC>CC>XC>CK>HC,分别较对照增加了44.06%,40.62%,17.71%,3.13%和-9.18%,MC和SC达到显著水平;不同复合配置模式碱性磷酸酶活性的方差分析表明,0—20 cm土层,除HC模式外,与对照均无显著差异(p>0.05),40—60 cm土层碱性磷酸酶较对照增加19.17%~204.98%,且不同复合配置模式均达到显著水平(p<0.05)。
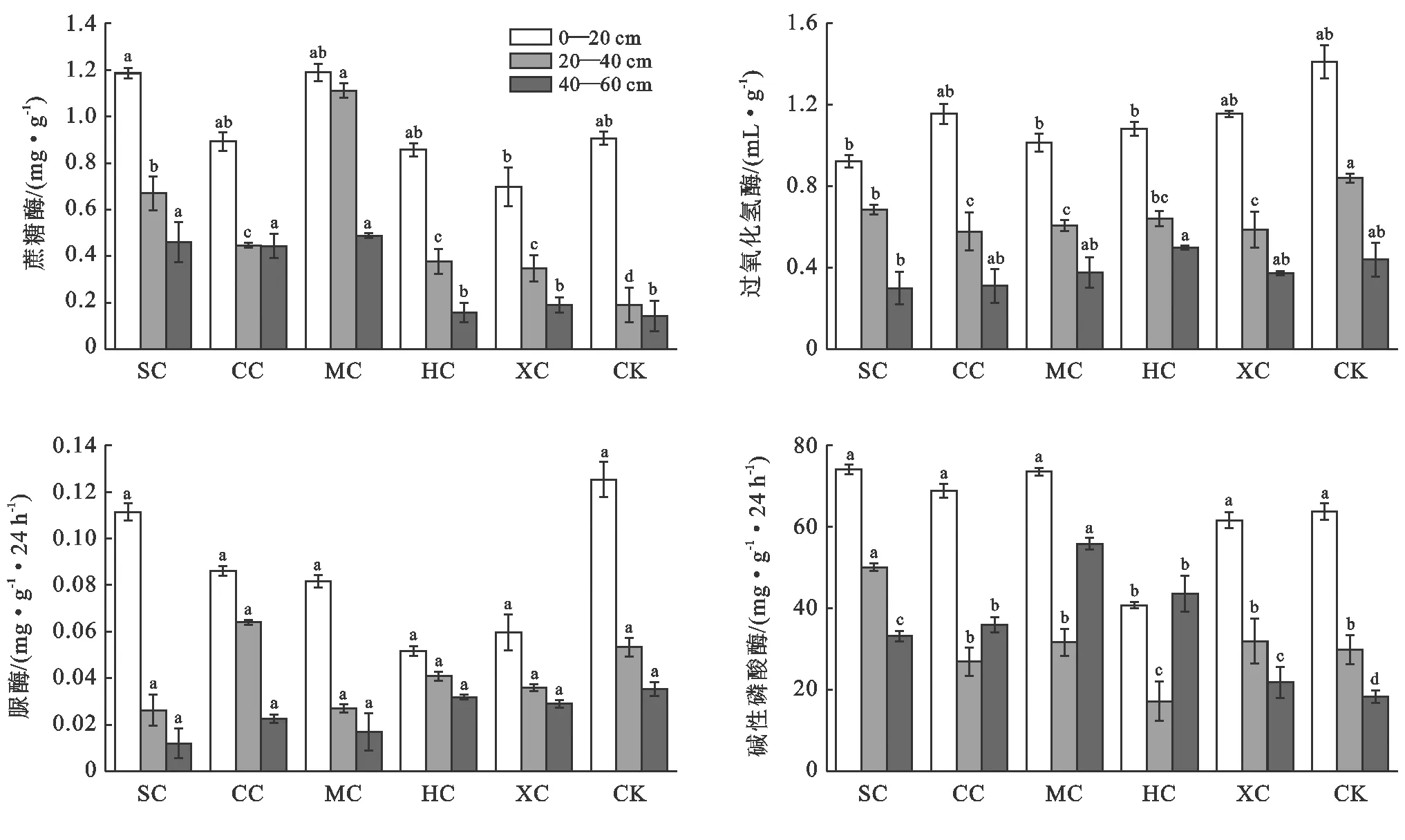
图2 不同林草复合配置模式长柄扁桃土壤酶活性特征
3.3 土壤养分与土壤酶活性的相关性分析
土壤养分含量的高低制约着土壤酶活性的高低,土壤酶活性的反馈作用又会使土壤养分含量发生变化,二者之间相互作用,密不可分[22-23]。为了揭示不同复合配置模式长柄扁桃土壤的变化,对土壤酶活性和养分进行了相关性分析,结果详见表2,蔗糖酶除过在0—20 cm,20—40 cm土层与速效钾含量不显著相关(p>0.05)外,与其余养分含量在不同土层间均显著正相关(p<0.05);过氧化氢酶酶只有在20—40 cm土层与速效磷含量呈显著正相关(p<0.05);脲酶在40—60 cm土层与土壤有机质、碱解氮、速效钾含量呈及显著负相关(p<0.01);碱性磷酸酶与土壤养分普遍具有显著正相关(p<0.05)。整个土层中蔗糖酶除和速效钾外与其它养分指标均呈极显著正相关(p<0.01),过氧化氢酶、碱性磷酸酶与土壤养分指标均达到极显著正相关(p<0.01),而脲酶只与速效磷有显著正相关(p<0.05)。由此说明土壤中各种酶和养分含量之间相互作用共同影响长柄扁桃林地土壤状况。可见,在长柄扁桃林下中种植不同草本,将增强土壤酶活性,从而促进土壤养分的积累。
4 讨论与结论
4.1 讨 论
各种养分含量在剖面上都具有明显的层次差异性,表层0—20 cm土层养分含量最高,向下逐渐降低,不同土层间差异显著。表明土壤养分不仅受植物根系的影响,而且更易受枯枝落叶、气候等其他外界环境影响,同时又反映了植被对土壤养分的表聚效应。在0—60 cm土层中,不同草本与长柄扁桃复合种植有机质含量比对照均显著增加,表现为:MC>CC>SC>XC>HC>CK,分别比对照增加了4.50,3.30,2.94,2.35,2.12倍;全氮、碱解氮、速效磷、速效钾与有机质变化趋势相似,MC,CC,SC增加较为明显,XC和HC增加较少;具体而言,全氮增加幅度为1.29~2.79倍,碱解氮是对照的1.18~1.79倍,速效磷达到1.33~2.23倍,速效钾为1.15~1.62倍。可以看出,不同草本与长柄扁桃复合配置模式对土壤养分状况改善作用不同,但均高于对照。土壤养分含量高低受多种因素影响,但可归纳为两方面:一方面是输入,即进入土壤的养分,主要包括植物凋落物、地下根系分泌和微生物腐解作用等使土壤酶活性和有机质含量增加,提高土壤肥力;另一方面是输出,即通过淋溶作用等将部分土壤养分带走。植物腐殖质分泌的胡敏酸类物质能促进植物生长发育,同时通过络合作用有助于修复污染土壤[17]。单纯种植一种植物容易造成沙地裸露,有机质含量偏低。对照土壤养分含量较低,可能是由于只有长柄扁桃单一植物,没有草本对下层土壤进行覆盖遮蔽,导致降雨对土壤养分的淋溶更高,更不利于土壤养分积累。林下植物能够促进营养元素在土壤表层富集,且草本植物能够通过减少地表径流量来降低磷、钾元素的流失。各种配置模式中全氮、碱解氮含量的大小顺序不同,说明土壤含氮量高的土壤中真正能被植物吸收利用的氮元素不一定高,但几种配置模式氮素含量均高于对照,可能是由于种植豆科牧草的固氮作用,同时促进土壤有机质的累积[24]。
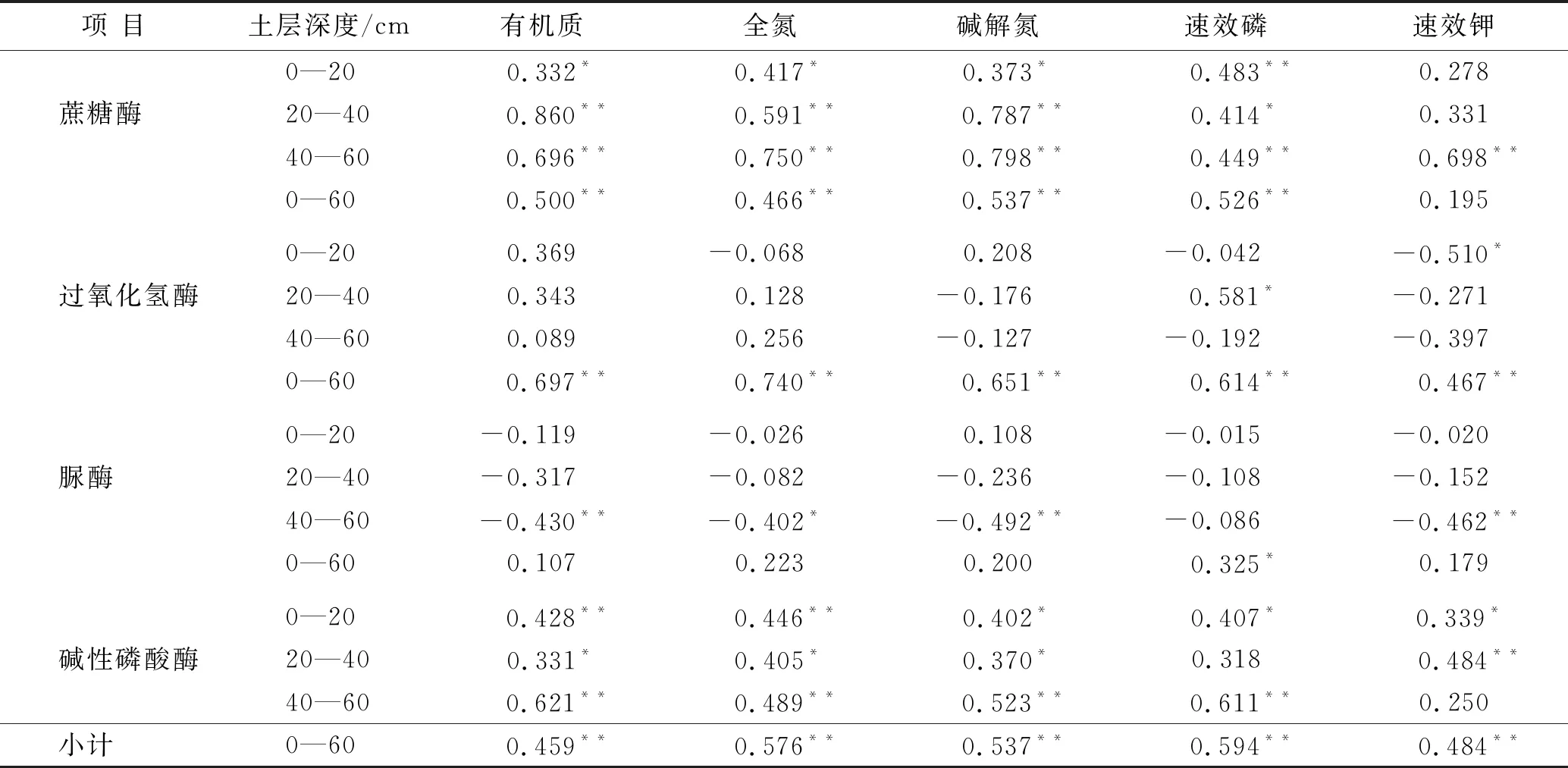
表2 土壤养分含量与土壤酶活性相关性分析
注:**在0.01水平(双侧)上显著相关;*在0.05水平(双侧)上显著相关。
土壤养分和酶活性是林草复合系统的重要组成部分,直接影响林木生长,参与或促进土壤中一系列复杂的生理生化反应,可以客观地反映土壤肥力状况。土壤蔗糖酶、脲酶、碱性磷酸酶和过氧化氢酶能很好地表征土壤碳、氮、磷的供应程度与土壤腐殖化程度及有机质的积累程度[25]。本研究中不同土层土壤酶活性差异显著,均表现为0—20 cm最高。有研究表明红壤丘陵区0—40 cm 土层的脲酶、蔗糖酶和碱性磷酸酶活性随土层深度的增加而逐渐下降[26]。土壤酶活性不仅与土壤理化性质有关,而且与植被类型、土壤微生物数量、种类、酶类本身的性质也密切相关,不同植被类型将产生不同的地被覆盖物,其种类和数量对土壤酶活性的影响也不同[27]。本研究不同复合配置模式在不同土层的土壤酶活性对比发现,MC和SC模式蔗糖酶和碱性磷酸酶活性要高于其他模式,反映了其良好的氮素和磷素供应状况以及较强的分解土壤中营养物质的能力,这与王风芹[28]等和翟辉[25]等研究结果较一致,原因可能是苜蓿、沙打旺等豆科植物其根部共生的根瘤菌可以更好地固定大气中的氮素,进而改善该配置模式长柄扁桃土壤系统的物质组成及肥力水平。复合配置模式物种多样性高于对照,增加了土壤微生物种类和数量,从而促进土壤酶活性的增加,故复合配置模式蔗糖酶和碱性磷酸酶活性高于对照。但过氧化氢酶是氧化还原酶,其活性变化不同于蔗糖酶、脲酶和碱性磷酸酶,在不同复合配置模式中不同于其他酶活性的稳定性,可能是由于参与土壤物质和能量的转化过程,分解土壤中对植物有害的过氧化氢,而不同植被生长,促使土壤有机碳的积累增加,形成的还原条件土壤环境抑制了过氧化氢酶活性的增强。说明不同植物种类的选择对土壤过氧化氢酶的活性影响不同,而对于过氧化氢酶活性的异常变化需要进一步研究。
土壤养分与酶活性之间存在一定的相关性。有研究表明[29]不同植被恢复方式对土壤酶改善作用不同,且土壤全氮与土壤酶活性呈显著或极显著相关,土壤脲酶和碱性磷酸酶活性与土壤养分之间呈显著或极显著相关关系,认为脲酶和碱性磷酸酶可以作为衡量土壤肥力水平的指标。整个土层中,脲酶只与速效磷含量呈正相关(p<0.05),蔗糖酶与有机质、全氮、碱解氮和速效磷均呈显著正相关(p<0.05),过氧化氢酶和碱性磷酸酶与土壤养分指标呈极显著正相关(p<0.01)。本研究结果与桑巴叶等[30]的研究结果一致,但与其他文献结果不同,研究发现土壤中蔗糖酶、过氧化氢酶、碱性磷酸酶活性与土壤主要养分因子间均存在极显著或显著的正相关关系,这种结果差异的原因,还需在今后研究中进一步考证。
4.2 结 论
(1) 草本植物与长柄扁桃的复合种植模式间土壤养分含量与对照相比差异显著。在0—60 cm土层中,不同草本与长柄扁桃复合种植土壤养分含量比对照均显著增加,具体顺序为:MC>CC>SC>XC>HC>CK,其中MC,CC和SC增加较为明显,XC和HC增加较少。
(2) 不同林草复合配置间土壤酶活性差异显著,SC,CC和MC这3种模式明显优于其他模式,HC和XC模式不利于土壤酶活性提高。
(3) 土壤酶活性与养分间关系紧密,不同草种的土壤改良作用各异。该地区选用SC,CC和MC这3种林草配置模式对长柄扁桃林地土壤养分积累和酶活性的促进具有明显改善作用。
